
Phytochemical Characterization and Bioactivity of Asparagus acutifolius: A Focus on Antioxidant, Cytotoxic, Lipase Inhibitory and Antimicrobial Activities


 ,
,  ,
, 
Abstract

1. Introduction
2. Results and Discussion
2.1. Analysis of Phenolic Compounds
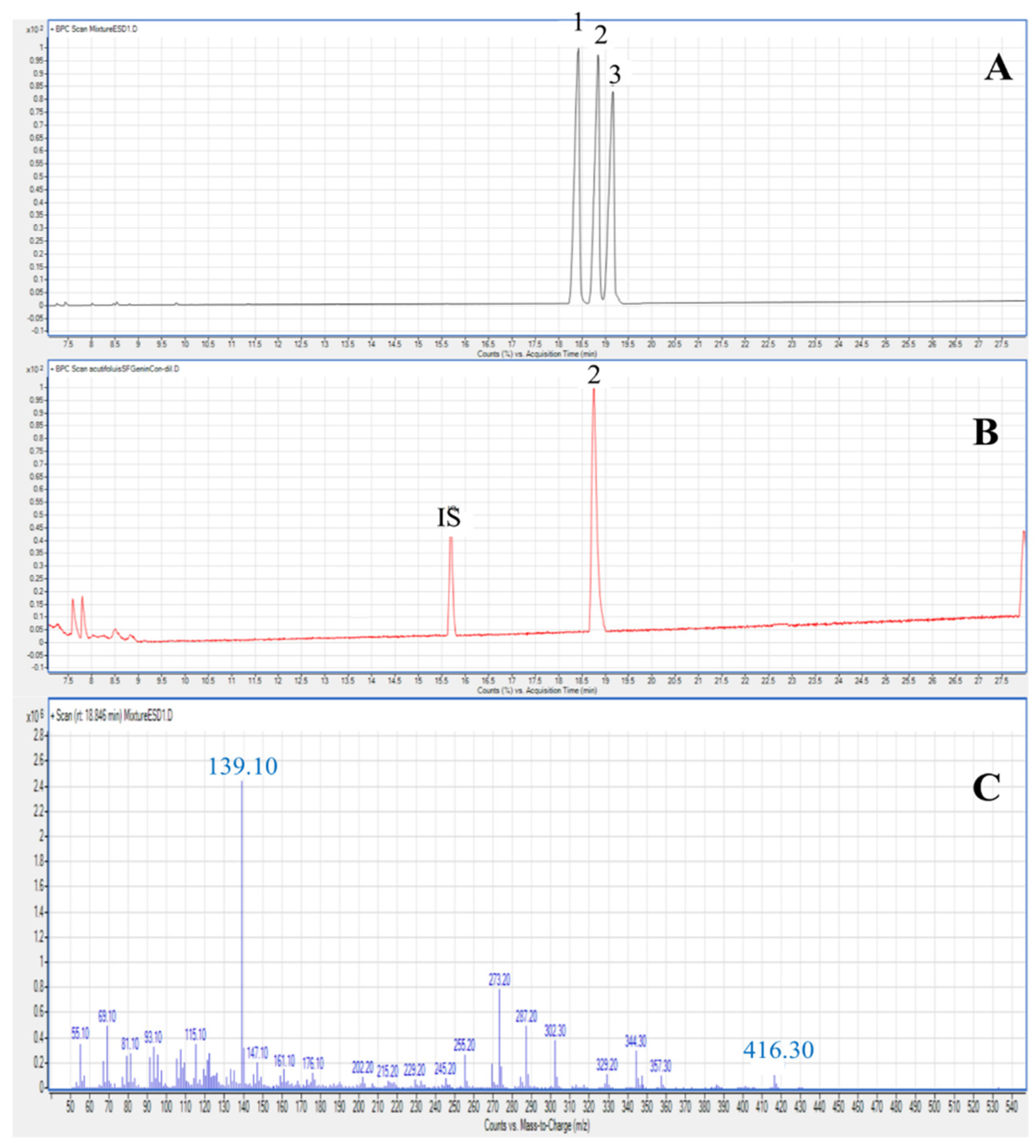
2.2. Saponins in the Different Plant Parts of A. acutifolius
2.3. Evaluation of Antioxidant Capacity
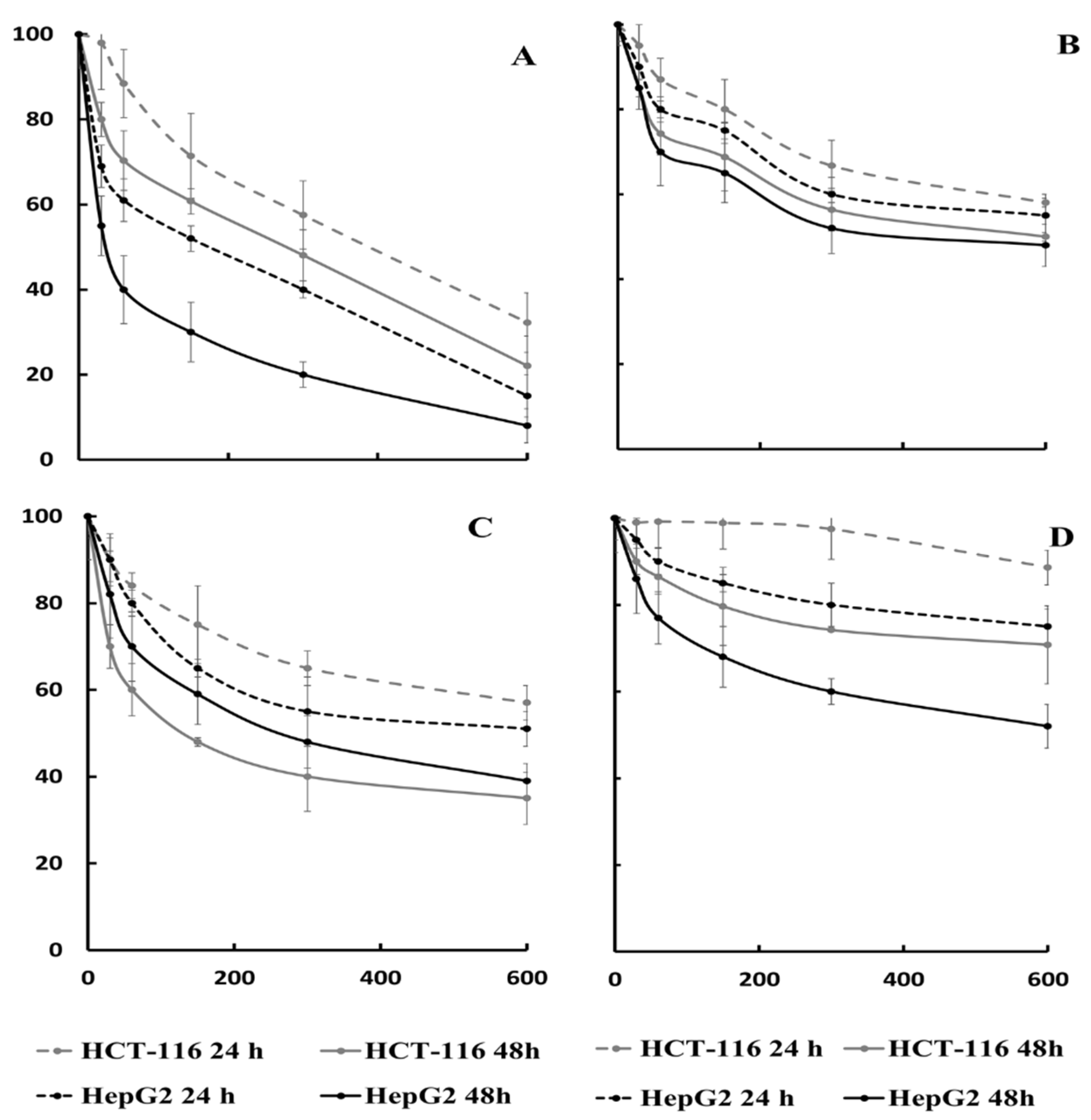
2.4. Cytotoxic Activity
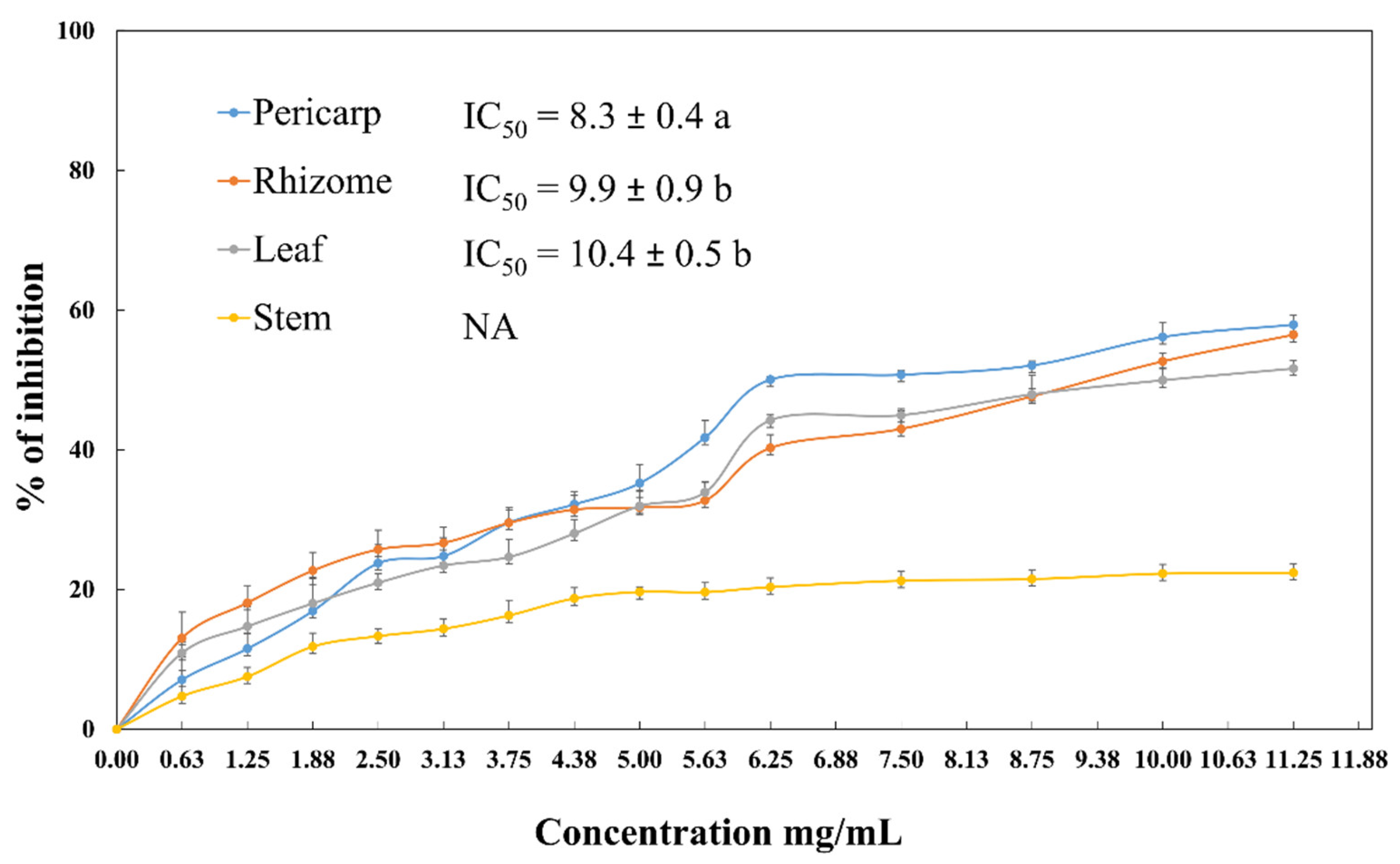
2.5. Lipase Inhibitory Activity
2.6. Antimicrobial Activity
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material
3.3. Preparation of Ethanolic Extract
3.4. Analysis of Phenolic Compounds
3.4.1. Extraction and Quantification of Free Phenolics
3.4.2. Analysis and Quantification of Flavonoids by HPLC–DAD
3.5. Analysis of Saponins
3.5.1. Characterization and Quantification of Saponins by LC-MS
3.5.2. Determination of Exact Mass
3.5.3. Preparation of Saponin Extract and Purification of Individual Saponins
3.5.4. Hydrolysis of Saponins
3.6. Determination of Antioxidant Capacity
3.7. Cytotoxicity against Cancer Cell Lines
3.8. Pancreatic Lipase Inhibitory Activity Assay In Vitro
3.9. Antimicrobial Activity
3.9.1. Microbial Strains
3.9.2. Well Diffusion Method
3.9.3. MIC and MBC Determinations for Bacteria
3.9.4. MIC and MFC Determinations for Fungal
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Barucha, Z.; Pretty, J. The roles and values of wild foods in agriculture systems. Phil. Trans. R. Soc. B. 2010, 365, 2913–2926. [Google Scholar] [CrossRef]
- Sánchez-Mata, M.C.; Cabrera, R.D.; Morales, P.; Fernández-Ruiz, V.; Cámara, M.; Díez, C.; Pardode-Santaya, M.; Tardío, J. Wild vegetables of the Mediterranean area as valuable sources of bioactive compounds. Genet. Resour. Crop. Evol. 2011, 59, 431–443. [Google Scholar] [CrossRef]
- Guarrera, P.M.; Savo, V. Perceived health properties of wild and cultivated food plants in local and popular traditions of Italy: A review. J. Ethnopharmacol. 2013, 146, 659–680. [Google Scholar] [CrossRef]
- García-Herrera, P.; Morales, P.; Cámara, M.; Fernández-Ruiz, V.; Tardío, J.; Cortes Sánchez-Mata, M. Nutritional and Phytochemical Composition of Mediterranean Wild Vegetables after Culinary Treatment. Foods 2020, 9, 1761. [Google Scholar] [CrossRef]
- Sergio, L.; Boari, F.; Pieralice, M.; Linsalata, V.; Cantore, V.; Di Venere, D. Bioactive Phenolics and Antioxidant Capacity of Some Wild Edible Greens as Affected by Different Cooking Treatments. Foods 2020, 9, 1320. [Google Scholar] [CrossRef] [PubMed]
- Bilušić, T.; Šola, I.; Rusak, G.; Poljuha, D.; Čikeš Čulić, V. Antiproliferative and pro-apoptotic activities of wild asparagus (Asparagus acutifolius L.), black bryony (Tamus communis L.) and butcher’s broom (Ruscus aculeatus L.) aqueous extracts against T24 and A549 cancer cell lines. J. Food Biochem. 2019, 43, e12781. [Google Scholar] [CrossRef] [PubMed]
- Di Maro, A.; Pacifico, S.; Fiorentino, A.; Galasso, S.; Gallicchio, M.; Guida, V.; Severino, V.; Monaco, P.; Parente, A. Raviscanina wild asparagus (Asparagus acutifolius L.): A nutritionally valuable crop with antioxidant and antiproliferative properties. Food Res. Int. 2013, 53, 180–188. [Google Scholar] [CrossRef]
- Sautour, M.; Miyamoto, T.; Lacaille-Dubois, M.A. Stereoidal saponins from Asparagus acutifolius. Phytochemistry 2007, 68, 2554–2562. [Google Scholar] [CrossRef]
- Maccioni, S.; Monti, G.; Flamini, G.; Cioni, P.L.; Morelli, I.; Guazzi, E. Ethnobotanical researches in liguria. the lerrone and the Lower Arroscia Valleys. Atti Soc. Tosc. Sci. Nat. Mem. Ser. B 2004, 111, 129–134. [Google Scholar]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.H. Effect of extraction solvent on total phenol content total flavonoid content and antioxidant activity of Limnophila aromatic. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef]
- Makris, D.P.; Rossiter, J.T. Domestic processing of onion bulbs and asparagus spears: Effect of flavonol content and antioxidant status. J. Agric. Food Chem. 2001, 49, 3216–3222. [Google Scholar] [CrossRef]
- Guillén, R.; Rodríguez, R.; Jaramillo, S.; Rodríguez, G.; Espejo, J.A.; Fernández-Bolaños, J.; Heredia, A.; Jiménez, A. Antioxidants from asparagus spears: Phenolics. Acta Hortic. 2008, 776, 247–253. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, J.H.; Yo, I.H.; Gorinstein, S.; Bae, J.H.; Ku, Y.G. Bioactive compounds, antioxidant and binding activities and spear yield of Asparagus officinalis L. Plant. Foods Hum. Nutr. 2014, 69, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Solana, M.; Boschiero, I.; Dall’Acqua, S.; Bertucco, A. A comparison between supercritical fluid and pressurized liquid extraction methods for obtaining phenolic compounds from Asparagus officinalis L. J. Supercrit Fluids 2015, 100, 201–208. [Google Scholar] [CrossRef]
- García Giménez, M.D.; De la Puerta, R.; Sáenz, M.T.; Marquez-Martín, A.; Fernández-Arche, M.A. Hypocholesterolemic and hepatoprotective effects of “Triguero” asparagus from Andalusian in rats fed a high cholesterol diet. Evid. Based Complementary Altern. Med. 2012, 814752. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Zhao, J.; Zhang, W.; Pang, X. Saponins extracted from by-product of Asparagus officinalis L. suppress tumour cell migration and invasion through targeting Rho GTPase signalling pathway. J. Sci. Food Agric. 2013, 93, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Palfi, M.; Jurković, Z.; Ćosić, J.; Tomić-Obrdalj, H.; Jurković, V.; Knežević, N.; Vrandečić, K. Total polyphenol content and antioxidant activity of wild and cultivated asparagus in Croatia. Poljoprivreda 2017, 23, 56–62. [Google Scholar] [CrossRef]
- Salvatore, S.; Pellegrini, N.; Brenna, O.V.; Del Rio, D.; Frasca, G.; Brighenti, F.; Tumino, R. Antioxidant characterization of some Sicilian edible wild greens. J. Agric. Food Chem. 2005, 53, 9465–9471. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Dueñas, M.; Ferreira, I.C.F.R.; Carvalho, A.M.; Santos-Buelga, C. Use of HPLC-DAD-ESI/MS to profile phenolic compounds in edible wild greens from Portugal. Food Chem. 2011, 127, 169–173. [Google Scholar] [CrossRef]
- Ferrara, L.; Dosi, R.; Di Maro, A.; Guida, V.; Cafarelli, G.; Pacifico, S.; Mastellone, C.; Fiorentino, A.; Rosati, A.; Parente, A. Nutritional values, metabolic profile and radical scavenging capacities of wild asparagus (A. acutifolius L.). J. Food Compos. Anal. 2011, 24, 326–333. [Google Scholar] [CrossRef]
- Lake, R.; Falloon, P.; Cook, D. Replant problem and chemical components of asparagus roots. N. Z. J. Crop. Hortic. Sci. 1993, 21, 53–58. [Google Scholar] [CrossRef]
- Symes, A.; Shavandi, A.; Zhang, H.; Mohamed Ahmed, I.A.; Al-Juhaimi, F.Y.; Alaa El-Bekhit, D.A. Antioxidant Activities and Caffeic Acid Content in New Zealand Asparagus (Asparagus officinalis) Roots Extracts. Antioxidants 2018, 7, 52. [Google Scholar] [CrossRef]
- Billau, W.; Buchloh, G.; Geiger, H.; Hartmann, H.D. HPLC analysis of phenolic compounds in storage roots of white asparagus (Asparagus officinalis L.) during the vegetation-period. Asparagus Res. Newslett. 1987, 5, 18–19. [Google Scholar]
- Hassan, R.A.; Tawfeek, W.A.; Habeeb, A.A.; Mohamed, M.S.; Abdelshafeek, K.A. Investigation of some chemical constituents and antioxidant activity of Asparagus sperngeri. Int. J. Pharm. Pharm. Sci. 2014, 6, 46–51. [Google Scholar]
- Vázquez-Castilla, S.; Jaramillo-Carmona, S.; Fuentes-Alventosa, J.M.; Jiménez-Araujo, A.; Rodríguez-Arcos, R.; Cermeño-Sacristán, P.; Espejo-Calvo, J.A.; Guillén-Bejarano, R. Optimization of a method for the profiling and quantification of saponins in different green asparagus genotypes. J. Agric. Food Chem. 2013, 61, 6250–6258. [Google Scholar] [CrossRef]
- Vázquez-Castilla, S.; Jaramillo-Carmona, S.; Fuentes-Alventosa, J.M.; Jiménez-Araujo, A.; Rodríguez-Arcos, R.; Cermeño-Sacristán, P.; Espejo-Calvo, J.A.; Guillén-Bejarano, R. Saponin Profile of green asparagus genotypes. J. Agric. Food Chem. 2013, 61, 11098–11108. [Google Scholar] [CrossRef]
- Jaramillo-Carmona, S.; Rodriguez-Arcos, R.; Jiménez-Araujo, A.; López, S.; Gil, J.; Moreno, R.; Guillén-Bejarano, R. Saponin Profile of Wild Asparagus Species. Food Sci. 2017, 82, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Sauvaire, Y.; Ribes, G.; Baccou, J.C.; Loubatieres-Mariani, M.M. Implication of steroid saponins and sapogenins in the hypocholesterolemic effect of fenugreek. Lipids 1991, 26, 191–197. [Google Scholar] [CrossRef]
- Zhu, G.L.; Hao, Q.; Li, R.T.; Li, H.Z. Steroidal Saponins from the Roots of Asparagus Cochinchinensis. Chin. J. Nat. Med. 2014, 12, 213–217. [Google Scholar] [CrossRef]
- Podolak, I.; Hubicka, U.; Żuromska-Witek, B.; Janeczko, Z.; Krzek, J. Quantification of saponins in different plant parts of Lysimachia L. Species by validated HPTLC-densitometric method. J. Planar Chromatogr. Mod. TLC 2013, 26, 248–253. [Google Scholar] [CrossRef]
- Hamdi, A.; Jaramillo-Carmona, S.; SrairiBeji, R.; Tej, R.; Zaouia, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Kasri, M.; Lachaal, M.; KarrayBouraoui, N.; et al. The phytochemical and bioactivity profiles of wild Asparagus albus L. plant. Int. Food Res. J. 2017, 99, 720–729. [Google Scholar] [CrossRef]
- Papadopoulou, K.; Melton, R.E.; Leggettt, M.; Daniels, M.J.; Osbourn, A.E. Compromised disease resistance in saponin-deficient plants. Proc. Natl. Acad. Sci. USA 1999, 96, 12923–12928. [Google Scholar] [CrossRef] [PubMed]
- Phrompittayarat, W.; Jetiyanon, K.; Wittaya-Areekul, S.; Putalun, W.; Tanaka, H.; Khan, I.; Ingkaninan, K. Influence of seasons, different plant parts, and plant growth stages on saponin quantity and distribution in Bacopa monnieri. Warasan Songkhla Nakharin. 2011, 33, 193–199. [Google Scholar]
- Jaramillo, S.; Muriana, F.J.G.; Guillen, R.; Jimenez-Araujo, A.; Rodriguez-Arcos, R.; Lopez, S. Saponins from edible spears of wild asparagus inhibit AKT, p70S6K, and ERK signalling, and induce apoptosis through G0/G1 cell cycle arrest in human colon cancer HCT-116 cells. J. Funct. Foods 2016, 26, 1–10. [Google Scholar] [CrossRef]
- Bousserouel, S.; Le Grandois, J.; Gossé, F.; Werner, D.; Barth, S.W.; Marchioni, E.; Marescaux, J.; Raul, F. Methanolic extract of white asparagus shoots activates TRAIL apoptotic death pathway in human cancer cells and inhibits colon carcinogenesis in preclinical model. Int. J. Oncol. 2013, 43, 394–404. [Google Scholar] [CrossRef]
- Khan, K.M.; Nahar, L.; Mannan, A.; Arfan, M.; Khan, G.A.; Al-Groshi, A.; Evans, A.; Dempster, N.M.; Ismail, F.M.D.; Sarker, S.D. Liquid chromatography mass spectrometry analysis and cytotoxicity of Asparagus adscendens roots against human cancer cell lines. Pharmacogn. Mag. 2017, 13, S890–S894. [Google Scholar]
- Liu, W.; Ning, R.; Chen, R.N.; Huang, X.F.; Dai, Q.S.; Hu, J.H.; Wang, Y.W.; Wu, L.L.; Xiong, J.; Hu, G.; et al. Aspafilioside B induces G2/M cell cycle arrest and apoptosis by up-regulating H-Ras and N-Ras via ERK and p38 MAPK signaling pathways in human hepatoma HepG2 cells. Mol. Carcinog. 2015, 55, 440–457. [Google Scholar] [CrossRef]
- Visavadiya, N.P.; Narasimhacharya, A.V. Asparagus root regulates cholesterol metabolism and improves antioxidant status in hypercholesterolemic rats. Evid. Based Compl. Alt. Med. 2007, 6, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhang, W.; Zhao, J.; Wang, J.; Qu, W. Hypolipidaemic and hepatoprotective effects of ethanolic and aqueous extracts from Asparagus officinalis L. by-products in mice fed a high-fat diet. J. Sci. Food Agric. 2010, 90, 1129–1135. [Google Scholar] [CrossRef]
- Weidner, S.; Paproka, J.; Lukaszewicz, D. Change in free, sterified and glycosidic phenolic acids in cereal grains during the after-ripening. Seed Sci. Technol. 1996, 24, 107–114. [Google Scholar]
- Sosulski, F.; Krygier, K.; Hogge, L. Free, esterified, and insoluble- bound phenolic acids 3. Composition of phenolic acids in cereal and potatoe flours. J. Agric. Food Chem. 1982, 30, 337–340. [Google Scholar] [CrossRef]
- Fuentes-Alventosa, J.M.; Rodríguez, G.; Cermeño-Sacristán, P.; Jiménez, A.; Guillén, R.; Fernández-Bolaños, J.; Rodríguez-Arcos, R. Identification of flavonoid diglycosides in several genotypes of asparagus from the Huétor Tájar population variety. J. Agric. Food Chem. 2007, 55, 10028–10035. [Google Scholar] [CrossRef] [PubMed]
- De Ruiter, J.; Burns, J. Characterization of trifluoroacetic acid hydrolyzed subtropical forage grass cell walls. J. Agric. Food Chem. 1987, 4, 308–316. [Google Scholar] [CrossRef]
- Englyst, H.N.; Cummings, J.H. Simplified method for the measurement of total non-starch polysaccharides by gas-liquid chromatography of constituent sugars as alditol acetates. Analyst 1984, 109, 937–942. [Google Scholar] [CrossRef]
- Rodriaguez, R.; Jaramillo, S.; Rodriaguez, G.; Espejo, J.A.; Guillean, R.; Fernaandez-Bolanos, J.; Heredia, A.; Jimeanez, A. Antioxidant Activity of Ethanolic Extracts from Several Asparagus Cultivars. J. Agric. Food Chem. 2005, 53, 5212–5217. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, B.; Falco, V.; Moutinho-Pereira, J.; Bacelar, E.; Peixoto, F.; Correia, C. Effects of elevated CO2 on grapevine (Vitis Vinifera L.): Volatile composition, phenolic content, and in vitro antioxidant activity of red wine. J. Agric. Food Chem. 2009, 57, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Senent, F.; Rodríguez-Gutiérrez, G.; Lama-Muñoz, A.; Fernández-Bolaños, J. New phenolic compounds hydrothermally extracted from the olive oil by-product alperujo and their antioxidative activities. J. Agric. Food Chem. 2012, 60, 1175–1186. [Google Scholar] [CrossRef]
- Psarra, E.; Makris, D.P.; Kallithraka, S.; Kefalas, P. Evaluation of the antiradical and reducing properties of selected Greek whitewines: Correlation with polyphenolic composition. J. Sci. Food Agric. 2002, 82, 1014–1020. [Google Scholar] [CrossRef]
- Kim, J.; Jang, D.S.; Kim, H.; Kim, J.S. Anti-lipase and lipolytic activities of ursolic acid isolated from the roots of Actinidia arguta. Arch. Pharm. Res. 2009, 32, 983–987. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolics | Leaf | Stem | Pericarp | Rhizome |
|---|---|---|---|---|
| Gallic acid | 36.63 ± 0.54 a | nd | 36.24 ± 0.07 a | 36.81 ± 0.73 a |
| Protocatechuic acid | 14.62 ± 0.06 a | 25.92 ± 0.27 d | 18.63 ± 0.07 c | 16.51 ± 0.06 b |
| p-hydroxybenzoic acid | 23.51 ± 0.04 b | 15.26 ± 0.09 a | 25.05 ± 0.10 c | 36.72 ± 0.18 d |
| Vanillic acid | 8.94 ± 0.07 b | 10.61 ± 0.05 c | 3.70 ± 0.10 a | 41.11 ± 0.02 d |
| Caffeic acid | 39.42 ± 0.11 a | 297.05 ± 5.89 d | 67.78 ± 0.33 c | 48.82 ± 0.26 b |
| p-hydroxybenzaldehyde | 11.36 ± 0.09 b | 9.97 ± 0.22 a | 12.26 ± 0.14 c | 17.67 ± 0.09 d |
| Syringic acid | 9.36 ± 0.05 b | 0.67 ± 0.14 a | nd | 37.98 ± 0.20 c |
| Vanillin | 13.95 ± 0.09 b | 12.07 ± 0.05 a | nd | 12.76 ± 0.65 a |
| p-Coumaric acid | 2.95 ± 0.24 a | 6.79 ± 0.13 b | 13.58 ± 0.10 c | 15.34 ± 0.12 d |
| t-ferulic acid | 34.73 ± 0.11 c | 27.53 ± 0.02 a | 30.06 ± 0.11 b | 34.90 ± 0.17 c |
| t-Sinapic acid | 9.75 ± 0.11 b | 9.11 ± 0.04 a | 9.59 ± 0.22 b | 13.17 ± 0.05 c |
| Rutin | nd | 316.27 ± 31.63 | nd | nd |
| Narcissin | nd | 154.10 ± 6. 86 | nd | nd |
| Quercetin | nd | nd | 9.08 ± 0.20 a | 9.60 ± 0.09 b |
| Naringenin | 22.89 ± 0.29 c | 61.27 ± 0.09 d | 1.51 ± 0.05 a | 4.88 ± 0.34 b |
| Kaempferol | 22.08 ± 0.22 b | 4.94 ± 0.15 a | nd | 27.06 ± 0.11 c |
| Total simple phenolics | 205.23 ± 1.28 a | 414.99 ± 6.90 d | 216.88 ± 1.25 b | 311.80 ± 1.23 c |
| Total flavonoids | 44.98 ± 0.51 c | 536.55 ± 25.01 d | 10.59 ± 0.26 a | 41.54 ± 0.54 b |
| Total phenolics | 250.20 ± 1.79 b | 951.54 ± 31.91 d | 227.47 ± 1.50 a | 353.34 ± 1.77 c |
| Molecular Ion (m/z) | Ion Fragmentation | ||||||
|---|---|---|---|---|---|---|---|
| Saponin | Rt a (min) | Formula | [M − H]-Experimental | [M − H]- Theoretical | Error (ppm) | Neg Mode | Pos Mode |
| HTSAP-1 | 24.9 | C50H83O23 | 1051.5331 | 1051.5284 | 4.4 | b Pen919-c Hex757-Hex595-Hex433 | [-Na-H2O]1035-Hex873-Hex711-Pen579-Hex417 e |
| ACSAP-1 | 27.8 | C50H83O23 | 1051.5331 | 1051.5306 | 2.4 | Pen919-Hex757-Hex595-Hex433 | [-Na-H2O]1035-Hex873-Hex711-Pen579-Hex417 |
| HTSAP-3 | 29.6 | C49H83O22 | 1021.5225 | 1021.5212 | 1.2 | Pen889-Pen757-Hex595-Hex433 | [-Na-H2O]1005-Pen873-Hex711-Pen579-Hex417 |
| HTSAP-6 | 32.9 | C50H83O23 | 1035.5381 | 1035.5383 | 0.1 | Pen903-d DoHex757-Hex595-Hex433 e | [-Na-H2O]1019-DoHex873-Pen741-Hex579-Hex417 |
| ACSAP-2 | 42.2 | C44H71O18 | 887.4646 | 887.4601 | 5 | Pen755-Hex593-Hex431 e | [-Na-H2O]871-Hex709-Pen577-Hex415 e |
| ACSAP-3 | 44.1 | C43H69O17 | 857.4540 | 857.4503 | 4.3 | Pen725-Pen593-Hex431 | [-Na-H2O]841-Pen709-Pen577-Hex415 |
| Saponin | Leaf | Stem | Pericarp | Rhizome |
|---|---|---|---|---|
| HTSAP-1 | 530.4 ± 8.1 7% | 254.3 ± 27.6 18% | 215.5 ± 4.5 2% | 8355.0 ± 607.8 24% |
| ACSAP-1 | 882.1 ± 37.6 12% | 220.0 ± 2.7 16% | 2988.3 ± 132.1 25% | nd |
| HTSAP-3 | 353.7 ± 2.3 5% | 96.3 ± 5.6 7% | 509.9 ± 51.5 4% | 16,273.6 ± 493.0 47% |
| HTSAP-6 | 5354.1 ± 10.8 75% | 899.5 ± 48.2 63% | 6935.1 ± 348.3 59% | 7169.2 ± 157.7 21% |
| ACSAP-2 | nd | nd | nd | 928.9 ± 39.6 3% |
| ACSAP-3 | nd | nd | nd | 1704.02 ± 134.3 5% |
| Total saponins | 7120.4 ± 20.9 b | 1418.6 ± 72.9 a | 10,648.7 ± 563.4 c | 34,430.8 ± 1353.2 d |
| Zone of Inhibition (mm) | ||||
|---|---|---|---|---|
| Bacteria (Clinical Isolate) | Ethanolic Extract | Standard Antibiotics | ||
| Leaf | Rhizome | Gent | Strepto | |
| Gram+ | ||||
| Staphylococcus aureus | 16 ± 4.3 a | 0 ± 0 | 26 ± 1.6 c | 21 ± 0.0 b |
| Methicillin resistant Staphylococcus aureus (MR) | 18 ± 1.6 c | 13 ± 1.2 a | 16 ± 1.6 b | 21 ± 0.0 d |
| Staphylococcus epidermidis | 14 ± 2.8 a | 13 ± 1.4 a | 36 ± 0.0 c | 25 ± 0.0 b |
| Enterococcus faecalis | 17 ± 2.8 c | 14 ± 4.3 b | 12 ± 0.0 a | 0 ± 0 |
| Enterococcus faecium | 17 ± 7.1 a | 20 ± 7.1 b | 45 ± 0.0 d | 30 ± 0.0 c |
| Listeria monocytogenes | 17 ± 3.2 c | 0 ± 0 | 14 ± 0.0 b | 12 ± 0.0 a |
| Gram- | ||||
| Klebsiella pneumonia | 14 ± 2.8 b | 11 ± 00 a | 35 ± 0.0 d | 30 ± 0.0 c |
| Multidrug resistant Klebsiella pneumoniae (MDR) | 18 ± 2.8 b | 12 ± 1.6 a | 21± 0.0 c | 0 ± 0 |
| Salmonella typhimurium | 22 ± 4.3 c | 12 ±1.6 a | 26 ± 0.0 d | 16 ± 0.0 b |
| Escherichia coli | 17 ± 8.5 | 11 ±1.6 | 20 ± 0.0 | 12 ± 0.0 |
| Extended spectrum ß-lactamase producing (ESBL) Escherichia coli | 15 ± 4.3 b | 0 ± 0 | 10 ± 1.6 a | 0 ± 0 |
| Enterobacter cloacae | 18 ± 5.8 c | 11 ± 0 a | 22 ± 0.0 d | 16 ± 0.0 b |
| Multidrug resistant Pseudomonas aeruginosa (MDR) | 16 ± 3.2 b | 14 ± 4.3 a | 22 ± 0.0 c | 14 ± 0.0 a |
| Citrobacter koseri | 16 ± 5.6 b | 12 ± 1.6 a | 24 ± 0.0 d | 18 ± 0.0 c |
| Proteus mirabilis | 14 ± 3.2 a | 0 ± 0 | 30 ± 0.0 b | 0 ± 0 |
| Fungal | Amph B | |||
| Geotricum capitatum | 26 ± 3.2 c | 16 ± 7.1 a | 22 ± 0.0 b | |
| Cándida albicans | 13 ± 1.6 b | 13 ± 1.6 b | 11 ± 0.0 a | |
| Cándida galabrata | 15 ± 2.8 a | 19 ± 3.2 b | 14 ± 0.0 a | |
| Cándida parapsilosis | 27 ± 4.3 b | 26 ± 7.1 b | 18 ± 0.0 a | |
| Cándida tropicalis | 21 ± 9 b | 21 ± 1.6 b | 12 ± 0.0 a | |
| Bacteria (Clinical Isolate) | Ethanolic Extract | |||
|---|---|---|---|---|
| Leaf | Rhizome | |||
| Gram+ | MIC | MBC | MIC | MBC |
| Staphylococcus aureus | 0.78 | 12.5 | - | - |
| Methicillin resistant Staphylococcus aureus (MR) | 0.39 | 6.25 | 3.12 | 50 |
| Staphylococcus epidermidis | 1.56 | 25 | 3.12 | 50 |
| Enterococcus faecalis | 0.39 | 6.25 | 1.56 | 25 |
| Enterococcus faecium | 0.39 | 6.25 | 0.195 | 3.125 |
| Listeria monocytogenes | 0.39 | 6.25 | - | - |
| Gram- | ||||
| Klebsiella pneumonia | 1.56 | 25 | 3.12 | 50 |
| Multidrug resistant (MDR) Klebsiella pneumonia | 0.39 | 6.25 | 3.12 | 50 |
| Salmonella typhimurium | 0.195 | 3.125 | 3.12 | 50 |
| Escherichia coli | 0.39 | 6.25 | 3.12 | 50 |
| Extended spectrum ß-lactamase producing (ESBL) Escherichia coli | 1.56 | 25 | - | - |
| Enterobacter cloacae | 0.39 | 6.25 | 3.12 | 50 |
| Multidrug resistant Pseu-domonas aeruginosa (MDR) | 0.78 | 12.5 | 1.56 | 25 |
| Citrobacter koseri | 0.78 | 12.5 | 3.12 | 50 |
| Proteus mirabilis | 1.56 | 25 | - | - |
| Fungal | MIC | MFC | MIC | MFC |
| Geotricum capitatum | 0.096 | 0.195 | 0.195 | 0.390 |
| Cándida albicans | 0.195 | 0.390 | 0.195 | 0.390 |
| Cándida galabrata | 0.195 | 0.390 | 0.195 | 0.390 |
| Cándida parapsilosis | 0.096 | 0.195 | 0.096 | 0.195 |
| Cándida tropicalis | 0.195 | 0.390 | 0.195 | 0.390 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdi, A.; Jaramillo-Carmona, S.; Rodríguez-Arcos, R.; Jiménez-Araujo, A.; Lachaal, M.; Karray-Bouraoui, N.; Guillén-Bejarano, R. Phytochemical Characterization and Bioactivity of Asparagus acutifolius: A Focus on Antioxidant, Cytotoxic, Lipase Inhibitory and Antimicrobial Activities. Molecules 2021, 26, 3328. https://doi.org/10.3390/molecules26113328
Hamdi A, Jaramillo-Carmona S, Rodríguez-Arcos R, Jiménez-Araujo A, Lachaal M, Karray-Bouraoui N, Guillén-Bejarano R. Phytochemical Characterization and Bioactivity of Asparagus acutifolius: A Focus on Antioxidant, Cytotoxic, Lipase Inhibitory and Antimicrobial Activities. Molecules. 2021; 26(11):3328. https://doi.org/10.3390/molecules26113328
Chicago/Turabian StyleHamdi, Amel, Sara Jaramillo-Carmona, Rocío Rodríguez-Arcos, Ana Jiménez-Araujo, Mokhtar Lachaal, Najoua Karray-Bouraoui, and Rafael Guillén-Bejarano. 2021. "Phytochemical Characterization and Bioactivity of Asparagus acutifolius: A Focus on Antioxidant, Cytotoxic, Lipase Inhibitory and Antimicrobial Activities" Molecules 26, no. 11: 3328. https://doi.org/10.3390/molecules26113328
APA StyleHamdi, A., Jaramillo-Carmona, S., Rodríguez-Arcos, R., Jiménez-Araujo, A., Lachaal, M., Karray-Bouraoui, N., & Guillén-Bejarano, R. (2021). Phytochemical Characterization and Bioactivity of Asparagus acutifolius: A Focus on Antioxidant, Cytotoxic, Lipase Inhibitory and Antimicrobial Activities. Molecules, 26(11), 3328. https://doi.org/10.3390/molecules26113328