Early Steps in the Biosynthetic Pathway of Rishirilide B

 , ,
, ,
Abstract

1. Introduction
2. Results
2.1. Overexpression of The Regulatory Genes rslR1, rslR2 and rslR3 Led to An Increased Rishirilide Production
2.2. Rishirilide Production Is Not Strongly Affected by Deletions of rslO2, rslO6, and rslH.
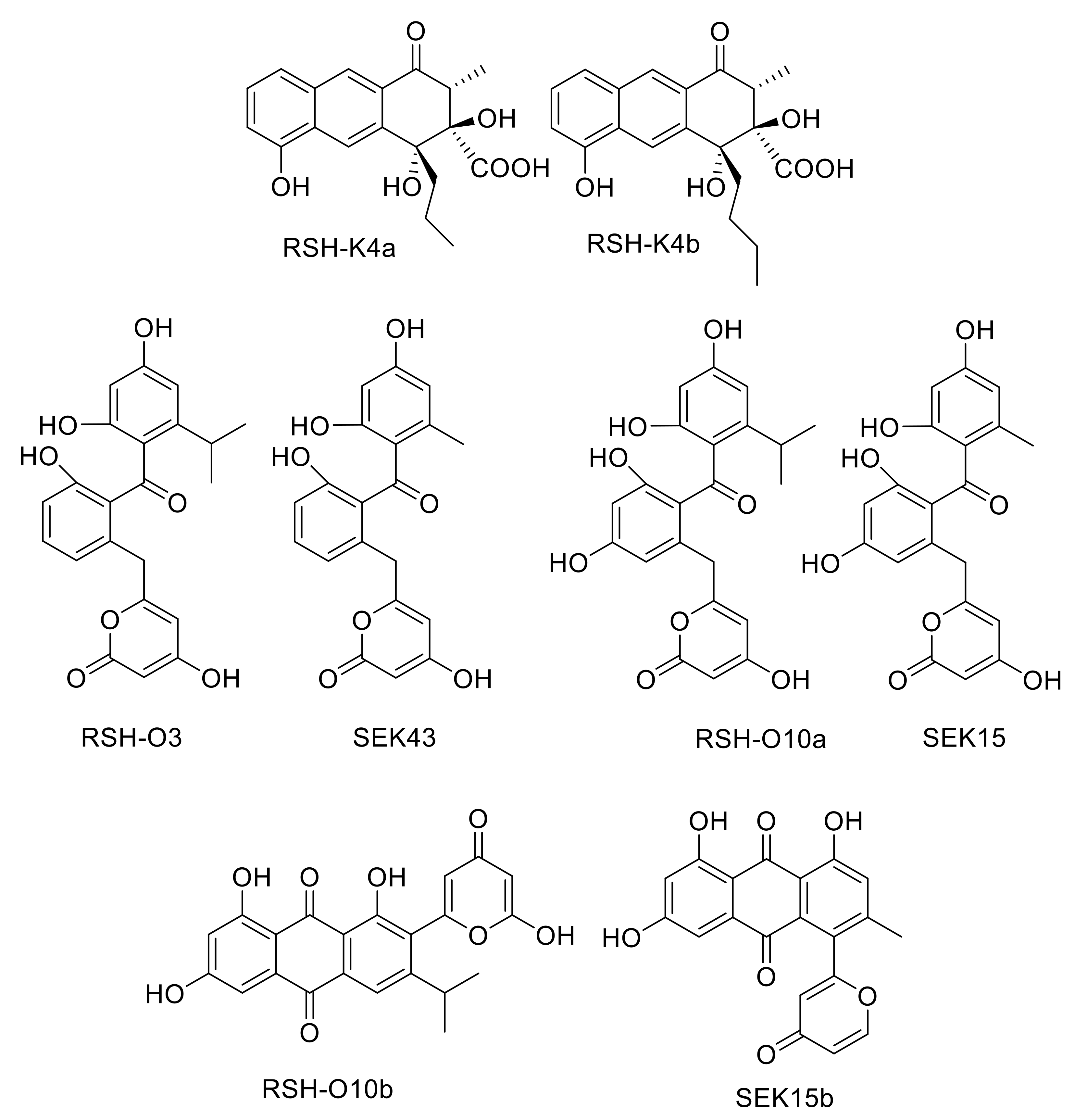
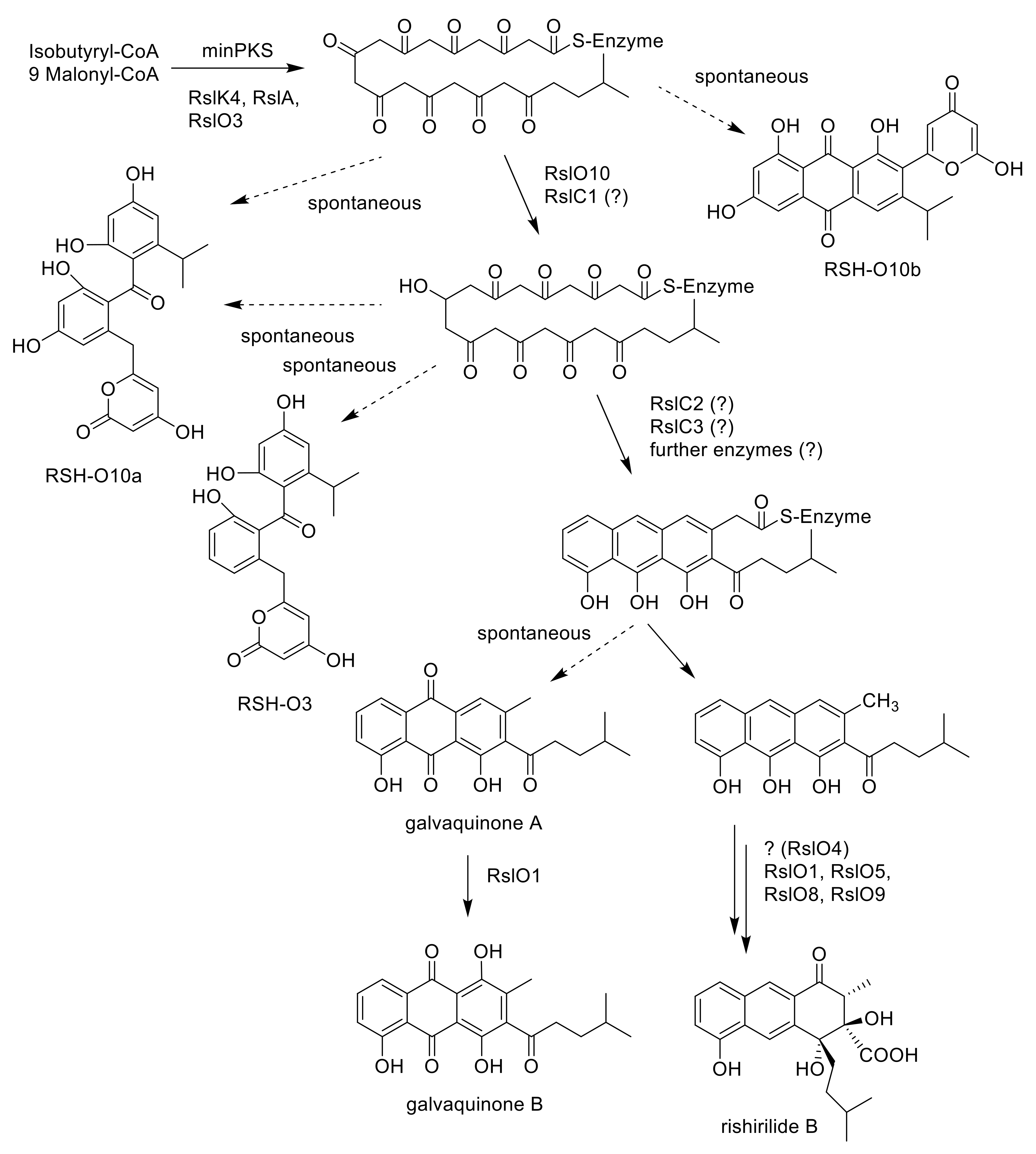
2.3. Deletion of rslK4 Leads to the Formation of Derivatives with Alterations in the Side Chain, Indicating That RslK4 Is Involved in Selecting the Starter Unit for the PKS
2.4. Deletion of rslO3 Leads to the Formation of A Shunt Product, Which Is Similar to SEK43, Indicating That RslO3 Is Involved in the Formation of the Starter Unit for the PKS
2.5. Deletion of rslO10 Revealed the Function of RslO10 as Early Acting Ketoreductase
2.6. Expression of Gene Cassettes in S. albus J1074
2.7. Galvaquinone A Is Produced by Mutants Lacking RslO1
2.8. Galvaquinone A and B Are Produced by Mutants Lacking rslO4
3. Discussion
4. Materials and Methods
4.1. General
4.2. Strains, Plasmids and Culture Conditions
4.3. Generation of Gene Deletion Mutants
4.4. Complementation of S. albus::cos4∆H, S. albus::cos4∆K4, S. albus::cos4∆O1, S. albus::cos4∆O2, S. albus::cos4∆O3, S. albus::cos4∆O4, S. albus::cos4∆O6 and S. albus::cos4∆O10.
4.5. Generation of Strains Containing Different Gene Cassettes
4.5.1. S. albus x pUWL-H-R1R2R3
4.5.2. S. albus J1074 x pUWLR1R2R3 x construct 1
4.5.3. S. albus J1074 x pUWLR1R2R3 x construct 2
4.5.4. S. albus J1074 x pUWLR1R2R3 x construct 3, S. albus J1074 x pUWLR1R2R3 x construct 4, S. albus J1074 x pUWLR1R2R3 x construct 5 and S. albus J1074 pUWLR1R2R3 x construct 6
4.6. Metabolite Analysis by HPLC-MS.
4.7. Production of RSH-K4a, RSH-O10a, RSH-O10b, RSH-O3 and Galvaquinones A and B
4.8. Isolation and Purification of RSH-K4a, RSH-O10a, RSH-O10b, RSH-O3 and Galvaquinone A and Galvaquinone B
4.9. NMR Analysis of Isolated Compounds
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. Engl. 2009, 48, 4688–4716. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, H.; Nakayama, Y.; Takahashi, M.; Uetsuki, S.; Kido, M.; Fukuyama, Y. Structures of rishirilides A and B, alpha 2-macroglobulin inhibitors produced by Streptomyces rishiriensis OFR-1056. J. Antibiot. Tokyo 1984, 37, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sun, C.; Huang, H.; Gui, C.; Wang, L.; Li, Q.; Ju, J. Biosynthetic Baeyer-Villiger Chemistry Enables Access to Two Anthracene Scaffolds from a Single Gene Cluster in Deep-Sea-Derived Streptomyces olivaceus SCSIO T05. J. Nat. Prod. 2018, 81, 1570–1577. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Probst, K.; Linnenbrink, A.; Arnold, M.; Paululat, T.; Zeeck, A.; Bechthold, A. Cloning and heterologous expression of three type II PKS gene clusters from Streptomyces bottropensis. ChemBioChem 2012, 13, 224–230. [Google Scholar] [CrossRef]
- Schwarzer, P.; Wunsch-Palasis, J.; Bechthold, A.; Paululat, T. Biosynthesis of Rishirilide B. Antibiotics 2018, 7, 20. [Google Scholar] [CrossRef]
- Tsypik, O.; Makitrynskyy, R.; Frensch, B.; Zechel, D.L.; Paululat, T.; Teufel, R.; Bechthold, A. Oxidative carbon backbone rearrangement in rishirilide biosynthesis. J. Am. Chem. Soc. 2020, 142, 5913–5917. [Google Scholar] [CrossRef]
- Teufel, R. Flavin-catalyzed redox tailoring reactions in natural product biosynthesis. Arch. Biochem. Biophys. 2017, 632, 20–27. [Google Scholar] [CrossRef]
- Chenprakhon, P.; Wongnate, T.; Chaiyen, P. Monooxygenation of aromatic compounds by flavin-dependent monooxygenases. Protein Sci. 2019, 28, 8–29. [Google Scholar] [CrossRef]
- Elyas, Y.Y.A.; Miyatani, K.; Bito, T.; Uraji, M.; Hatanaka, T.; Shimizu, K.; Arima, J. Active site pocket of Streptomyces d-stereospecific amidohydrolase has functional roles in aminolysis activity. J. Biosci. Bioeng. 2018, 126, 293–300. [Google Scholar] [CrossRef]
- Moore, B.S.; Hertweck, C. Biosynthesis and attachment of novel bacterial polyketide synthase starter units. Nat. Prod. Rep. 2002, 19, 70–99. [Google Scholar]
- Marti, T.; Hu, Z.; Pohl, N.L.; Shah, A.N.; Khosla, C. Cloning, nucleotide sequence, and heterologous expression of the biosynthetic gene cluster for R1128, a non-steroidal estrogen receptor antagonist. Insights into an unusual priming mechanism. J. Biol. Chem. 2000, 275, 33443–33448. [Google Scholar] [CrossRef] [PubMed]
- Harunari, E.; Komaki, H.; Igarashi, Y. Biosynthetic origin of butyrolactol A, an antifungal polyketide produced by a marine-derived Streptomyces. Beilstein J. Org. Chem. 2017, 13, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Räty, K.; Kantola, J.; Hautala, A.; Hakala, J.; Ylihonko, K.; Mäntsälä, P. Cloning and characterization of Streptomyces galilaeus aclacinomycins polyketide synthase (PKS) cluster. Gene 2002, 293, 115–122. [Google Scholar] [CrossRef]
- Metsä-Ketelä, M.; Palmu, K.; Kunnari, T.; Ylihonko, K.; Mäntsälä, P. Engineering anthracycline biosynthesis toward angucyclines. Antimicrob. Agents Chemother. 2003, 47, 1291–1296. [Google Scholar] [CrossRef][Green Version]
- Grimm, A.; Madduri, K.; Ali, A.; Hutchinson, C.R. Characterization of the Streptomyces peucetius ATCC 29050 genes encoding doxorubicin polyketide synthase. Gene 1994, 151, 1–10. [Google Scholar] [CrossRef]
- Tang, Y.; Lee, T.S.; Khosla, C. Engineered biosynthesis of regioselectively modified aromatic polyketides using bimodular polyketide synthases. PLoS Biol. 2004, 2, 0227–0238. [Google Scholar] [CrossRef]
- Pan, H.; Tsai, S.; Meadows, E.S.; Miercke, L.J.; Keatinge-Clay, A.T.; O’Connell, J.; Khosla, C.; Stroud, R.M. Crystal structure of the priming beta-ketosynthase from the R1128 polyketide biosynthetic pathway. Structure 2002, 10, 1559–1568. [Google Scholar] [CrossRef]
- McDaniel, R.; Ebert-Khosla, S.; Hopwood, D.A.; Khosla, C. Rational design of aromatic polyketide natural products by recombinant assembly of enzymatic subunits. Nature 1995, 375, 549–554. [Google Scholar] [CrossRef]
- Patel, M.P.; Liu, W.S.; West, J.; Tew, D.; Meek, T.D.; Thrall, S.H. Kinetic and chemical mechanisms of the fabG-encoded Streptococcus pneumoniae beta-ketoacyl-ACP reductase. Biochemistry 2005, 44, 16753–16765. [Google Scholar] [CrossRef]
- Pickens, L.B.; Kim, W.; Wang, P.; Zhou, H.; Watanabe, K.; Gomi, S.; Tang, Y.J. Biochemical analysis of the biosynthetic pathway of an anticancer tetracycline SF2575. J. Am. Chem. Soc. 2009, 131, 17677–17689. [Google Scholar] [CrossRef]
- Ames, B.D.; Lee, M.Y.; Moody, C.; Zhang, W.; Tang, Y.; Tsai, S.C. Structural and biochemical characterization of ZhuI aromatase/cyclase from the R1128 polyketide pathway. Biochemistry 2011, 50, 8392–8406. [Google Scholar] [CrossRef] [PubMed]
- Hertweck, C.; Luzhetskyy, A.; Rebets, Y.; Bechthold, A. Type II polyketide synthases: Gaining a deeper insight into enzymatic teamwork. Nat. Prod. Rep. 2007, 24, 162–190. [Google Scholar] [CrossRef] [PubMed]
- Maier, S.; Heitzler, T.; Asmus, K.; Brötz, E.; Hardter, U.; Hesselbach, K.; Paululat, T.; Bechthold, A. Functional characterization of different ORFs including luciferase-like monooxygenase genes from the mensacarcin gene cluster. Chembiochem. 2015, 16, 1175–1182. [Google Scholar] [CrossRef]
- Grocholski, T.; Koskiniemi, H.; Lindqvist, Y.; Mäntsälä, P.; Niemi, J.; Schneider, G. Crystal structure of the cofactor-independent monooxygenase SnoaB from Streptomyces nogalater: Implications for the reaction mechanism. Biochemistry 2010, 49, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Sciara, G.; Kendrew, S.G.; Miele, A.E.; Marsh, N.G.; Federici, L.; Malatesta, F.; Schimperna, G.; Savino, C.; Vallone, B. The structure of ActVA-Orf6, a novel type of monooxygenase involved in actinorhodin biosynthesis. EMBO J. 2003, 22, 205–215. [Google Scholar] [CrossRef]
- Summers, R.G.; Wendt-Pienkowski, E.; Motamedi, H.; Hutchinson, C.R. The tcmVI region of the tetracenomycin C biosynthetic gene cluster of Streptomyces glaucescens encodes the tetracenomycin F1 monooxygenase, tetracenomycin F2 cyclase, and, most likely, a second cyclase. J. Bacteriol. 1993, 175, 7571–7580. [Google Scholar] [CrossRef][Green Version]
- Li, W.; Ju, J.; Rajski, S.R.; Osada, H.; Shen, B. Characterization of the tautomycin biosynthetic gene cluster from Streptomyces spiroverticillatus unveiling new insights into dialkylmaleic anhydride and polyketide biosynthesis. J. Biol. Chem. 2008, 283, 28607–18617. [Google Scholar] [CrossRef]
- Zhang, Y.; Muyrers, J.P.; Rientjes, J.; Stewart, A.F. Phage annealing proteins promote oligonucleotide-directed mutagenesis in Escherichia coli and mouse ES cells. BMC Mol. Biol. 2003, 4, 1–14. [Google Scholar] [CrossRef]
- Doumith, M.; Weingarten, P.; Wehmeier, U.F.; Salah-Bey, K.; Benhamou, B.; Capdevila, C.; Michel, J.M.; Piepersberg, W.; Raynal, M.C. Analysis of genes involved in 6-deoxyhexose biosynthesis and transfer in Saccharopolyspora erythraea. Mol. Gen. Genet. 2000, 264, 477–485. [Google Scholar] [CrossRef]
- Herrmann, S.; Siegl, T.; Luzhetska, M.; Petzke, L.; Jilg, C.; Welle, E.; Erb, A.; Leadlay, P.F.; Bechthold, A.; Luzhetskyy, A. Site-specific recombination strategies for engineering actinomycete genomes. Appl. Environ. Microbiol. 2012, 78, 1804–1812. [Google Scholar] [CrossRef]
Sample Availability: All samples of the compounds are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description of Mutant Strains | Compound Produced by the Strain |
|---|---|
| S. albus::cos4 x pUWLR1R2R3 | Rishirilide B |
| S. albus::cos4∆rslO2 x pUWLR1R2R3 | Rishirilide B |
| S. albus::cos4∆rslO6 x pUWLR1R2R3 | Rishirilide B |
| S. albus::cos4∆rslH x pUWLR1R2R3 | Rishirilide B |
| S. albus::cos4∆rslK4 x pUWLR1R2R3 | RSH-K4a, RSH-K4b and rishirilide B |
| S. albus::cos4∆rslO3 x pUWLR1R2R3 | RSH-O3 |
| S. albus::cos4∆rslO10 x pUWLR1R2R3 | RSH-O10a and RSH-O10b |
| S. albus J1074 x pUWLR1R2R3 x construct 1 (construct 1: rslK1, rslK2, rslK3, rslK4, rslA) | RSH-O10a and RSH-O10b |
| S. albus J1074 x pUWLR1R2R3 x construct 2 (construct 2: rslK1, rslK2, rslK3, rslK4, rslA, rslC1, rslC2, rslC3) | RSH-O10a and RSH-O10b |
| S. albus J1074 x pUWLR1R2R3 x construct 3 (construct 3: rslK1, rslK2, rslK3, rslK4, rslA, rslO10) | RSH-O10a and RSH-O3 |
| S. albus J1074 x pUWLR1R2R3 x construct 4 (construct 4: rslK1, rslK2, rslK3, rslK4, rslA, rslO10, rslC1, rslC2, rslC3) | RSH-O10a and RSH-O3 |
| S. albus J1074 x pUWLR1R2R3 x construct 5 (construct 5: rslK1, rslK2, rslK3, rslK4, rslA, rslO10, rslO3) | RSH-O10a |
| S. albus J1074 x pUWLR1R2R3 x construct 6 (construct 6: rslK1, rslK2, rslK3, rslK4, rslA, rslO10, rslO3, rslC1, rslC2, rslC3) | RSH-O10a |
| S. albus::cos4∆rslO1 x pUWLR1R2R3 | Galvaquinone A |
| S. albus::cos4∆rslO4 x pUWLR1R2R3 | Galvaquinone A and B |
| Analyzed Genes in This Study | Deduced Function |
|---|---|
| rslK4 | ketosynthetase, selection of unusual starter unit |
| rslO1 | luciferase-like monooxygenase, involved in late stage rishirilide biosynthesis, but before rearrangement by rslO9 |
| rslO2 | Flavin reductase |
| rslO3 | 3-oxoacyl ACP reductase, catalysis of l-valine to isohexanoyl-ACP conversion for formation of the starter unit |
| rslO4 | antibiotic biosynthesis monooxygenase, involved in late stage rishirilide biosynthesis, but before rearrangement by rslO9 |
| rslO6 | Flavin dependent oxidoreductase, not essential for rishirilide biosynthesis |
| rslO10 | C9-ketoreductase, support of C7–C12 ring cyclisation |
| rslH | amidohydrolase, not essential for rishirilide biosynthesis |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarzer, P.; Tsypik, O.; Zuo, C.; Alali, A.; Wunsch-Palasis, J.; Heitzler, T.; Derochefort, J.; Bernhardt, M.; Yan, X.; Paululat, T.; et al. Early Steps in the Biosynthetic Pathway of Rishirilide B. Molecules 2020, 25, 1955. https://doi.org/10.3390/molecules25081955
Schwarzer P, Tsypik O, Zuo C, Alali A, Wunsch-Palasis J, Heitzler T, Derochefort J, Bernhardt M, Yan X, Paululat T, et al. Early Steps in the Biosynthetic Pathway of Rishirilide B. Molecules. 2020; 25(8):1955. https://doi.org/10.3390/molecules25081955
Chicago/Turabian StyleSchwarzer, Philipp, Olga Tsypik, Chijian Zuo, Ahmad Alali, Julia Wunsch-Palasis, Tanja Heitzler, Jana Derochefort, Mirjam Bernhardt, Xiaohui Yan, Thomas Paululat, and et al. 2020. "Early Steps in the Biosynthetic Pathway of Rishirilide B" Molecules 25, no. 8: 1955. https://doi.org/10.3390/molecules25081955
APA StyleSchwarzer, P., Tsypik, O., Zuo, C., Alali, A., Wunsch-Palasis, J., Heitzler, T., Derochefort, J., Bernhardt, M., Yan, X., Paululat, T., & Bechthold, A. (2020). Early Steps in the Biosynthetic Pathway of Rishirilide B. Molecules, 25(8), 1955. https://doi.org/10.3390/molecules25081955