
Multitarget Approach to Drug Candidates against Alzheimer’s Disease Related to AChE, SERT, BACE1 and GSK3β Protein Targets

Abstract
:1. Introduction
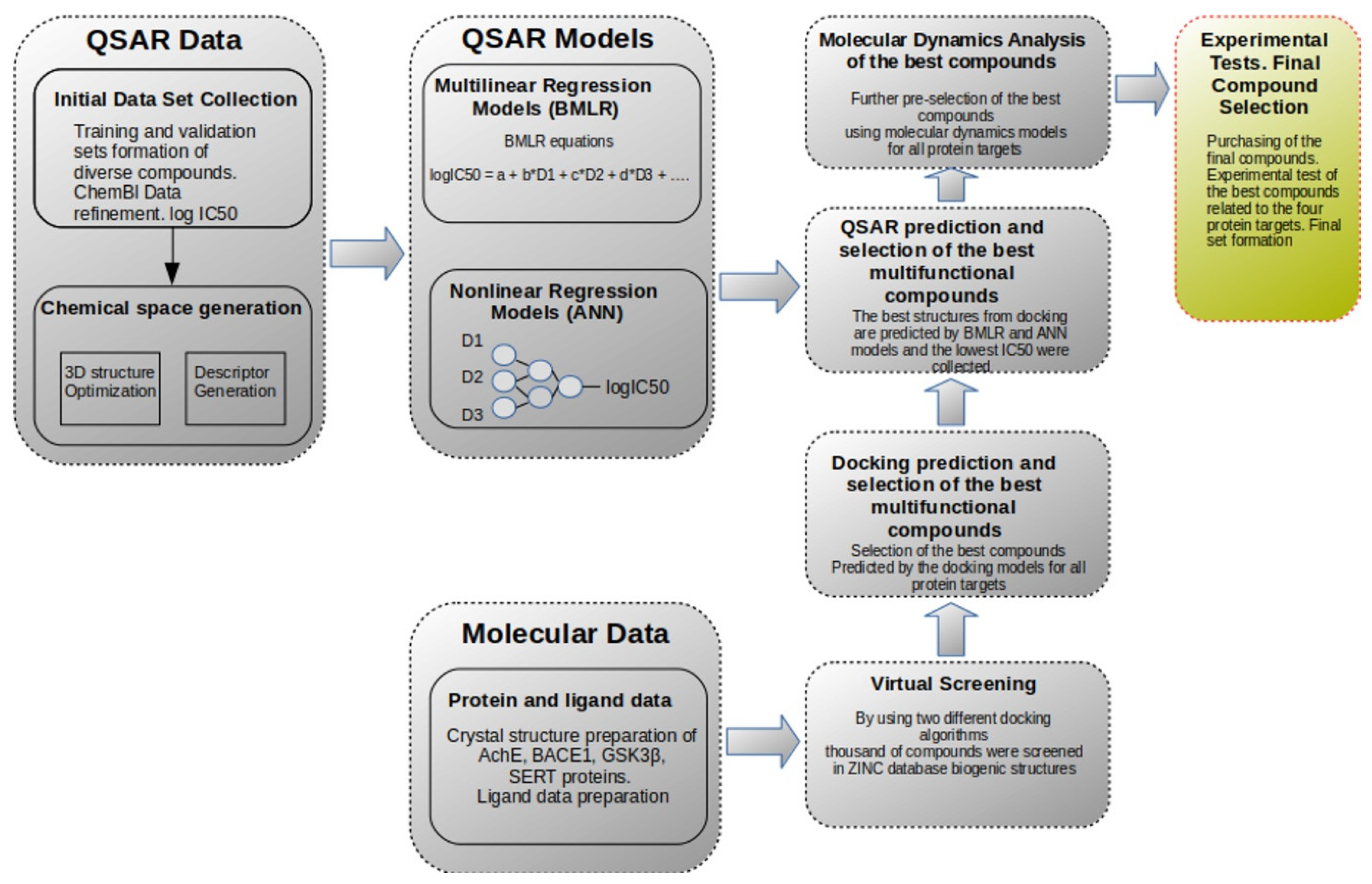
2. Results and Discussion
2.1. Computational Studies
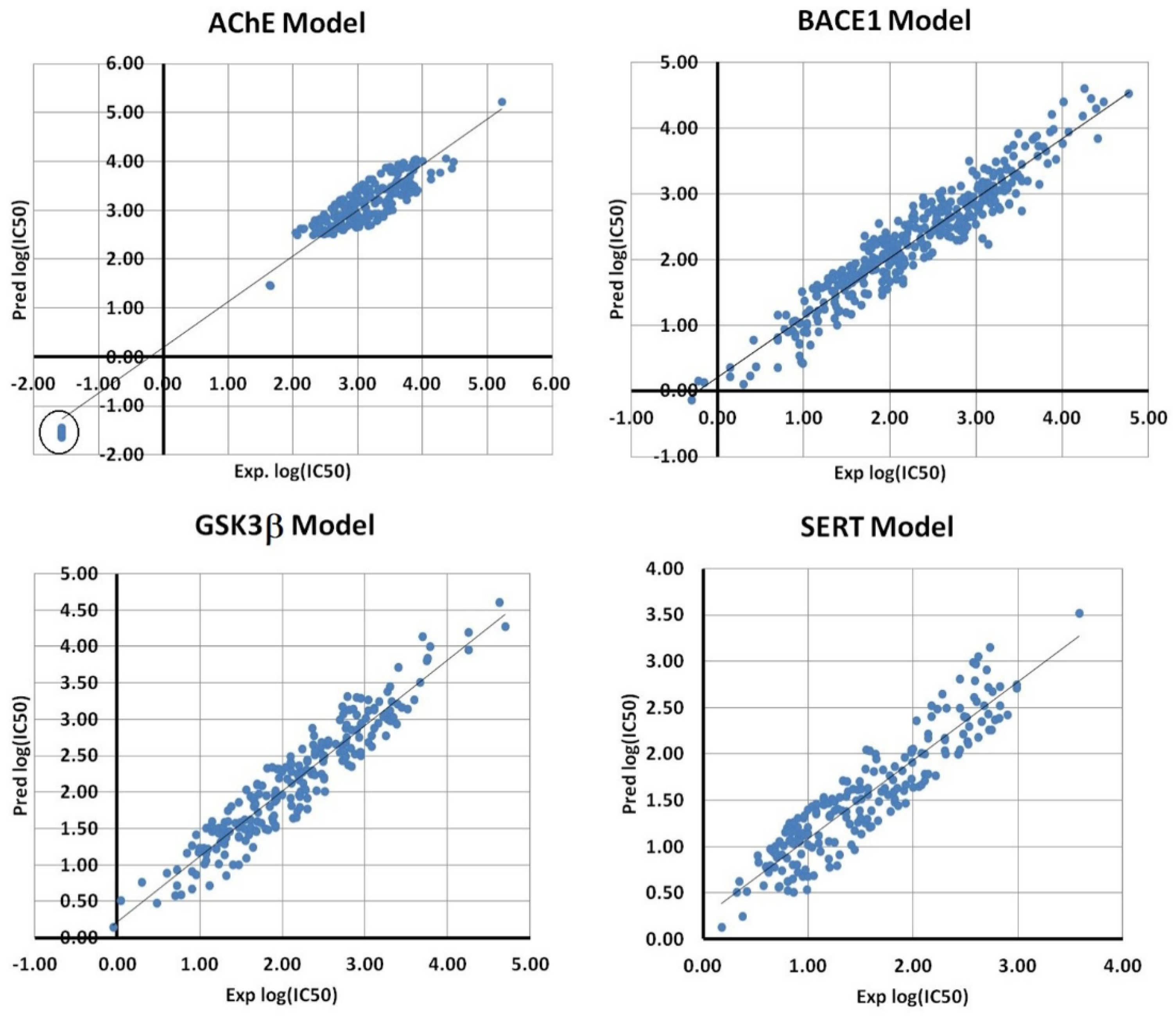
2.2. BMLR Models
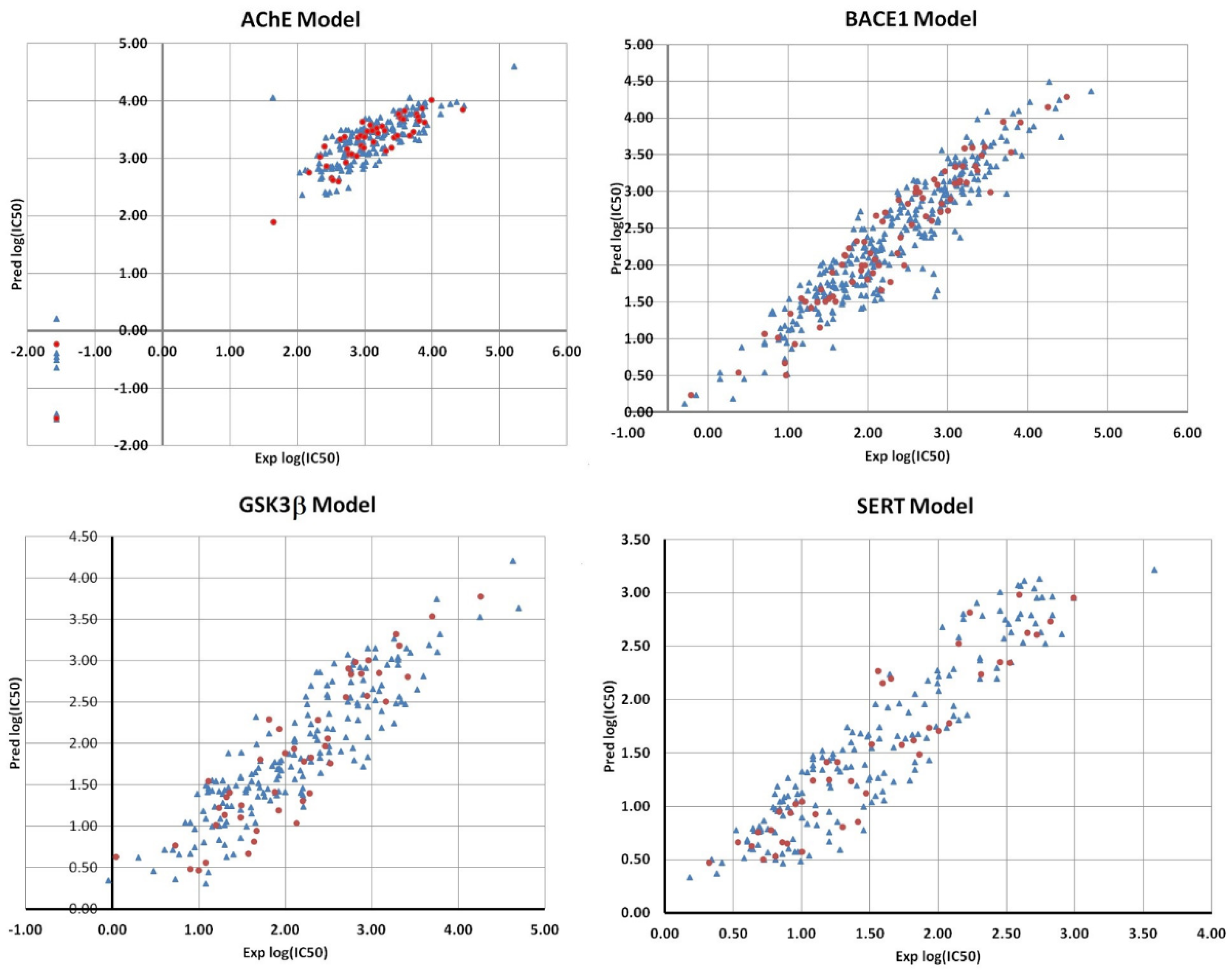
2.3. ANN Models
2.4. Virtual Screening with Glide VSW Module of the Schrödinger Suite
- -
- AChE: 526 compounds; ∆G = −18.71…−10.97 kcal/mol; LE = 0.32…0.71
- -
- BACE1: 457 compounds; ∆G = −12.69…−6.72 kcal/mol; LE = 0.19…0.48;
- -
- GSK3β: 506 compounds; ∆G = −20.39…−12.27 kcal/mol; LE = 0.31…0.79;
- -
- SERT: 526 compounds; ∆G = −15.47…−8.99 kcal/mol; LE = 0.24…0.53.
2.5. Virtual Screening with Autodock Vina 1.1.2
- -
- AChE: 832 compounds; ∆G = −13.8…−8.1 kcal/mol; LE = 0.35…0.66;
- -
- BACE1: 1633 compounds; ∆G = −12.9…−8.0 kcal/mol; LE = 0.35…0.53;
- -
- GSK3β: 470 compounds; ∆G = −10.3…−8.0 kcal/mol; LE = 0.35…0.46;
- -
- SERT: 1692 compounds; ∆G = −12.5…−8.0 kcal/mol; LE = 0.35…0.51.
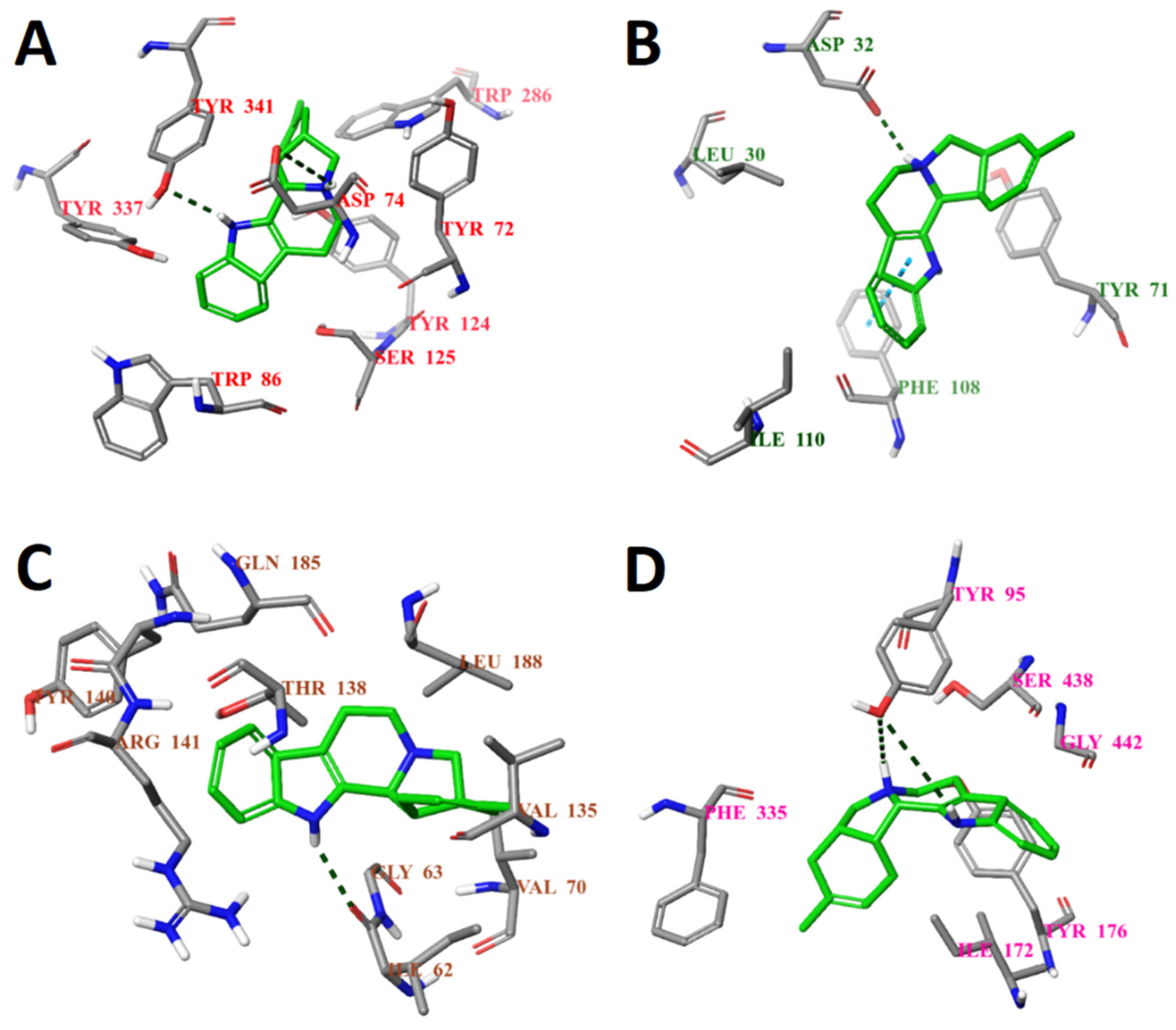
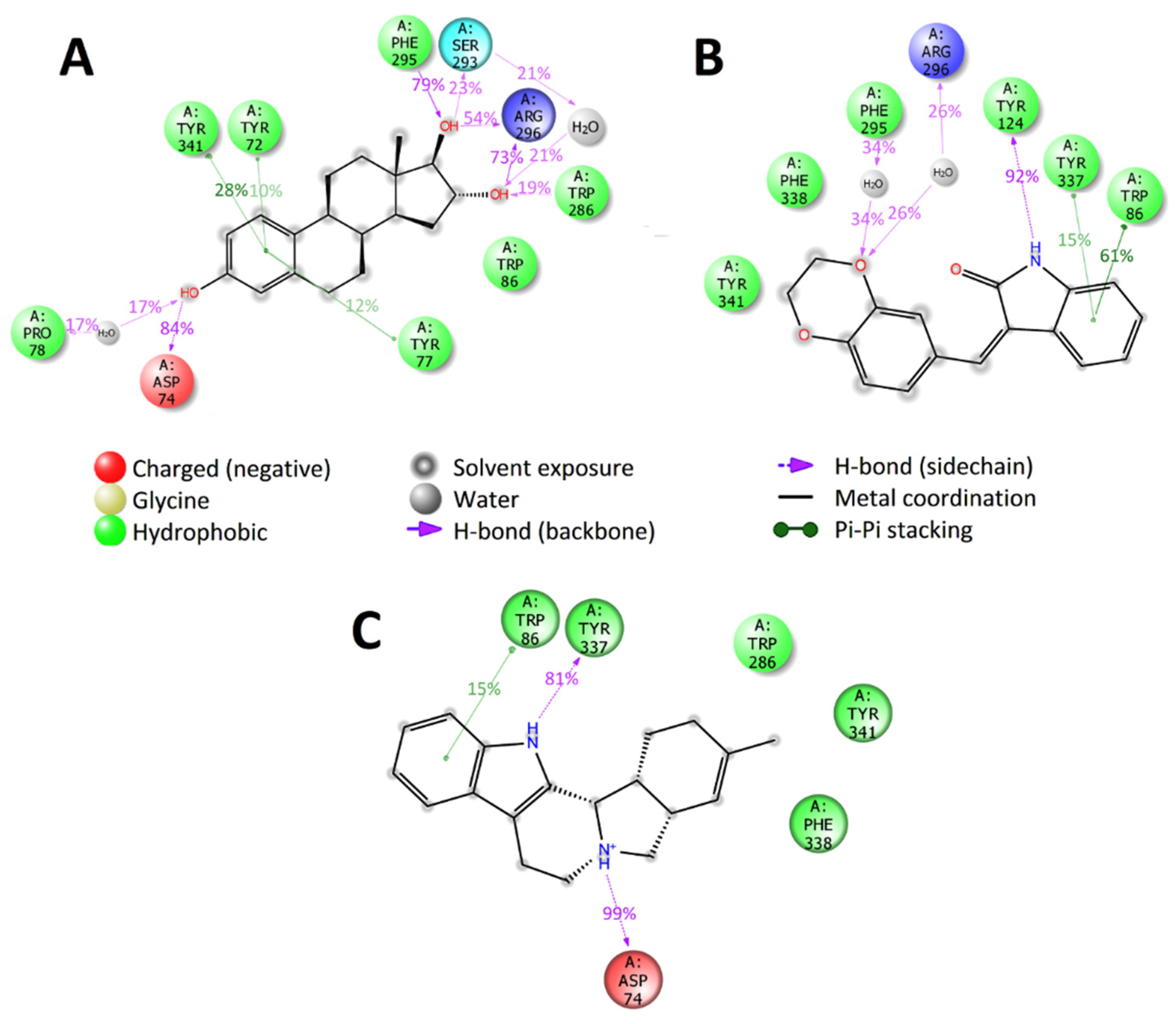
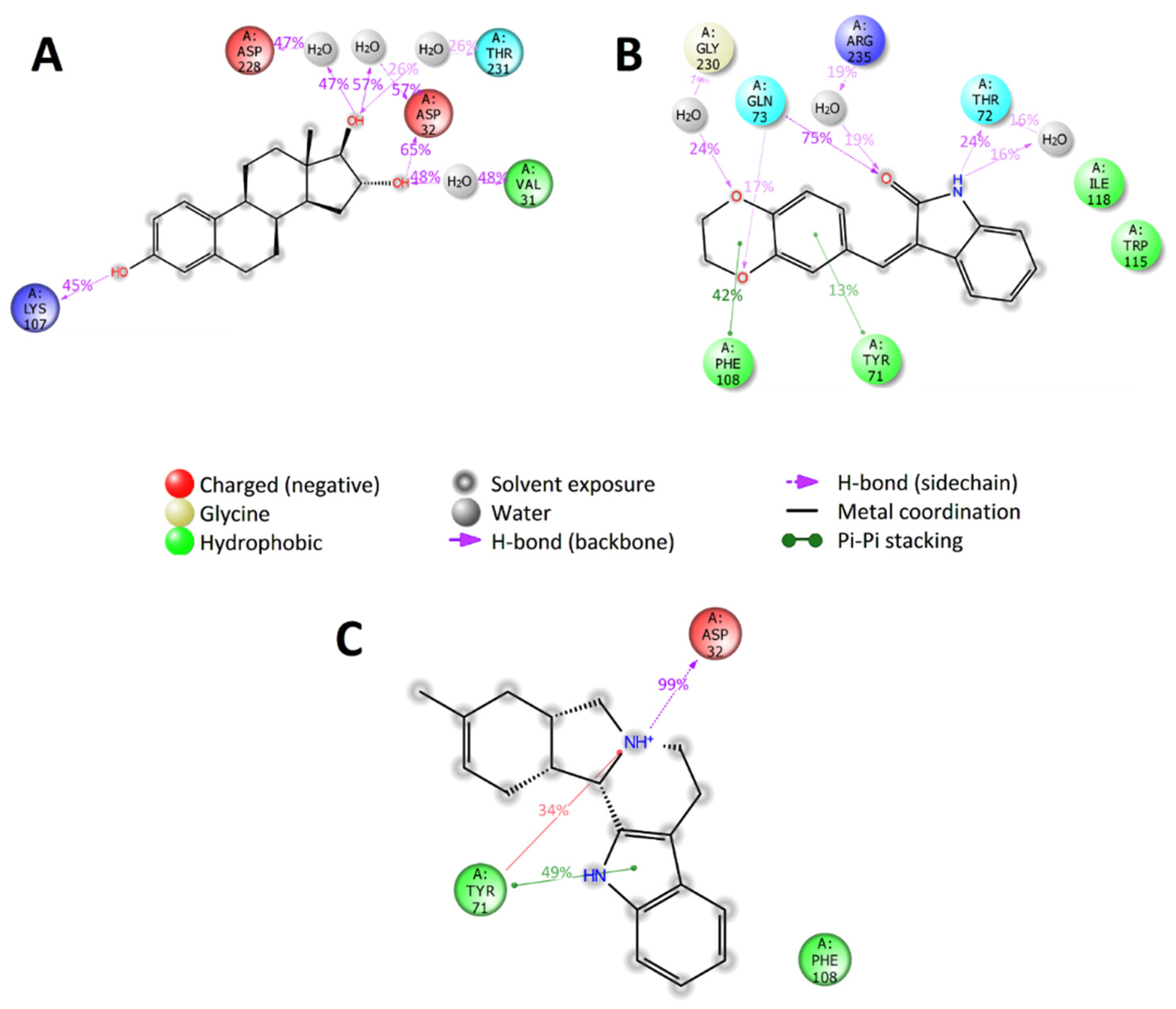
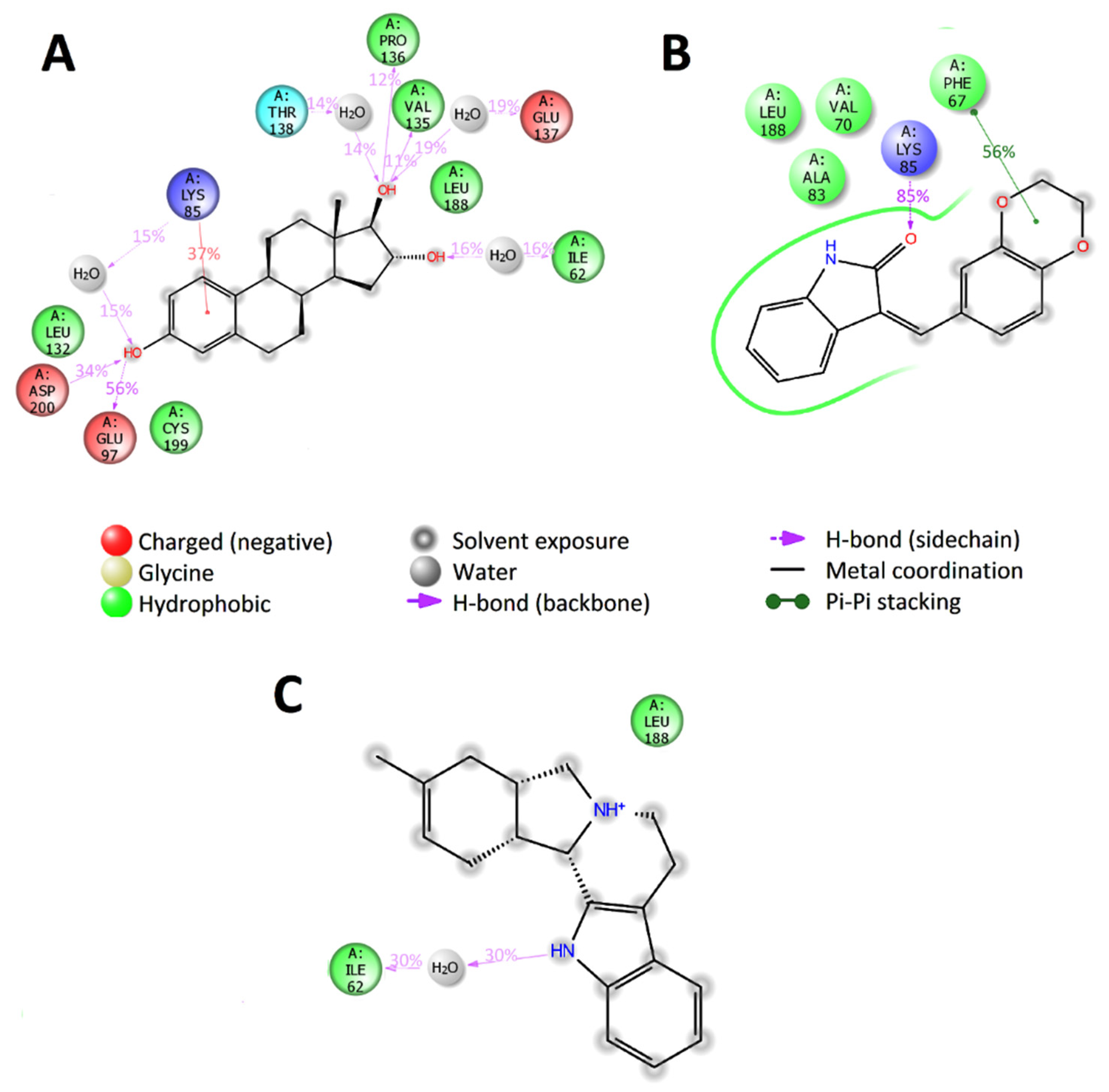
2.6. Selection of Active Compounds
2.7. Enzymatic Assay Results
3. Materials and Methods
3.1. Data and Compound Libraries
3.1.1. QSAR Modeling Datasets
3.1.2. Protein Structures
3.2. Preparation of Protein Target Structures and Compounds Library
3.3. High-Throughput Virtual Molecular Docking Screening (HTVS)
3.4. Molecular Dynamics Simulations
3.5. Structure Optimizations and Molecular Descriptor Generation of the QSAR Datasets
3.6. Multilinear Models Based on the BMLR Method
3.7. Nonlinear Models Based on ANN
- (1)
- Training and validation sets: the respective data sets in Section 3.1.1 were divided into training and validation sets and normalized within (−1,1) range. The validation set consists of each 5th data point selected by using the log IC50 values ordered in ascending way. This selected set is employed to evaluate the root mean squared error (RMSval) or the squared Pearson’s correlation coefficient R2val during the training procedure. The RMSval was used as a stopping criterion for the training algorithm, once it started to increase above a certain value.
- (2)
- Input variable selection: a smaller pool of statistically significant descriptors was generated from the total pool of descriptors obtained in Section 3.5 by the following manner: (1) the first 10 to 20 descriptors that correlated best with the given activity together with (2) the descriptors obtained from the BMLR models in Section 3.6.
- (3)
- Network topology: in order to follow the generality principle for predictability of an ANN model [68], we limited ourselves to architecture with not more than two hidden layers of the nets. By doing this, we also tried to keep the total number of weights as low as possible in order to avoid overparameterizing the network. Thus, networks with the following architectures were considered n-h1-h2-1 or n-h1-1, where n is the number of input descriptors, h1 is the number of neurons in the first hidden layer, h2 is the number of neurons in the second hidden layer and one is the single output neuron in the output layer corresponding to log IC50.
- (4)
- ANN training and best model development (BeANN): we used the following ANN parameters for all models prior the sequential training procedure: learning rate η = 0.1 or 0.2, momentum α = 0.02 and number of training epochs (stopping criterion) not more than 700. For all nets the hidden and output neurons used tanh activation function confined within (−1,1). The initial set of the weights comprised of values between (−1,1) with the closest to zero total mean chosen among 20 random trials. The reason for this is the selection of “good” initial weights that would lead to faster convergence during training procedure.
3.8. Virtual Screening Based on the QSAR Models
3.9. Experimental Enzymatic Assays
3.9.1. Compounds
3.9.2. Enzymes Inhibition Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Patterson, C. World Alzheimer Report 2018. The State of the Art of Dementia Research: New Frontiers; Alzheimer’s Disease International (ADI): London, UK, 2018; pp. 32–36. [Google Scholar]
- Cornutiu, G. The epidemiological scale of Alzheimer’s disease. J. Clin. Med. Res. 2015, 7, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Srivastava, P.; Seth, A.; Tripathi, P.N.; Banerjee, A.G.; Shrivastava, S.K. Comprehensive review of mechanisms of pathogenesis involved in Alzheimer’s disease and potential therapeutic strategies. Prog. Neurobiol. 2019, 174, 53–89. [Google Scholar] [CrossRef]
- Wong, D.F.; Rosenberg, P.B.; Zhou, Y.; Kumar, A.; Raymont, V.; Ravert, H.T.; Dannals, R.F.; Nandi, A.; Brašić, J.R.; Ye, W. In vivo imaging of amyloid deposition in Alzheimer disease using the radioligand 18F-AV-45 (flobetapir F 18). J. Nucl. Med. 2010, 51, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Olabarria, M.; Noristani, H.N.; Verkhratsky, A.; Rodríguez, J.J. Concomitant astroglial atrophy and astrogliosis in a triple transgenic animal model of Alzheimer’s disease. Glia 2010, 58, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Shepardson, N.; Yang, T.; Chen, G.; Walsh, D.; Selkoe, D.J. Soluble amyloid β-protein dimers isolated from Alzheimer cortex directly induce Tau hyperphosphorylation and neuritic degeneration. Proc. Natl. Acad. Sci. USA 2011, 108, 5819–5824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heredia, L.; Helguera, P.; de Olmos, S.; Kedikian, G.; Vigo, F.S.; LaFerla, F.; Staufenbiel, M.; de Olmos, J.; Busciglio, J.; Cáceres, A. Phosphorylation of actin-depolymerizing factor/cofilin by LIM-kinase mediates amyloid β-induced degeneration: A potential mechanism of neuronal dystrophy in Alzheimer’s disease. J. Neurosci. 2006, 26, 6533–6542. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D. Evidence of oxidative stress in Alzheimer’s disease brain and antioxidant therapy. Ann. N. Y. Acad. Sci. 2008, 1147, 70–78. [Google Scholar] [CrossRef]
- Kumar, A.; Nisha, C.M.; Silakari, C.; Sharma, I.; Anusha, K.; Gupta, N.; Nair, P.; Tripathi, T.; Kumar, A. Current and novel therapeutic molecules and targets in Alzheimer’s disease. J. Formos. Med. Assoc. 2016, 115, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Eftekharzadeh, B.; Daigle, J.G.; Kapinos, L.E.; Coyne, A.; Schiantarelli, J.; Carlomagno, Y.; Cook, C.; Miller, S.J.; Dujardin, S.; Amaral, A.S.; et al. Tau protein disrupts nucleocytoplasmic transport in Alzheimer’s disease. Neuron 2018, 99, 925–940. [Google Scholar] [CrossRef] [Green Version]
- Readhead, B.; Haure-Mirande, J.V.; Funk, C.C.; Richards, M.A.; Shannon, P.; Haroutunian, V.; Sano, M.; Liang, W.S.; Beckmann, N.D.; Price, N.D.; et al. Multiscale analysis of independent Alzheimer’s cohorts finds disruption of molecular, genetic, and clinical networks by human Herpesvirus. Neuron 2018, 99, 64–82. [Google Scholar] [CrossRef] [Green Version]
- Dominy, S.S.; Lynch, C.; Ermini, F.; Benedyk, M.; Marczyk, A.; Konradi, A.; Nguyen, M.; Haditsch, U.; Raha, D.; Griffin, C.; et al. Porphyromonas gingivalis in Alzheimer’s disease brains: Evidence for disease causation and treatment with small-molecule inhibitors. Sci. Adv. 2019, 5, eaau33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korolev, I.O. Alzheimer’s disease: A clinical and basic science review. Med. Stud. Res. J. 2014, 4, 24–33. [Google Scholar]
- Ibrahim, M.M.; Gabr, M.T. Multitarget therapeutic strategies for Alzheimer’s disease. Neural Regen. Res. 2019, 14, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Tripathi, A.; Tripathi, P.N.; Prajapati, S.K.; Seth, A.; Tripathi, M.K.; Srivastava, P.; Tiwari, V.; Krishnamurthy, S.; Shrivastava, S.K. Design and development of multitarget-directed N-Benzylpiperidine analogs as potential candidates for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 167, 510–524. [Google Scholar] [CrossRef]
- Cong, L.; Dong, X.; Wang, Y.; Deng, Y.; Li, B.; Dai, R. On the role of synthesized hydroxylated chalcones as dual functional amyloid-β aggregation and ferroptosis inhibitors for potential treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 166, 11–21. [Google Scholar] [CrossRef]
- Fang, Y.; Zhou, H.; Gu, Q.; Xu, J. Synthesis and evaluation of tetrahydroisoquinoline-benzimidazole hybrids as multifunctional agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 167, 133–145. [Google Scholar] [CrossRef]
- González-Naranjo, P.; Pérez-Macias, N.; Pérez, C.; Roca, C.; Vaca, G.; Girón, R.; Sánchez-Robles, E.; Martín-Fontelles, M.I.; de Ceballos, M.L.; Martin-Requero, A.; et al. Indazolylketones as new multitarget cannabinoid drugs. Eur. J. Med. Chem. 2019, 166, 90–107. [Google Scholar] [CrossRef]
- Wang, D.; Hu, M.; Li, X.; Zhang, D.; Chen, C.; Fu, J.; Shao, S.; Shi, G.; Zhou, Y.; Wu, S.; et al. Design, synthesis, and evaluation of isoflavone analogs as multifunctional agents for the treatment of Alzheimer’s disease. Eur. J. Med. Chem. 2019, 168, 207–220. [Google Scholar] [CrossRef]
- Dos Santos, T.C.; Gomes, T.M.; Pinto, B.A.S.; Camara, A.L.; Paes, A.M.A. Naturally occurring acetylcholinesterase inhibitors and their potential use for Alzheimer’s disease therapy. Front. Pharmacol. 2018, 9, 1192. [Google Scholar] [CrossRef] [Green Version]
- Llorens-Martín, M.; Jurado, J.; Hernández, F.; Avila, J. GSK-3β, a pivotal kinase in Alzheimer disease. Front. Mol. Neurosci. 2014, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, J.R.M.; Marques, D.F.F.; Baptista, S.J.; Pereira, C.M.F.; Moreira, P.I.; Dinis, T.C.P.; Santos, A.E.; Salvador, J.A.R. Highlights in BACE1 inhibitors for Alzheimer’s disease treatment. Front. Chem. 2018, 6, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, J.J.; Noristani, H.N.; Verkhratsky, A. The serotonergic system in ageing and Alzheimer’s disease. Prog. Neurobiol. 2012, 99, 15–41. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, L.; Karelson, M.; Dobchev, D.A. Identification of natural compounds against neurodegenerative diseases using in silico techniques. Molecules 2018, 23, 1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shaw, D.E.; Shelley, M.; et al. Glide: A new approach for rapid, accurate docking and scoring method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Congreve, M.; Carr, R.; Murray, C.; Jhoti, H. A ‘rule of three’ for fragment-based lead discovery? Drug Discov. Today 2003, 8, 876–877. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- JChem for Office. 19.18.0, 2019, ChemAxon. Available online: http://www.chemaxon.com (accessed on 1 September 2019).
- Mao, F.; Wang, H.; Ni, W.; Zheng, X.; Wang, M.; Bao, K.; Ling, D.; Li, X.; Xu, Y.; Zhang, H.; et al. Design, synthesis, and biological evaluation of orally available first-generation dual-target selective inhibitors of Acetylcholinesterase (AChE) and Phosphodiesterase 5 (PDE5) for the treatment of Alzheimer’s disease. ACS Chem. NeuroSci. 2018, 9, 328–345. [Google Scholar] [CrossRef]
- Cheung, J.; Rudolph, M.J.; Burshteyn, F.; Cassidy, M.S.; Gary, E.N.; Love, J.; Franklin, M.C.; Height, J.J. Structures of human acetylcholinesterase in complex with pharmacologically important ligands. J. Med. Chem. 2012, 55, 10282–10286. [Google Scholar] [CrossRef]
- Gálvez, J.; Polo, S.; Insuasty, B.; Gutiérrez, M.; Cáceres, D.; Alzate-Morales, J.H.; De-la-Torre, P.; Quiroga, J. Design, facile synthesis, and evaluation of novel spiro- and pyrazolo [1,5-c]quinazolines as cholinesterase inhibitors: Molecular docking and MM/GBSA studies. Comput. Biol. Chem. 2018, 74, 218–229. [Google Scholar] [CrossRef]
- Neumann, U.; Ufer, M.; Jacobson, L.H.; Rouzade-Dominguez, M.L.; Huledal, G.; Kolly, C.; Luond, R.M.; Machauer, R.; Veenstra, S.J.; Hurth, K.; et al. The BACE-1 inhibitor CNP520 for prevention trials in Alzheimer’s disease. EMBO Mol. Med. 2018, 10, e9316. [Google Scholar] [CrossRef]
- Rueeger, H.; Lueoend, R.; Rogel, O.; Rondeau, J.-M.; Möbitz, H.; Machauer, R.; Jacobson, L.; Staufenbiel, M.; Desrayaud, S.; Neumann, U. Discovery of cyclic sulfone hydroxyethylamines as potent and selective β-Site APP-Cleaving Enzyme 1 (BACE1) inhibitors: Structure-based design and in vivo reduction of amyloid β-peptides. J. Med. Chem. 2012, 55, 3364–3386. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Li, L.; Chen, W.; Chen, T.; Ma, L.; Wang, X.; Xiong, B.; Xu, Y.; Shen, J. Virtual screening and structure-based discovery of indole acylguanidines as potent β-secretase (BACE1) inhibitors. Molecules 2013, 18, 5706–5722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, F.F.; Benajiba, L.; Campbell, A.J.; Weïwer, M.; Sacher, J.R.; Gale, J.P.; Ross, L.; Puissant, A.; Alexe, G.; Conway, A.; et al. Exploiting an Asp-Glu “switch” in glycogen synthase kinase 3 to design paralog-selective inhibitors for use in acute myeloid leukemia. Sci. Transl. Med. 2018, 10, eaam8460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.H.; Chen, J.M.; Normandin, M.D.; Chang, J.S.; Chang, G.C.; Taylor, C.K.; Trapa, P.; Plummer, M.S.; Para, K.S.; Conn, E.L.; et al. Discovery of a highly selective glycogen synthase kinase-3 inhibitor (PF-04802367) that modulates Tau phosphorylation in the brain: Translation for PET neuroimaging. Angew. Chem. Int. Ed. Engl. 2016, 55, 9601–9605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivaprakasam, P.; Han, X.; Civiello, R.L.; Jacutin-Porte, S.; Kish, K.; Pokross, M.; Lewis, H.A.; Ahmed, N.; Szapiel, N.; Newitt, J.A.; et al. Discovery of new acylaminopyridines as GSK-3 inhibitors by a structure guided in-depth exploration of chemical space around a pyrrolopyridinone core. Bioorg. Med. Chem. Lett. 2015, 25, 1856–1863. [Google Scholar] [CrossRef]
- Davis, B.A.; Nagarajan, A.; Forrest, L.R.; Singh, S.K. Mechanism of paroxetine (paxil) inhibition of the serotonin transporter. Sci. Rep. 2016, 6, 23789. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.A.; Green, E.M.; Gouaux, E. X-ray structure and mechanism of the human serotonin transporter. Nature 2016, 532, 334–339. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.A.B.; Plenge, P.; Andersen, J.; Eildal, J.N.N.; Kristensen, A.S.; Bøgesø, K.P.; Loland, C.J. Structure-activity relationship studies of citalopram derivatives: Examining substituents conferring selectivity for the allosteric site in the serotonin transporter. Br. J. Pharmacol. 2016, 173, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Krout, D.; Rodriquez, M.; Brose, S.A.; Golovko, M.Y.; Henry, L.K.; Thompson, B.J. Inhibition of the serotonin transporter is altered by metabolites of selective serotonin and norepinephrine reuptake inhibitors and represents a caution to acute or chronic treatment paradigms. ACS Chem. Neurosci. 2017, 8, 1011–1018. [Google Scholar] [CrossRef]
- Coleman, J.A.; Gouaux, E. Structural basis for recognition of diverse antidepressants by the human serotonin transporter. Nat. Struct. Mol. Biol. 2018, 25, 170–175. [Google Scholar] [CrossRef]
- Arfeen, M.; Patel, R.; Khan, T.; Bharatam, P.V. Molecular dynamics simulation studies of GSK-3β ATP competitive inhibitors: Understanding the factors contributing to selectivity. J. Biomol. Struct. Dyn. 2015, 33, 2578–2593. [Google Scholar] [CrossRef] [PubMed]
- Gaulton, A.; Hersey, A.; Nowotka, M.; Bento, A.P.; Chambers, J.; Mendez, D.; Mutowo, P.; Atkinson, F.; Bellis, L.J.; Cibrián-Uhalte, E.; et al. The ChEMBL database in 2017. Nucleic Acids Res. 2017, 45, D945. [Google Scholar] [CrossRef] [PubMed]
- RCSB Protein Data Bank. Available online: https://rcsb.org (accessed on 20 December 2019).
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, J.A.; Thieffine, S.; Vulpetti, A.; Cristiani, C.; Valsasina, B.; Knapp, S.; Kalisz, H.M.; Flocco, M. Structural characterization of the GSK-3beta active site using selective and non-selective ATP-mimetic inhibitors. J. Mol. Biol. 2003, 333, 393–407. [Google Scholar] [CrossRef]
- Sastry, G.M.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput. Aid. Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Schrödinger Release 2016-3: Schrödinger Suite 2016-3 Protein Preparation Wizard; Epik Schrödinger LLC; Impact Schrödinger LLC;Prime, Schrödinger, LLC: New York, NY, USA, 2016.
- Irwin, J.J.; Sterling, T.; Mysinger, M.M.; Bolstad, E.S.; Coleman, R.G. ZINC: A free tool to discover chemistry for biology. J. Chem. Inf. Model. 2012, 52, 1757–1768. [Google Scholar] [CrossRef]
- Schrödinger Release 2018-1: LigPrep; Schrödinger, LLC: New York, NY, USA, 2018.
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger Release 2018-1: Glide; Schrödinger, LLC: New York, NY, USA, 2018.
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable algorithms for molecular dynamics simulations on commodity clusters. SC ’06. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; p. 43. [Google Scholar] [CrossRef] [Green Version]
- Zielkiewicz, J. Structural properties of water: Comparison of the SPC, SPCE, TIP4P, and TIP5P models of water. J. Chem. Phys. 2006, 123, 104501, J. Chem. Phys.2006, 124, 109901. [Google Scholar] [CrossRef]
- Banks, J.L.; Beard, H.S.; Cao, Y.; Cho, A.E.; Damm, W.; Farid, R.; Felts, A.K.; Halgren, T.A.; Mainz, D.T.; Maple, J.R.; et al. Integrated modeling program, applied chemical theory (IMPACT). J. Comput. Chem. 2005, 26, 1752–1780. [Google Scholar] [CrossRef] [Green Version]
- Martyna, G.J.; Klein, M.L. Nosé–hoover chains: The canonical ensemble via continuous dynamics. J. Chem. Phys. 1992, 97, 2635–2643. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open babel: An open chemical toolbox. J. Cheminformatics 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halgren, T.A. Merck molecular force field. I. Basis, form, scope, parameterization, and performance of MMFF94. J. Comput. Chem. 1996, 17, 490–519. [Google Scholar] [CrossRef]
- Stewart, J.J.P. MOPAC: A semiempirical molecular orbital program. J. Comput. Aided Mol. Des. 1990, 4, 1–103. [Google Scholar] [CrossRef] [PubMed]
- Karelson, M.; Dobchev, D.A.; Karelson, G.; Tamm, T.; Tämm, K.; Nikonov, A.; Mutso, M.; Merits, A. Fragment-based development of HCV protease inhibitors for the treatment of hepatitis C. Curr. Comput Aided Drug Des. 2012, 8, 55–61. [Google Scholar] [CrossRef]
- Karelson, M. Molecular Descriptors in QSAR/QSPR; John Wiley & Sons Inc. Publication: New York, NY, USA, 2000; ISBN 978-0-471-35168-9. [Google Scholar]
- Katritzky, A.R.; Mu, L.; Lobanov, V.S.; Karelson, M. Correlation of boiling points with molecular structure. 1. A training set of 298 diverse organics and a test set of 9 simple inorganics. J. Phys. Chem. 1996, 100, 10400–10407. [Google Scholar] [CrossRef]
- Katritzky, A.R.; Kuanar, M.; Slavov, S.; Hall, C.D.; Karelson, M.; Kahn, I.; Dobchev, D.A. Quantitative correlation of physical and chemical properties with chemical structure: Utility for prediction. Chem. Rev. 2010, 110, 5714–5789. [Google Scholar] [CrossRef]
- Karelson, M.; Karelson, G.; Tamm, T.; Tulp, I.; Jänes, J.; Tämm, K.; Lomaka, A.; Savchenko, D.; Dobchev, D.A. QSAR study of pharmacological permeabilities. Arkivoc 2009, 2, 218–238. [Google Scholar] [CrossRef] [Green Version]
- Dobchev, D.A.; Pillai, G.G.; Karelson, M. In silico machine learning methods in drug development. Curr. Top Med. Chem. 2014, 14, 1913–1922. [Google Scholar] [CrossRef]
- Haykin, S. Neural Networks a Comprehensive Foundation; Pearson: London, UK, 1999; ISBN 81-7808-300-0. [Google Scholar]
- Guha, R.; Stanton, D.T.; Jurs, P.C. Interpreting computational neural network quantitative structure-property relationship models: A detailed interpretation of the weights and biases. J. Chem. Inf. Model. 2005, 45, 1109–1121. [Google Scholar] [CrossRef]
- MolPort, Lacplesa iela 41, Riga, LV-1011, Latvia. Available online: http://www.molport.com/shop/index (accessed on 10 January 2020).
- La Jolla California USA. Available online: www.graphpad.com (accessed on 8 March 2020).
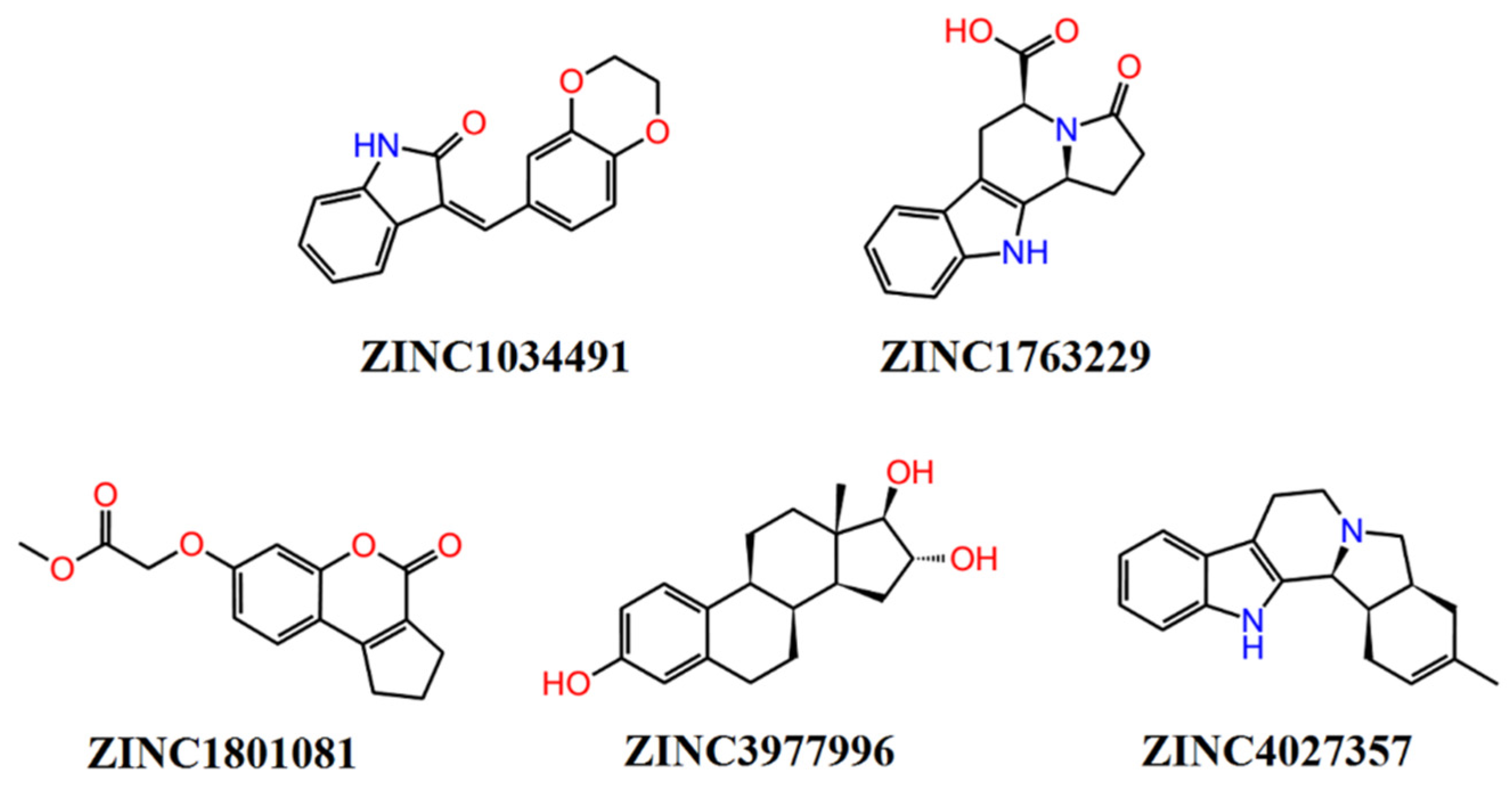
Sample Availability: Samples of the compounds ZINC1034491, ZINC4027357, ZINC3977996, ZINC1801081, and ZINC1763229 are available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AChE BMLR Model—Log (IC50) = D0 + ∑(Bi ± ErrorsBi)*Di | |||||
|---|---|---|---|---|---|
| N = 239, R2 = 0.932026, R2cv = 0.929713, R2abc = 0.929519, s2 = 0.0808643, F = 802.121 | |||||
| Descriptor Di | Bi | Errors Bi | ti-Statistics | Descriptors | |
| 0 | 2.880 | 0.051 | 56.989 | Intercept | |
| 1 | 0.312 | 0.008 | 38.327 | Lowest resonance energy (AM1) for N–S bonds | |
| 2 | −2.475 | 0.184 | −13.462 | Max net atomic charge (AM1) for O atoms | |
| 3 | 0.174 | 0.009 | 19.494 | Final heat of formation (AM1)/# atoms | |
| 4 | −0.014 | 0.001 | −14.213 | Highest e-e repulsion (AM1) for N–H bonds | |
| ABC Crossvalidation | |||||
| (AB,C): R2ab = | 0.923 | R2ab_cv = | 0.919 | R2c_pred = | 0.944 |
| (BC,A): R2bc = | 0.932 | R2bc_cv = | 0.929 | R2a_pred = | 0.930 |
| (CA,B): R2ca = | 0.938 | R2ca_cv = | 0.935 | R2b_pred = | 0.915 |
| XY-Randomizations | |||||
| X-scrambling R2 = | 0.0170677 | ||||
| Y-scrambling R2 = | 0.0170352 | ||||
| XY-scrambling R2 = | 0.0175535 | ||||
| BACE1 BMLR Model | |||||
| N = 378, R2 = 0.908694, R2cv = 0.90621, R2abc = 0.903581, s2 = 0.0761742, F = 928.037 | |||||
| 0 | 23.461 | 0.735 | 31.898 | Intercept | |
| 1 | 0.001 | 0.000 | 10.494 | PPSA2 Total charge weighted PPSA (AM1) | |
| 2 | −0.182 | 0.003 | −55.132 | Structural Information content (order 2) | |
| 3 | −4.527 | 0.217 | −20.886 | Average valency (AM1) | |
| 4 | −2.230 | 0.129 | −17.320 | Balaban index | |
| ABC Crossvalidation | |||||
| (AB,C): R2ab = | 0.905 | R2ab_cv = | 0.901 | R2c_pred = | 0.909 |
| (BC,A): R2bc = | 0.912 | R2bc_cv = | 0.908 | R2a_pred = | 0.897 |
| (CA,B): R2ca = | 0.908 | R2ca_cv = | 0.904 | R2b_pred = | 0.905 |
| XY-Randomizations | |||||
| X-scrambling R2 = | 0.0106888 | ||||
| Y-scrambling R2 = | 0.0106174 | ||||
| XY-scrambling R2 = | 0.0110244 | ||||
| GSK3β BMLR Model | |||||
| N = 229, R2 = 0.900783, R2cv = 0.896404, R2abc = 0.900, s2 = 0.0737752, F = 508.42 | |||||
| 0 | 6.432 | 0.142 | 45.152 | Intercept | |
| 1 | −0.593 | 0.017 | −34.239 | HA dependent HDCA-2 (AM1) | |
| 2 | 1.569 | 0.058 | 27.210 | LUMO energy (AM1) | |
| 3 | 0.710 | 0.036 | 19.703 | HACA-2 (Zefirov) | |
| 4 | 11.621 | 0.694 | 16.746 | Min net atomic charge (Zefirov) for any atom type | |
| ABC Crossvalidation | |||||
| (AB,C): R2ab = | 0.902 | R2ab_cv = | 0.896 | R2c_pred = | 0.899 |
| (BC,A): R2bc = | 0.900 | R2bc_cv = | 0.893 | R2a_pred = | 0.896 |
| (CA,B): R2ca = | 0.894 | R2ca_cv = | 0.887 | R2b_pred = | 0.907 |
| XY-Randomizations | |||||
| X-scrambling R2 = | 0.0181116 | ||||
| Y-scrambling R2 = | 0.0171723 | ||||
| XY-scrambling R2 = | 0.0173102 | ||||
| SERT BMLR Model | |||||
| N = 213, R2 = 0.846334, R2cv = 0.838471, R2abc = 0.844291, s2 = 0.0767979, F = 286.397 | |||||
| 0 | 154.961 | 10.036 | 15.440 | Intercept | |
| 1 | 0.598 | 0.021 | 27.881 | Kier shape index (order 3) | |
| 2 | −3.882 | 0.255 | −15.202 | Lowest n-n repulsion (AM1) | |
| 3 | −0.119 | 0.005 | −22.512 | Bonding Information content (order 1) | |
| 4 | 7.575 | 0.452 | 16.740 | FHASA Fractional HASA (HASA/TMSA) (AM1) | |
| ABC Crossvalidation | |||||
| (AB,C): R2ab = | 0.839 | R2ab_cv = | 0.826 | R2c_pred = | 0.857 |
| (BC,A): R2bc = | 0.839 | R2bc_cv = | 0.827 | R2a_pred = | 0.833 |
| (CA,B): R2ca = | 0.852 | R2ca_cv = | 0.840 | R2b_pred = | 0.843 |
| XY-Randomizations | |||||
| X-scrambling R2 = | 0.018852 | ||||
| Y-scrambling R2 = | 0.0190767 | ||||
| XY-scrambling R2 = | 0.0183609 | ||||
| Target | Predicted Log (IC50) (nM) | |||
|---|---|---|---|---|
| ANN | MLR | |||
| min | max | min | max | |
| AChE | 1.975 | 3.882 | 3.407 | 4.082 |
| BACE1 | 3.149 | 3.958 | 2.452 | 5.280 |
| GSK3β | 2.385 | 4.166 | 1.548 | 9.760 |
| SERT | 0.567 | 1.737 | −0.770 | 3.799 |
| Compound | Binding Energy, ∆G, kcal/mol | Ligand Efficiency | Compound | Binding Energy, ∆G, kcal/mol | Ligand Efficiency |
|---|---|---|---|---|---|
| AChE | GSK3β | ||||
| (−)-huperzine A | −7.9 | 0.44 | 2WF | −7.1 | 0.32 |
| galantamine | −4.9 | 0.23 | BRD0209 | −6.9 | 0.27 |
| donepezil (DPZ) | −10.2 | 0.36 | PF-04802367 | −6.9 | 0.28 |
| ZINC1034491 | −9.8 | 0.47 | ZINC1034491 | −8.7 | 0.41 |
| ZINC4027357 | −9.3 | 0.44 | ZINC4027357 | −8.6 | 0.41 |
| ZINC3977996 | −9.2 | 0.44 | ZINC3977996 | −9.1 | 0.43 |
| ZINC1763229 | −9.2 | 0.46 | ZINC1763229 | −8.0 | 0.40 |
| ZINC1801081 | −9.1 | 0.46 | ZINC1801081 | −8.1 | 0.41 |
| BACE1 | SERT | ||||
| CNP520 | −10.7 | 0.31 | Paroxetine | −10.8 | 0.45 |
| VTI | −9.9 | 0.34 | S-citalopram | −9.5 | 0.39 |
| NVP-BXD552 | −9.2 | 0.23 | sertraline | −9.1 | 0.46 |
| ZINC1034491 | −10.0 | 0.48 | ZINC1034491 | −10.3 | 0.49 |
| ZINC4027357 | −10.0 | 0.48 | ZINC4027357 | −10.3 | 0.49 |
| ZINC3977996 | −10.2 | 0.49 | ZINC3977996 | −10.1 | 0.48 |
| ZINC1763229 | −8.9 | 0.45 | ZINC1763229 | −8.9 | 0.44 |
| ZINC1801081 | −9.6 | 0.48 | ZINC1801081 | −8.8 | 0.44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, L.; Karelson, M.; Dobchev, D.A. Multitarget Approach to Drug Candidates against Alzheimer’s Disease Related to AChE, SERT, BACE1 and GSK3β Protein Targets. Molecules 2020, 25, 1846. https://doi.org/10.3390/molecules25081846
Ivanova L, Karelson M, Dobchev DA. Multitarget Approach to Drug Candidates against Alzheimer’s Disease Related to AChE, SERT, BACE1 and GSK3β Protein Targets. Molecules. 2020; 25(8):1846. https://doi.org/10.3390/molecules25081846
Chicago/Turabian StyleIvanova, Larisa, Mati Karelson, and Dimitar A. Dobchev. 2020. "Multitarget Approach to Drug Candidates against Alzheimer’s Disease Related to AChE, SERT, BACE1 and GSK3β Protein Targets" Molecules 25, no. 8: 1846. https://doi.org/10.3390/molecules25081846
APA StyleIvanova, L., Karelson, M., & Dobchev, D. A. (2020). Multitarget Approach to Drug Candidates against Alzheimer’s Disease Related to AChE, SERT, BACE1 and GSK3β Protein Targets. Molecules, 25(8), 1846. https://doi.org/10.3390/molecules25081846