
Invasive Plant Species Biomass—Evaluation of Functional Value

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results and Discussion
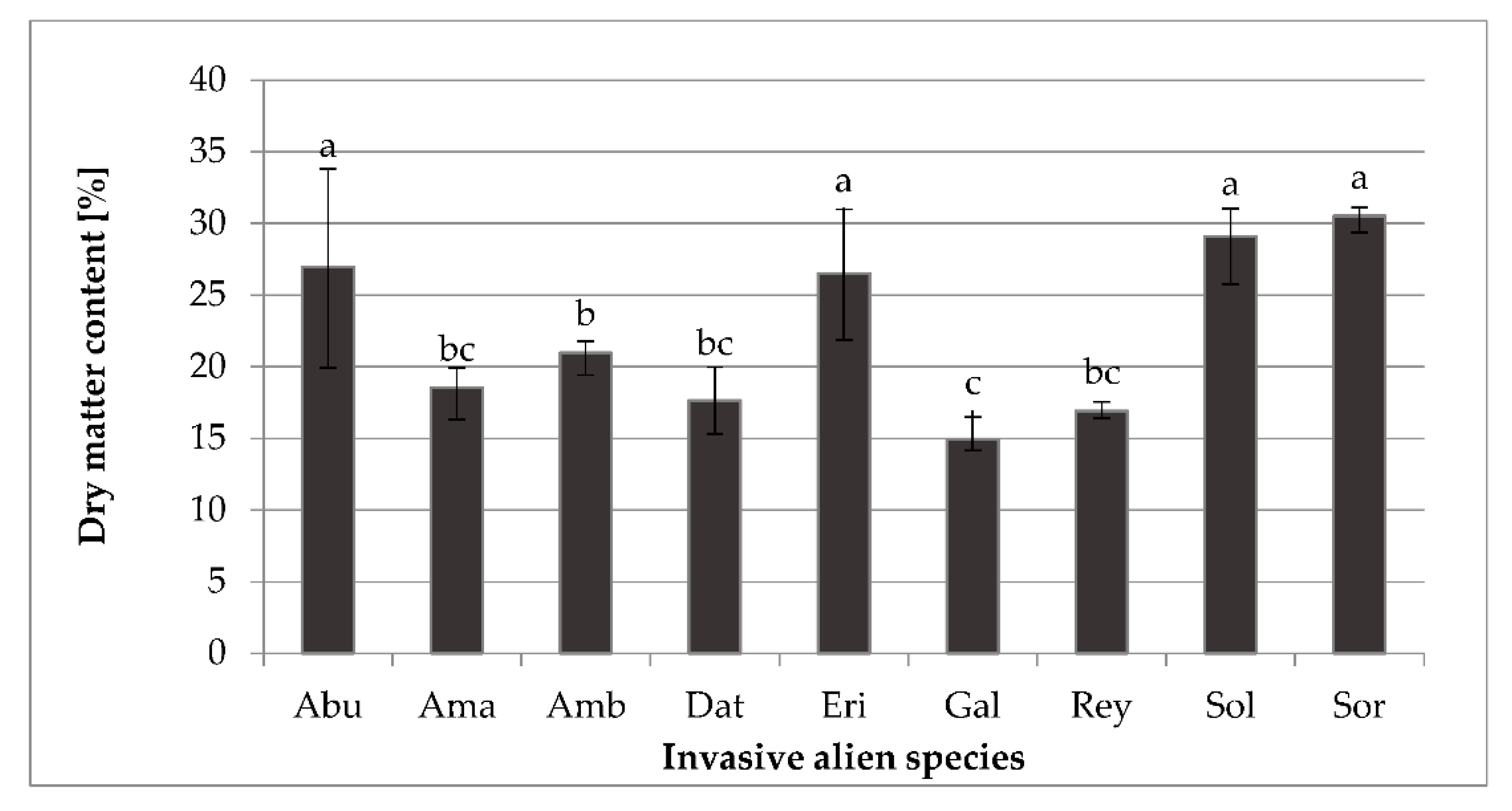
2.1. Bioactive Compounds Content
2.2. Pigment Compounds Content
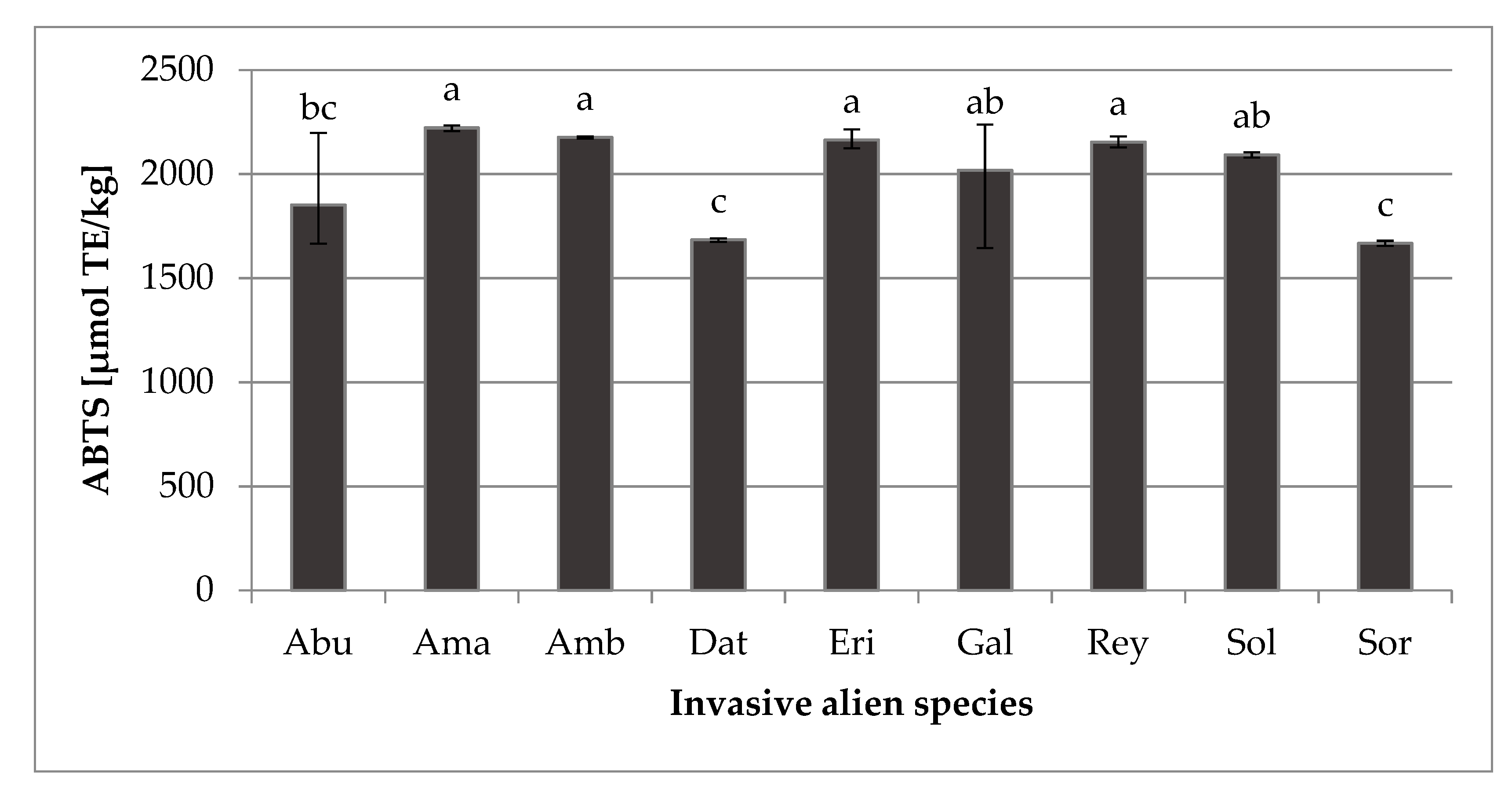
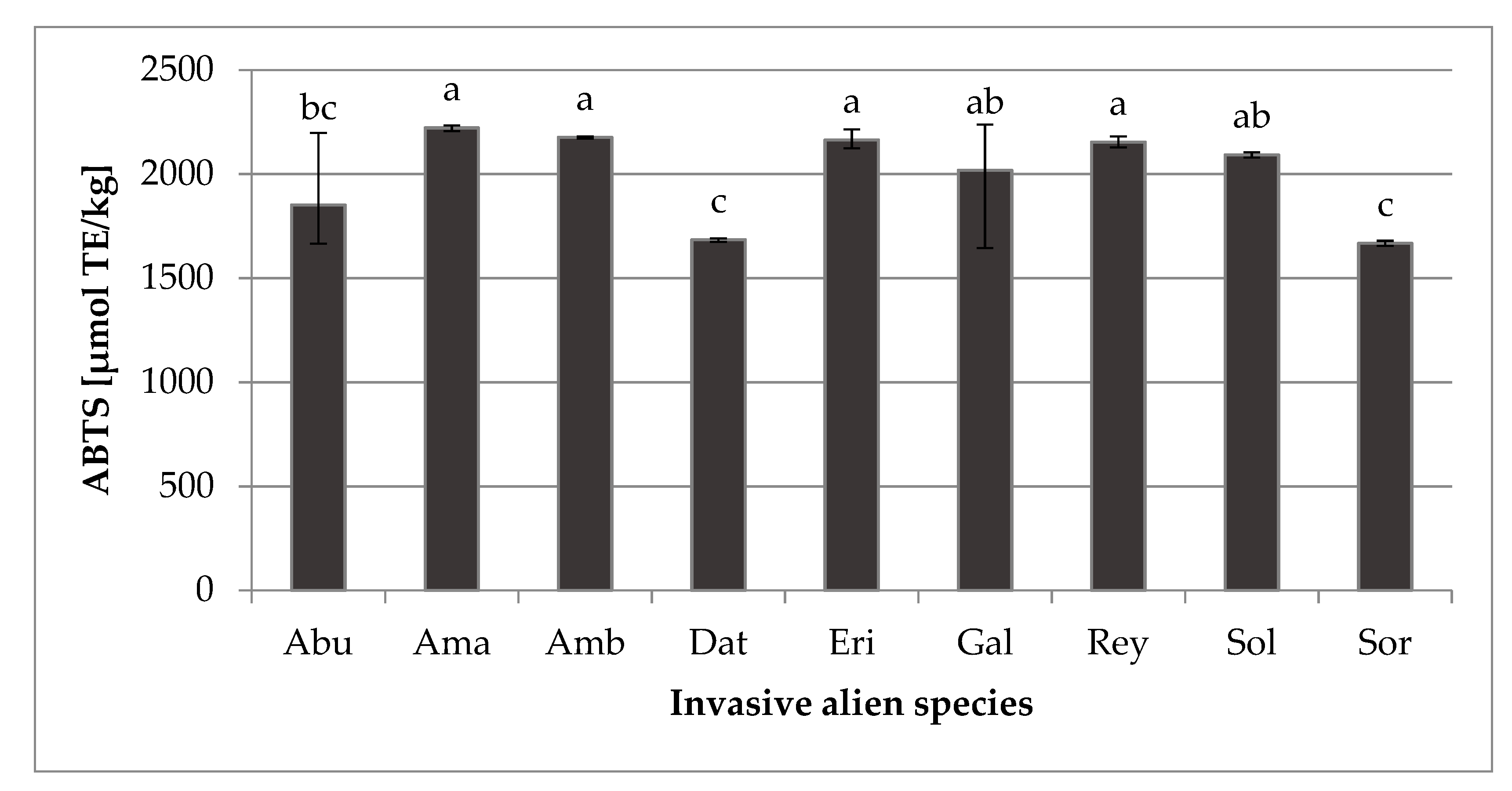
2.3. Antioxidant Capacity (ABTS)
2.4. Correlation Matrix between the Observed Parameters
2.5. Cluster Analysis
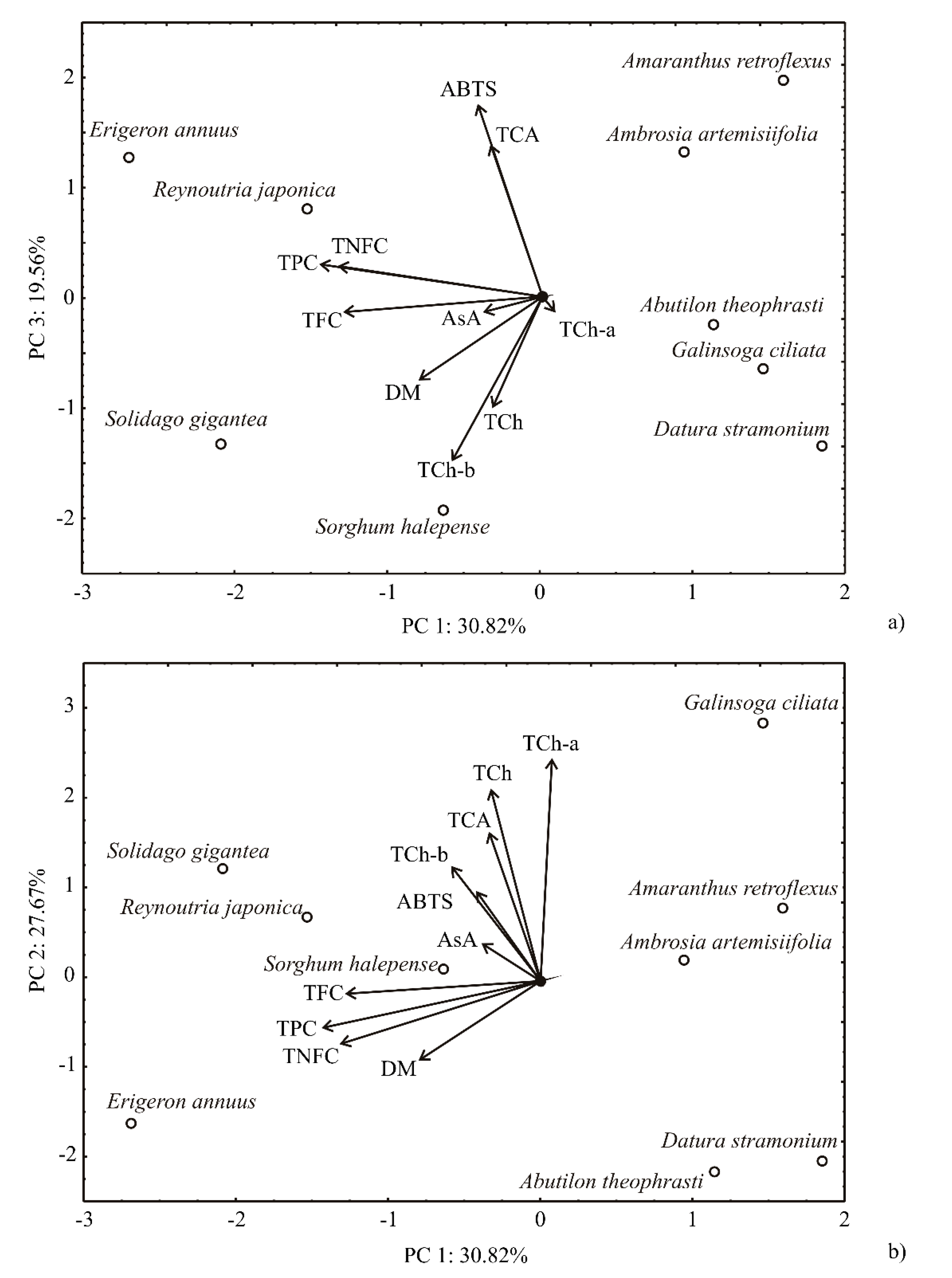
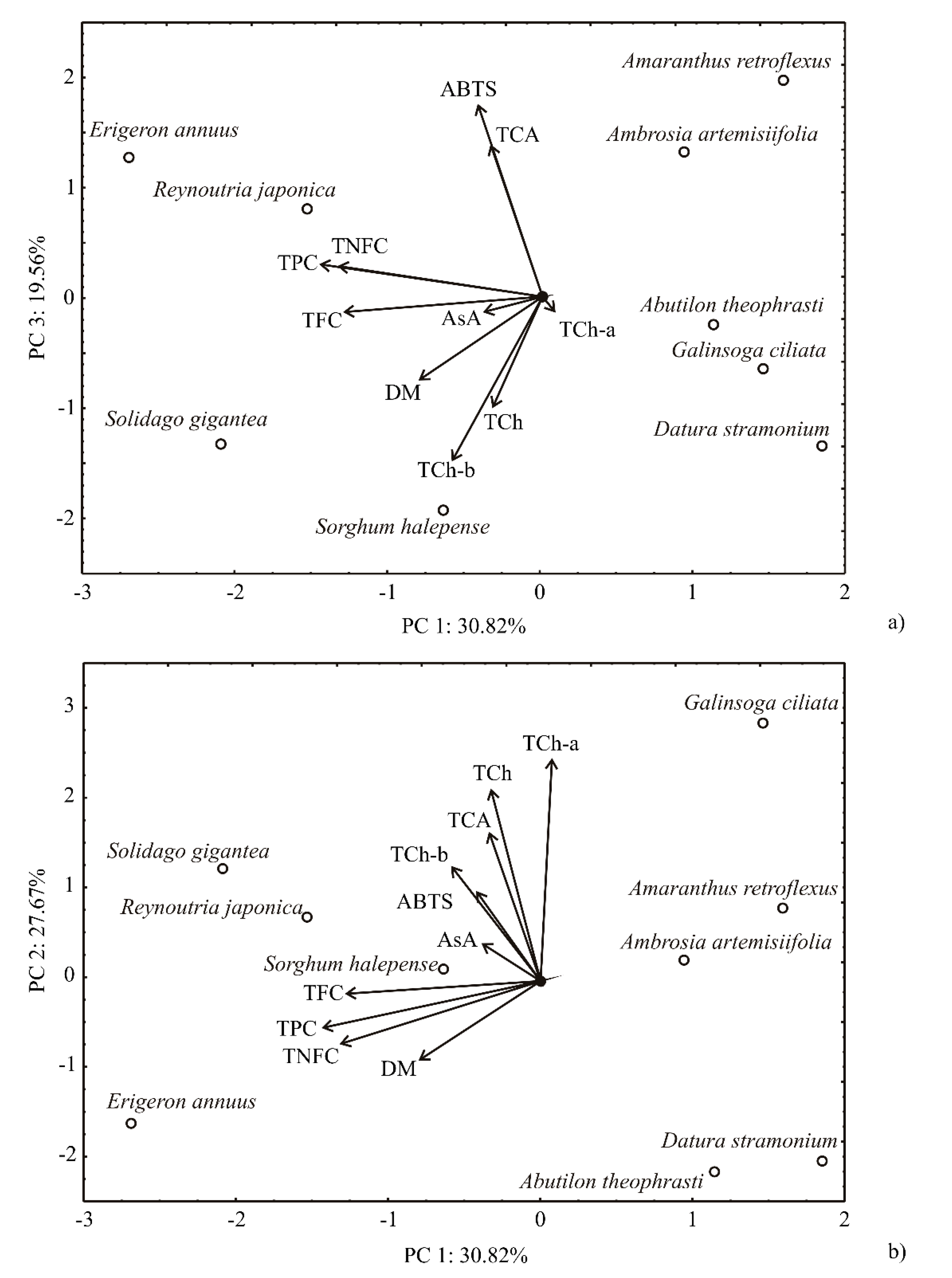
2.6. Principal Component and First-Order Polynomial Model
3. Materials and Methods
3.1. Plant Material
3.2. Determination of Dry Matter Content, Bioactive Compounds Content, and Antioxidant Capacity
3.3. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Secretariat of the Convention on Biological Diversity. Global Biodiversity Outlook 3; UNEP/Earthprint: Montréal, QC, Canada, 2010; pp. 3–73. [Google Scholar]
- Millennium Ecosystem Assessment (Program). Ecosystems and Human Well-Being; Island Press: Washington, DC, USA, 2005; Available online: https://www.millenniumassessment.org/documents/document.766.aspx.pdf (accessed on 15 May 2021).
- IPBES secretariat. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; IPBES: Bonn, Germany; pp. 1–4.
- Herceg, N. Environment and Sustainable Development; Synopsis d.o.o.: Zagreb, Croatia, 2013; pp. 42–386. [Google Scholar]
- Avise, J.C.; Hubbell, S.P.; Ayala, F.J. In the light of evolution II: Biodiversity and extinction. Proc. Natl. Acad. Sci. USA 2008, 105, 11453–11457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IUCN. Red List of Threatened Species. Version 2019-3. 2019. Available online: http://www.iucnredlist.org (accessed on 23 May 2021).
- Walter, K.S.; Gillett, H.J. 1997 IUCN Red List of Threatened Plants; IUCN: Gland, Switzerland; Cambridge, UK, 1998; Available online: https://www.iucnredlist.org/resources/walter1998 (accessed on 23 May 2021).
- Petrović, J.; Ćuričić, S.; Stavretović, N. Invasive Plant Species and Ecological Factors Influencing the Spreading in the Area of the Obrenovački Zabran Nature Monument (Central Serbia). Sumar. List 2016, 140, 45–52. [Google Scholar] [CrossRef]
- Mitić, B.; Boršić, I.; Dujmović, I.; Bogdanović, S.; Milović, M.; Cigić, P.; Rešetnik, I.; Nikolić, T. Alien flora of Croatia: Proposals for standards in terminology, criteria and related database. Nat. Croat. 2008, 17, 73–90. [Google Scholar]
- Barney, J.N.; Tekiela, D.; Dollete, E.; Tomasek, B. What is the “real” impact of invasive plant species? Front. Ecol. Environ. 2013, 11, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Vuković, N. Ekogeografija Invazivne Flore Hrvatske. Ph.D. Thesis, Prirodoslovno-Matematički Fakultetu Zagrebu, Zagreb, Croatia, 2015. [Google Scholar]
- Hulina, N. Korovi; Školska Knjiga: Zagreb, Croatia, 1998; pp. 9–41. [Google Scholar]
- Nikolić, T.; Mitić, B.; Boršić, I. Flora Hrvatske Invazivne Biljke; Alfa d.d.: Zagreb, Croatia, 2014; pp. 7–45. [Google Scholar]
- Novak, N.; Kravarščan, M. Invazivne Strane Korovne Vrste u Republici Hrvatskoj; Hrvatski Centar za Poljoprivredu, Hranu i Selo: Zagreb, Croatia, 2011; pp. 6–35. [Google Scholar]
- Gibert, A.; Gray, E.F.; Westoby, M.; Wright, I.J.; Falster, D.S. On the link between functional traits and growth rate: Meta-analysis shows effects change with plant size, as predicted. J. Ecol. 2016, 104, 1488–1503. [Google Scholar] [CrossRef] [Green Version]
- Moravcová, L.; Pyšek, P.; Jarošík, V.; Pergl, J. Getting the right traits: Reproductive and dispersal characteristics predict the invasiveness of herbaceous plant species. PLoS ONE 2015, 10, e0123634. [Google Scholar] [CrossRef] [Green Version]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? Biol. Invasions 2008, 193, 97–125. [Google Scholar] [CrossRef]
- Vila, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jaros, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pys, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecology 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Bradley, B.A.; Blumenthal, D.M.; Wilcove, D.S.; Ziska, L.H. Predicting plant invasions in an era of global change. Trends Ecol. Evol. 2010, 25, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.A.; Jachetta, J.J.; Brown, M.L.; Carrithers, V.F.; Clark, J.K.; DiTomaso, J.M.; Lym, R.G.; McDaniel, K.C.; Renz, M.J.; Rice, P.M. Assessing the Economic, Environmental, and Societal Losses from Invasive Plants on Rangeland and Wildlands. Weed Technol. 2014, 18 (Suppl. 1), 1411–1416. [Google Scholar] [CrossRef]
- Barokini, T.I.; Babalola, F.D. Management of invasive plant species in Nigeria through economic exploitation: Lessons from other countries. Manag. Biol. Invasions 2012, 3, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Denslow, J. Managing dominance of invasive plants in wild lands. Curr. Sci. 2007, 93, 1579–1586. [Google Scholar]
- Rinella, M.J.; Maxwell, B.D.; Fay, P.K.; Weaver, T.; Sheley, R.L. Control effort exacerbates invasive-species problem. Ecol. Appl. 2009, 19, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.G.; Maxwell, B.D.; Menalled, F.D.; Rew, L.J. Lessons from agriculture may improve the management of invasive plants in wildland systems. Front. Ecol. Environ. 2006, 4, 428–434. [Google Scholar] [CrossRef]
- EC Europa. LIFE Guidelines. Nature and Biodiversity; EC Europa: Brussels, Belgium, 2014; pp. 18–34. Available online: https://ec.europa.eu/environment/archives/life/toolkit/pmtools/life2014_2020/documents/2014nat_application_guide.pdf (accessed on 3 April 2021).
- Van Meerbeek, K.; Appels, L.; Dewil, R.; Calmeyn, A.; Lemmens, P.; Muys, B.; Hermy, M. Biomass of invasive plant species as a potential feedstock for bioenergy production. Biofuels Bioprod. Bioref. 2015, 9, 273–282. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Karimi, M.; Venditti, A. Plants adapted to arid areas: Specialized metabolites. Nat. Prod. Res. 2019, 1–18. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, X.; Liao, Y.; Yang, Z. Roles of specialized metabolites in biological function and environmental adaptability of tea plant (Camellia sinensis) as a metabolite studying model. J. Adv. Res. 2020. [Google Scholar] [CrossRef]
- Parkhomenko, A.Y.; Andreeva, O.A.; Oganesyan, E.T.; Ivashev, M.N. Ambrosia artemisiifolia as a Source of Biologically Active Substances. Pharm. Chem. J. 2005, 39, 37–41. [Google Scholar] [CrossRef]
- Sayyed, A.; Shah, M. Phytochemistry, pharmacological and traditional uses of Datura stramonium L. review. J. Pharmacogn. Phytochem. 2014, 2, 123–125. [Google Scholar]
- Sladonja, B.; Sušek, M.; Guillermic, J. Review on Invasive Tree of Heaven (Ailanthus altissima (Mill.) Swingle) Conflicting Values: Assessment of Its Ecosystem Services and Potential Biological Threat. Environment 2015, 56, 1009–1034. [Google Scholar] [CrossRef]
- Soetan, K.O. Pharmacological and other beneficial effects of anti-nutritional factors in plants—A review. Afr. J. Bot. 2008, 7, 4713–4721. [Google Scholar] [CrossRef]
- Meela, M.M.; Mdee, L.K.; Masoko, P.; Eloff, J.N. Acetone leaf extracts of seven invasive weeds have promising activity against eight important plant fungal pathogens. S. Afr. J. Bot. 2019, 121, 442–446. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of Amaranthus spinosus and Amaranthus viridis weedy species. Sci. Rep. 2019, 9, 20413. [Google Scholar] [CrossRef] [Green Version]
- Reza, M.S.; Ahmed, A.; Caesarendra, W.; Bakar, M.S.A.; Shams, S.; Saidur, R.; Aslfattahi, N.; Azad, A.K. Acacia Holosericea: An Invasive Species for Bio-char, Bio-oil, and Biogas Production. Bioengineering 2019, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Matt, F.; Dietenberger, M.A.; Weise, D. Summative and ultimate analysis of live leaves from southern U.S. forest plants for use in fire modeling. Energ. Fuel 2020, 34, 4703–4720. [Google Scholar] [CrossRef]
- Chew, K.W.; Chia, S.R.; Yen, H.-W.; Nomanbhay, S.; Ho, Y.-C.; Show, P.L. Transformation of Biomass Waste into Sustainable Organic Fertilizers. Sustainability 2019, 11, 2266. [Google Scholar] [CrossRef] [Green Version]
- Luís, Â.; Gil, N.; Amaral, M.E.; Domingues, F.; Duarte, A.P.C. Ailanthus Altissima (Miller) Swingle: A Source of Bioactive Compounds with Antioxidant Activity. BioResources 2012, 7, 2105–2120. [Google Scholar] [CrossRef] [Green Version]
- Pintó-Marijuan, M.; Munné-Bosch, S. Ecophysiology of invasive plants: Osmotic adjustment and antioxidants. Trends Plant Sci. 2013, 18, 660–666. [Google Scholar] [CrossRef]
- Zhou, X.; Gu, Z.; Xu, H.; Chen, L.; Tao, G.; Yu, Y.; Li, K. The Effects of Exogenous Ascorbic Acid on the Mechanism of Physiological and Biochemical Responses to Nitrate Uptake in Two Rice Cultivars (Oryza sativa L.) Under Aluminum Stress. J. Plant Growth Regul. 2016, 35, 1013–1024. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52. [Google Scholar] [CrossRef] [Green Version]
- Barata-Soares, A.D.; Gomez, M.L.; Mesquita, C.H.; Lajolo, F.M. Ascorbic acid biosynthesis: A precursor study on plants. Braz. J. Plant Physiol. 2004, 16, 147–154. [Google Scholar] [CrossRef]
- Bassi, M.; Lubes, G.; Bianchi, F.; Agnolet, S.; Ciesa, F.; Brunner, K.; Guerra, W.; Robatscher, P.; Oberhuber, M. Ascorbic acid content in apple pulp, peel, and monovarietal cloudy juices of 64 different cultivars. Int. J. Food Prop. 2017, 20, S2626–S2634. [Google Scholar] [CrossRef] [Green Version]
- Favell, D.J. A comparison of the vitamin C content of fresh and frozen vegetables. Food Chem. 1998, 62, 59–64. [Google Scholar] [CrossRef]
- Ghani, U. Polyphenols. In Alpha-Glucosidase Inhibitors; Elsevier: Amsterdam, The Netherlands, 2020; pp. 61–100. [Google Scholar] [CrossRef]
- Belščak-Cvitanović, A.; Durgo, K.; Huđek, A.; Bačun-Družina, V.; Komes, D. Overview of polyphenols and their properties. In Polyphenols: Properties, Recovery, and Applications; Woodhead Publishing: Cambridge, UK, 2018; pp. 3–44. [Google Scholar] [CrossRef]
- Čvorović, J.; Ziberna, L.; Fornasaro, S.; Tramer, F.; Passamonti, S. Bioavailability of Flavonoids: The Role of Cell Membrane Transporters. In Polyphenols: Mechanisms of Action in Human Health and Disease; Academic Press: Cambridge, MA, USA, 2018; pp. 295–320. [Google Scholar] [CrossRef]
- Kumar Rai, P.; Singh, J.S. Invasive alien plant species: Their impact on environment, ecosystem services and human health. Ecol. Indic. 2020, 111, 106020. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.M.; Tyler, A.C.; Hellquist, C.E.; Pagano, T. Phenolic content of invasive and non-invasive emergent wetland plants. Aquat. Bot. 2017, 136, 146–154. [Google Scholar] [CrossRef]
- Shah, M.A.R.; Khan, R.A.; Ahmed, M. Phytochemical analysis, cytotoxic, antioxidant and anti-diabetic activities of the aerial parts of Sorghum halepense. Bangladesh. J. Pharmacol. 2019, 14, 144–151. [Google Scholar] [CrossRef]
- Hosikian, A.; Lim, S.; Halim, R.; Danquah, M.K. Chlorophyll Extraction from Microalgae: A Review on the Process Engineering Aspects. Int. J. Chem. Eng. 2010, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Brotosudarmo, T.H.P.; Limantara, L.; Chandra, R.D. Chloroplast Pigments: Structure, Function, Assembly and Characterization. In Plant Growth and Regulation—Alterations to Sustain Unfavorable Conditions; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic Response of Plants Under Different Abiotic Stresses: A Review. J. Plant. Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Galanakis, C.M. Introduction. In Nutraceutical and Functional Food Components; Academic Press: Cambridge, MA, USA, 2017; pp. 1–14. [Google Scholar] [CrossRef]
- Marksa, M.; Zymone, K.; Ivanauskas, L.; Radušienė, J.; Pukalskas, A.; Raudone, L. Antioxidant profiles of leaves and inflorescences of native, invasive and hybrid Solidago species. Ind Crops Prod. 2020, 145, 112123. [Google Scholar] [CrossRef]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of Antioxidant Potential of Plants and its Relevance to Therapeutic Applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [Green Version]
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal prospects of antioxidants: A review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Domac, R. Flora of Croatia; Školska Knjiga: Zagreb, Croatia, 1994. [Google Scholar]
- Nikolić, T. Flora Croatica 4—Vascular Flora of Republic of Croatia; Alfa d.d.: Zagreb, Croatia, 2019. [Google Scholar]
- Nikolić, T. Flora Croatica Database; University of Zagreb, Faculty of Science, Department of Botany: Zagreb, Croatia, 2021; Available online: http://hirc.botanic.hr/fcd/ (accessed on 3 April 2021).
- Ostročki, L. Statistical Yearbook of the Republic of Croatia; Croatian Bureau of Statistics: Zagreb, Croatia, 2018; pp. 1–158. [Google Scholar]
- Bogunović, M.; Vidaček, Ž.; Racz, Z.; Husnjak, S.; Sraka, M. Soil Suitability Map for Cultivation; Soil Science Department of Agricultural Faculty University of Zagreb: Zagreb, Croatia, 1996; Available online: https://esdac.jrc.ec.europa.eu/images/Eudasm/HR/PDF/cro2.pdf (accessed on 15 May 2021).
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1955. [Google Scholar]
- EN ISO 18122:2015. Solid Biofuels. Determination of Ash Content; ISO: Geneva, Switzerland, 2015. [CrossRef]
- EN 15104:2011. Solid Biofuels. Determination of Total Content of Carbon, Hydrogen and Nitrogen—Instrumental Methods; ISO: Geneva, Switzerland, 2011.
- EN 15289:2011. Solid Biofuels. Determination of Total Content of Sulfur and Chlorine; ISO: Geneva, Switzerland, 2011.
- Desta, B.H.; Melese, G.M. Determination of Protein Value and Alcoholic Content in Locally Prepared Different Types of Cheka at Different Stages Using CHNS Elemental Analyzer and Specific Gravity Methods. Sci. Res. J. 2019, 7, 168–174. [Google Scholar] [CrossRef]
- Jones, D.B. Factors for Converting Percentages of Nitrogen in Foods and Feeds into Percentages of Proteins; U.S. Department of Agriculture: Washington, DC, USA, 1931; Volume 183, pp. 1–22. [Google Scholar]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2002. [Google Scholar]
- Ough, C.S.; Amerine, M.A. Methods for Analysis of Musts and Wines; John Wiley & Sons: New York, NY, USA, 1988. [Google Scholar]
- Holm, G. Chlorophyll mutations in barley. Acta Agric. Scand. 1954, 4, 457–471. [Google Scholar] [CrossRef]
- Wettstein, D. Chlorophyll letale und der submikroskopische Formwechsel der Plastiden. Exp. Cell Res. 1957, 12, 427–434. [Google Scholar] [CrossRef]
- Miller, N.J.; Diplock, A.T.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A novel method for measuring antioxidant capacity and its application to monitoring the antioxidant status in premature neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef] [Green Version]
- SAS®/STAT 9.3; SAS Institute Inc.: Cary, NC, USA, 2010.
- STATISTICA 10.0; StatSoft Inc.: Tulsa, OK, USA, 2010.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | AsA [mg/100 g FW] | TPC [mg GAE/100 g FW] | TNFC [mg GAE/100 g FW] | TFC [mg GAE/100 g FW] |
|---|---|---|---|---|
| A. theophrasti | 20.50 ± 4.42 bc | 334.03 ± 80.66 d | 157.18 ± 47.65 cde | 176.86 ± 35.10 d |
| A. retroflexus | 19.51 ± 0.39 bc | 187.15 ± 62.59 de | 82.02 ± 33.26 de | 105.12 ± 29.96 d |
| A. artemisiifolia | 19.64 ± 0.91 bc | 342.21 ± 75.99 d | 162.20 ± 37.94 cde | 180.00 ± 38.27 d |
| D. stramonium | 21.36 ± 3.29 bc | 285.62 ± 73.91 de | 134.99 ± 37.84 de | 150.64 ± 36.41 d |
| E. annuus | 16.00 ± 3.61 c | 1365.92 ± 185.74 a | 878.92 ± 203.57 a | 487.00 ± 24.61 b |
| G. ciliata | 19.13 ± 5.68 bc | 142.66 ± 40.79 e | 42.74 ± 37.57 e | 102.89 ± 40.39 d |
| R. japonica | 38.46 ± 9.39 a | 937.72 ± 75.39 b | 290.11 ± 21.89 c | 647.50 ± 64.44 a |
| S. gigantea | 21.66 ± 4.84 bc | 905.69 ± 95.87 b | 541.56 ± 80.37 b | 364.13 ± 26.12 c |
| S. halepense | 25.08 ± 4.50 b | 506.94 ± 88.83 c | 205.36 ± 59.59 cd | 506.94 ± 88.83 b |
| ANOVA | p ≤ 0.0013 | p ≤ 0.0001 | p ≤ 0.0001 | p ≤ 0.0001 |
| Sample | TCh-a [mg/g] | TCh-b [mg/g] | TCh [mg/g] | TCA [mg/g] |
|---|---|---|---|---|
| A. theophrasti | 0.58 ± 0.21 d | 0.42 ± 0.11 d | 1.00 ± 0.12 d | 0.14 ± 0.13 bc |
| A. retroflexus | 0.87 ± 0.07 ab | 0.35 ± 0.07 d | 1.21 ± 0.13 bcd | 0.26 ± 0.02 a |
| A. artemisiifolia | 0.80 ± 0.16 bc | 0.42 ± 0.17 d | 1.22 ± 0.32 bcd | 0.23 ± 0.03 ab |
| D. stramonium | 0.58 ± 0.05 d | 0.54 ± 0.04 bcd | 1.12 ± 0.09 cd | 0.07 ± 0.01 c |
| E. annuus | 0.63 ± 0.07 cd | 0.50 ± 0.06 cd | 1.13 ± 0.13 cd | 0.21 ± 0.04 ab |
| G. ciliata | 1.05 ± 0.04 a | 0.76 ± 0.20 ab | 1.81 ± 0.21 a | 0.22 ± 0.06 ab |
| R. japonica | 0.82 ± 0.16 bc | 0.53 ± 0.08 cd | 1.35 ± 0.23 bc | 0.21 ± 0.05 ab |
| S. gigantea | 0.85 ± 0.07 b | 0.92 ± 0.04 a | 1.77 ± 0.04 a | 0.19 ± 0.04 ab |
| S. halepense | 0.87 ± 0.07 ab | 0.65 ± 0.20 bc | 1.52 ± 0.27 ab | 0.18 ± 0.05 ab |
| ANOVA | p ≤ 0.0010 | p ≤ 0.0005 | p ≤ 0.0004 | p ≤ 0.0290 |
| AsA | TPC | TNFC | TFC | TCh-a | TCh-b | TCh | TCA | ABTS | |
|---|---|---|---|---|---|---|---|---|---|
| DM | −0.204 | 0.422 | 0.508 | 0.369 | −0.265 | 0.239 | 0.005 | −0.130 | −0.266 |
| AsA | 0.196 | −0.144 | 0.645 ** | 0.144 | 0.054 | 0.113 | 0.012 | −0.012 | |
| TPC | 0.941 ++ | 0.797 * | −0.265 | 0.209 | −0.014 | 0.125 | 0.320 | ||
| TNFC | 0.578 | −0.326 | 0.217 | −0.042 | 0.097 | 0.310 | |||
| TFC | −0.040 | 0.201 | 0.104 | 0.101 | 0.052 | ||||
| TCh-a | 0.458 | 0.832 + | 0.645 | 0.274 | |||||
| TCh-b | 0.875 + | −0.078 | −0.165 | ||||||
| TCh | 0.303 | 0.047 | |||||||
| TCA | 0.821 + |
| Variable | dF | ABTS | p-Value |
|---|---|---|---|
| AsA | 1 | 1597.8 | 0.454 |
| TPC | 1 | 1593.6 | 0.455 |
| TNFC | 1 | 125.5 | 0.82 |
| TFC | 1 | 73,841.0 * | 0.025 |
| TCh | 1 | 2938.8 | 0.338 |
| TCA | 1 | 144,906.0 * | 0.013 |
| Error | 2 | 3766.5 |
| Invasive Alien Plant Species | |||
|---|---|---|---|
| A. theophrasti | |||
| Coordinates | 45° 41′ 33.7812″ N 15° 29′ 24.9684″ E | 45° 39′ 46.3716″ N 15° 35′ 26.8476″ E | 45° 40′ 51.4488″ N 15° 30′ 49.4856″ E |
| Dominant Soil type | Rendzina on marl and soft limestone | Pseudogley on sloping terrain | Rendzina on marl and soft limestone |
| Time of sampling | May | May | May |
| A. retroflexus | |||
| Coordinates | 45° 41′ 33.7812″ N 15° 29′ 24.9684″ E | 45° 44′ 57.9120″ N 15° 23′ 30.6384″ E | 45° 47′ 26.2824″ N 15° 29′ 29.8608″ E |
| Dominant Soil type | Rendzina on marl and soft limestone | Rendzina on marl and soft limestone | Cambisol eutric on flysch or soft limestone |
| Time of sampling | June | June | June |
| A. artemisiifolia | |||
| Coordinates | 45° 41′ 38.9364″ N 15° 29′ 15.6480″ E | 45° 45′ 10.8360″ N 15° 25′ 25.9284″ E | 45° 44′ 18.9564″ N 15° 28′ 14.2284″ E |
| Dominant Soil type | Rendzina on marl and soft limestone | Rendzina on marl and soft limestone | Cambisol eutric on flysch or soft limestone |
| Time of sampling | May | May | May |
| D. stramonium | |||
| Coordinates | 45° 39′ 46.3716″ N 15° 35′ 26.8476″ E | 45° 40′ 49.9620″ N 15° 30′ 39.2364″ E | 45° 40′ 8.9148″ N 15° 34′ 58.2960″ E |
| Dominant Soil type | Pseudogley on sloping terrain | Rendzina on marl and soft limestone | Pseudogley on sloping terrain |
| Time of sampling | April | April | April |
| E. annuus | |||
| Coordinates | 45° 39′ 25.2108″ N 15° 31′ 14.0376″ E | 45° 40′ 45.2532″ N 15° 30′ 47.2896″ E | 45° 40′ 49.8972″ N 15° 30′ 42.5268″ E |
| Dominant Soil type | Calcocambisol on dolomite | Pseudogley on sloping terrain | Rendzina on marl and soft limestone |
| Time of sampling | April | April | April |
| G. ciliata | |||
| Coordinates | 45° 41′ 33.7812″ N 15° 29′ 24.9684″ E | 45° 43′ 16.7520″ N 15° 26′ 12.5808″ E | 45° 40′ 49.9620″ N 15° 30′ 39.2364″ E |
| Dominant Soil type | Rendzina on marl and soft limestone | Rendzina on skeletal limestone | Rendzina on marl and soft limestone |
| Time of sampling | April | April | April |
| R. japonica | |||
| Coordinates | 45° 50′ 3.8400″ N 15° 37′ 3.5616″ E | 45° 50′ 5.7660″ N 15° 36′ 52.3008″ E | 45° 46′ 08.7492” N 15° 24′ 48.9996” E |
| Dominant Soil type | Rendzina, Luvisol, Cambisol on Terra rossa | Rendzina, Luvisol, Cambisol on Terra rossa | Cambisol eutric on flysch or soft limestone |
| Time of sampling | May | May | May |
| S. gigantea | |||
| Coordinates | 45° 40′ 49.8972″ N 15° 30′ 42.5268″ E | 45° 50′ 21.8904″ N 15° 35′ 55.572″ E | 45° 43′ 16.7520″ N 15° 26′ 12.5808″ E |
| Dominant Soil type | Rendzina on marl and soft limestone | Rendzina, Luvisol, Cambisol on Terra rossa | Rendzina on skeletal limestone |
| Time of sampling | June | June | June |
| S. halepense | |||
| Coordinates | 45° 39′ 46.3716″ N 15° 35′ 26.8476″ E | 45° 40′ 51.4488″ N 15° 30′ 49.4856″ E | 45° 40′ 8.9148″ N 15° 34′ 58.2960″ E |
| Dominant Soil type | Pseudogley on sloping terrain | Rendzina on marl and soft limestone | Pseudogley on sloping terrain |
| Time of sampling | April | April | April |
| Average air temperature (°C) | 9.8 | 9.8 | 9.8 |
| Average precipitation (mm) | 1315.2 | 1315.2 | 1315.2 |
| Relative air humidity (%) | 71 | 71 | 71 |
| Number of sunny days | 56 | 56 | 56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peter, A.; Žlabur, J.Š.; Šurić, J.; Voća, S.; Purgar, D.D.; Pezo, L.; Voća, N. Invasive Plant Species Biomass—Evaluation of Functional Value. Molecules 2021, 26, 3814. https://doi.org/10.3390/molecules26133814
Peter A, Žlabur JŠ, Šurić J, Voća S, Purgar DD, Pezo L, Voća N. Invasive Plant Species Biomass—Evaluation of Functional Value. Molecules. 2021; 26(13):3814. https://doi.org/10.3390/molecules26133814
Chicago/Turabian StylePeter, Anamarija, Jana Šic Žlabur, Jona Šurić, Sandra Voća, Dubravka Dujmović Purgar, Lato Pezo, and Neven Voća. 2021. "Invasive Plant Species Biomass—Evaluation of Functional Value" Molecules 26, no. 13: 3814. https://doi.org/10.3390/molecules26133814
APA StylePeter, A., Žlabur, J. Š., Šurić, J., Voća, S., Purgar, D. D., Pezo, L., & Voća, N. (2021). Invasive Plant Species Biomass—Evaluation of Functional Value. Molecules, 26(13), 3814. https://doi.org/10.3390/molecules26133814