Abstract
The inhibition of viral protease is an important target in antiviral drug discovery and development. To date, protease inhibitor drugs, especially HIV-1 protease inhibitors, have been available for human clinical use in the treatment of coronaviruses. However, these drugs can have adverse side effects and they can become ineffective due to eventual drug resistance. Thus, the search for natural bioactive compounds that were obtained from bio-resources that exert inhibitory capabilities against HIV-1 protease activity is of great interest. Fungi are a source of natural bioactive compounds that offer therapeutic potential in the prevention of viral diseases and for the improvement of human immunomodulation. Here, we made a brief review of the current findings on fungi as producers of protease inhibitors and studies on the relevant candidate fungal bioactive compounds that can offer immunomodulatory activities as potential therapeutic agents of coronaviruses in the future.
1. Introduction
Coronaviruses (CoVs) are a large group of enveloped viruses with non-segmented, single-strand, and positive-sense RNA genomes. CoVs are classified in the family Coronaviridae of the order Nidovirales. Notably, CoVs have been identified as zoonotic viruses that can be transmitted between humans and animals and they are known to cause a wide range of infections. These infections can appear as symptoms that range from those of the common cold to much more fatal diseases, like respiratory syndrome, as well as enteric and central nervous system diseases. Two highly pathogenic microorganisms with approximately 30,000 nucleotides, the Severe Acute Respiratory Syndrome (SARS-CoV, or SARS) and the Middle East Respiratory Syndrome (MERS-CoV, or MERS), have resulted in regional and global outbreaks. In 2002, SARS originated in southern China, while MERS was first known to infect a patient in Saudi Arabia in 2012 [1,2,3]. A novel coronavirus, which was previously designated as SARS-CoV-2, was identified as a causal agent of pneumonia in Wuhan, a city in the Hubei Province, China, at the end of 2019 [4]. It has subsequently spread throughout China and elsewhere and it is now considered a global health emergency. In February 2020, the World Health Organization (WHO) labeled the disease SARS-CoV-2 or 2019-nCoV, which has been more commonly referred to as the coronavirus disease since its emergence in 2019. The mortality rate of SARS-CoV-2 infection has been seen to be around two percent in China, which is much less than the mortality rates of SARS-CoV and MERS-CoV infection [5]. However, it has caused global concern by its efficient human-to-human transmission, leading to its widespread outbreak in many countries around the world [4,5,6]. Currently, the WHO has referred to the SARS-CoV-2 outbreak as a “pandemic”, as emphasized the global risk of its spread and predictive elevates the risk of its impact to “very high”. Clinical practice guidelines and the treatment protocols of WHO and the Center for Disease Control and Prevention (CDC) for a patient infected with SARS-CoV-2 are similar to those of other viral causes of pneumonia. These include prompt supportive care, like oxygen therapy, fluid management, empiric antimicrobials (in case of sepsis), and others [5].
2. Protease Inhibitor Drugs for CoVs
Generally, the inhibition of the viral replication process is of significant consideration in the treatment of viral infections (Figure 1). Protease is one of the necessary enzymes required for the replication, transcription, and maturation of a range of viruses [7,8]. Several studies have focused on the identification of an inhibitory target of protease, which is necessary for viral transcription/replication. Currently, the approved protease inhibitors are recognized as peptidomimetics and they are one of the first examples of a structure-based drug design that utilizes the structural information of inhibitor binding to the active site of viral protease [9,10].
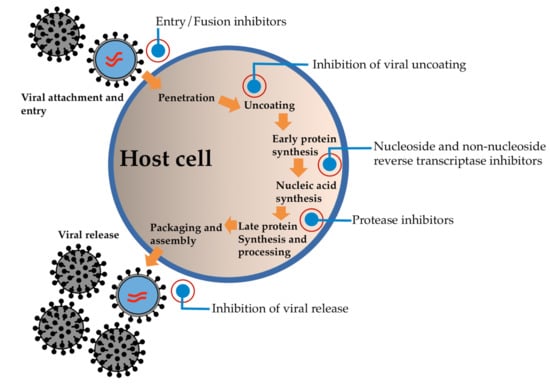
Figure 1.
Major sites of antiviral drug action.
Protease inhibitors play an important role in viral replication by selectively binding to viral proteases and blocking proteolytic cleavage of the protein precursors that are necessary for the production of infectious viral particles [7,8,11]. The papain-like protease (PLpro) and the 3-chymotrypsin-like protease (3CLpro, also known as the main proteases that are suitable targets for viral inhibitors) have been identified in CoVs, for which both proteases are believed to be essential in the role of viral replication and are considered to be attractive targets for antiviral therapeutics [12,13]. Numerous previous studies have identified compounds and drugs that can inhibit protease activity through docking/molecular dynamic experimentation and their inhibition activity on CoVs replication in the cell cultures of mice and in non-human primate (NHP) models, as has been previously reported [14]. On the other hand, human immunodeficiency virus type 1 (HIV-1) protease inhibitors (tipranavir, saquinavir, ritonavir, nelfinavir, lopinavir, indinavir, darunavir, atazanavir, and amprenavir) that have been approved for clinical applications by the Food and Drug Administration (FDA) are widely reported to be able to deactivate 3CLpro. Hence, they have been identified as potential drugs in the treatment of CoV infections. Lopinavir, atazanavir and indinavir have been identified as potential candidates as 3CLpro inhibitors [15,16]. Furthermore, RNA-dependent RNA polymerase inhibitors, e.g., remdesivir and favilavir, have been used to effectively treat CoVs infections [16,17]. Remdesivir or chloroquine, glucocorticoids and the combined protease inhibitor lopinavir-ritonavir have been used to treat SARS-CoV and MERS-CoV infections [4,16,17,18].
Currently, there is a lack of an effective treatment or a vaccine to prevent SARS-CoV-2 infection. Parent compounds are being tested to prevent SARS-CoV-2 infection in in vitro or clinical studies-based SARS-CoV and MERS-CoV trials. Remdesivir or chloroquine has been highly effective in the inhibition of SARS-CoV-2 infection in vitro [18]. Lopinavir-ritonavir has been suggested to be one of the therapeutic agents of SARS-CoV-2 [17,19,20]. Hence, presently, there are no approved vaccines or drugs for SARS-CoV-2 infection. Potential candidates as targets for further in vitro and in vivo studies for SARS-CoV-2 prevention and treatment are 3CLpro, Spike, RNA-dependent RNA polymerase (RdRp), PLpro and human angiotensin-converting enzyme 2 (human ACE2). Interestingly, the potential unitality of clinical drugs and natural products for the treatment of SARS-CoV-2 infection was studied while using computational methods. Specifically, 3CLpro is recognized as an important target anti-SARS-CoV-2 drug that was designed and modified to play an important role in the maturation of SARS-CoV-2. Nelfinavir has been predicted as a potential inhibitor of the SARS-CoV-2 main protease, but has been used in treatment application [4]. Nevertheless, favilavir has been approved for use in the investigational therapy for SARS-CoV-2 infected patients in China [21]. The US patient infected with SARS-CoV-2 has been treated by supportive care and remdesivir, for which no adverse reactions were observed during administration [22,23]. In February 2020, the remdesivir treatment of SARS-CoV-2 was initiated in Wuhan and Beijing, China for clinical trials, but the safety and efficacy of remdesivir in the treatment process need to be fully evaluated [24]. Therefore, this study has summarized the current findings on the natural antiviral compounds that were obtained from fungi for the purpose of employing them as protease inhibitors. The findings reveal a noteworthy potential in the development of antiviral agents for CoVs. Moreover, the fungal bioactive compounds that possess immunomodulatory activities reveal themselves to be a potential resource in the treatment of CoVs.
3. Potential of Fungal Antiviral Bioactive Compounds as Protease Inhibitors to Treat CoVs
Fungi (including filamentous fungi and mushrooms) represent a rich source of various biologically active compounds that can serve as a major source of new compounds in the development of small-molecule drugs. This development process could involve direct or semi-synthetic methods, while the findings of this research could serve as a source of inspiration in the investigation of chemical scaffolds. Bioactive compounds that were obtained from fungi with potent antiviral activity are presently under investigation, and the number of studies is continually increasing [25,26,27,28,29,30]. Fungal bioactive compounds can be divided into two major groups of molecules; small organic molecules (secondary metabolites) produced by filamentous fungi, especially endophytic fungi, and high molecular weight compounds in the extracts or products that were obtained from the fruiting bodies of edible or medicinal mushrooms [26,31,32,33]. Fungal small organic molecules are low molecular weight compounds that are produced by filamentous fungi. These compounds were synthesized by fungal hyphae and later secreted. They are commonly studied through the cultivation of fungal hyphae in culture media. Small molecular weight compounds with antiviral capabilities have been classified as indole alkaloids, non-ribosomal peptides, polyketides, and hybrids of non-ribosomal peptides and polyketides, and terpenoids [24,25,30,31,33,34,35,36,37,38]. The antiviral activity of high molecular weight compounds that are extracted from fruiting bodies and fungal mycelia have been reported and classified as lignin derivatives, polysaccharides (e.g. chitin, glucan, lentinan, and mannan), proteins, and polysaccharide-protein/amino acid complexes [28,29,30,39,40,41,42]. Drugs or compounds with special effects on viral protease inhibitors, like HIV-1 protease inhibitors and hepatitis C virus (HCV) NS3/4A protease inhibitors, have been considered as potential drugs against CoVs, according to the findings of previous studies. Therefore, fungal compounds that have the potential to be candidates as protease inhibitors have been the focus of numerous present studies.
The major viral protease classes have been identified based on the relevant catalytic types including serine, cysteine, aspartic, threonine, glutamic acid, and metalloproteases [43,44]. Most viral proteases can recognize the specific sequences of amino acids in their substrates and cleave the peptide bond via a nucleophilic attack on the side chain of catalytic site [43,44,45]. HIV-1 protease is formed by two identical monomers as shown in Figure 2. Each monomer contributes on catalytic aspartyl residues (Asp25 and Asp25ʹ) in the active site which lie on the bottom of the cavity that plays a crucial role in substrate binding [46]. Additionally, HCV polyprotein is processed proteolytically upon translation by both host cells and viral proteases to at least 10 individual proteins [47,48]. These include four structural proteins and non-structural (NS) proteins. NS3/4A serine protease is further involved in the proteolytic processing of NS proteins and is also considered necessary for the direct-inhibition of HCV. NS3 is comprised of protease and helicase domains and forms a heterodimer with NS4A. Additionally, NS4A binds to the N-terminal region of NS3 and acts as a cofactor of the protease to enhance cleavage (Figure 2). The catalytic triad of the NS3/4A protease is formed by His57, Asp81, and Ser139 [47,48,49]. Currently, the NS3 protein has emerged as an important target for anti-HCV drug discovery and development. Notably, a 3CLpro of CoVs is comprised of three-domain cysteine proteases. Furthermore, domain I and II contain β-barrels of the chymotrypsin structure, but domain III consists mainly of α-helices (Figure 2). Moreover, 3CLpro contains a catalytic dyad defined by His41 and Cys145 [50,51]. This main protease is responsible for maturation of functional proteins and currently represents as a key target for antiviral drugs. Therefore, most of antiviral agents are peptidomimetics and macrocyclic compounds that interact with the active site of a targeted viral protease [52,53]. According to the HIV-1 protease and HCV protease exhibited a similar function as CoVs protease, so protease inhibitors are hypothesized to have the preventive and therapeutic potential against CoVs infection.
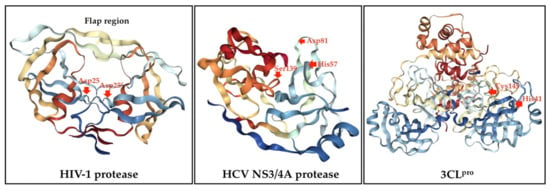
Figure 2.
Crystal structures of HIV-1 protease (PDB: 2NMZ), HCV NS3/4A protease (PDB: 1DY8) and 3CLpro (PDB: 2DUC). The catalytic sites are arrowed.
3.1. HIV-1 Protease Inhibitors Isolated from Fungi
Several HIV-1 protease inhibitor drugs have been made available in the human clinical use of CoVs [4,7,8,16,17,18]. Nelfinavir was found to strongly inhibit the replication of SARS-CoV [54,55]. Antiviral drugs (ribavirin, lopinavir, and ritonavir), steroids, proteins that are known as immunoglobulins, type I interferon, and convalescent plasma have been used in the clinical treatment of SARS and MERS patients [56,57,58,59,60]. A diagnostic test for early SARS and MERS illnesses has not been validated; therefore, treatment could only be initiated once patients have met the criteria of a clinical and epidemiological case definition. Patient characteristics, such as age and the presence of diabetes, have been associated with severe diseases and they can confound treatment effects. Certain potential drugs that have been approved by the FDA and identified as potential inhibitors of 3CLpro of SARS-CoV-2 have been reported by Hosseini and Amanlou [11] while using a virtual screening and the molecular docking procedure. The ten potential drugs include paclitaxel, simeprevir, docetaxel, palbociclib, cabazitaxel, alctinib, imatinib, plerixafor, azelastine, and dasabuvir. Paclitaxel and simeprevir HCV NS3/4A protease inhibitors) revealed a strong degree of interaction with the SARS-CoV-2 protease binding pocket and it has been placed well into the pocket when compared to the antiviral drugs. Interestingly, virtual screening has confirmed that indinavir was selected as the SARS-CoV-2 main protease (PDB code 6LU7).
HIV-1 protease inhibition has been most thoroughly tested by purified and unpurified fungal metabolites. Table 1 and Figure 2 show fungal bioactive agents for HIV-1 protease inhibitors are shown in Table 1 and Figure 3. Interestingly, paclitaxel or taxol, a chemotherapeutic diterpenoid natural compound, was first extracted from the bark of trees that belong to the genus Taxus. This compound was produced by several endophytic fungi in the genera Alternaria, Aspergillus, Beauveria, Cladosporium, Chaetomella, Fusarium, Guignadia, Monochaetia, Nodulisporium, Pestlotia, Pestalotiopsis, Pithomyces, Penicillium, Phomopsis, Phyllostica, Sporormia, Taxomyces, Trichoderma, Trichothecium, Tubercularia, and Xylaria [61,62,63,64,65,66,67,68,69]. More than sixty endophytic fungal strains have been identified as paclitaxel producers [70,71]. Generally, paclitaxel has been used as an anticancer drug against breast cancer, non-small cell lung cancer, ovarian cancer, and prostate cancer [72,73]. However, paclitaxel is now being considered for its inhibitory effect on HIV-1 protease activity.

Table 1.
Fungal bioactive compounds for HIV-1 protease inhibitors that potential candidate to treat CoVs.
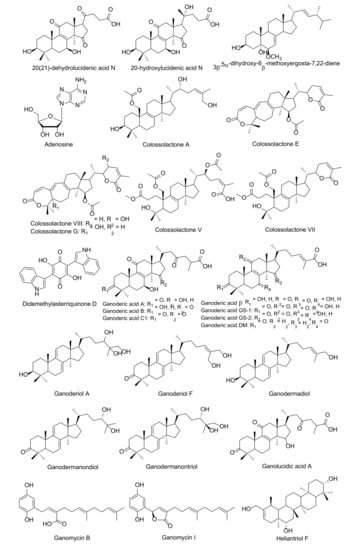
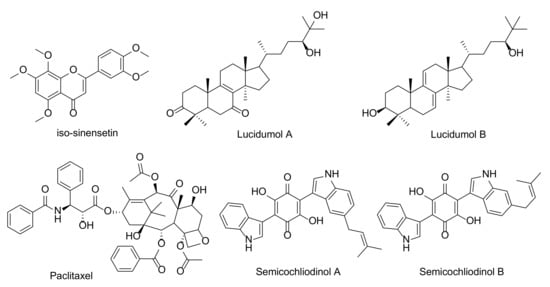
Figure 3.
Fungal bioactive compounds for inhibition of HIV-1 protease activity.
Ryang et al. [74] reported that 20 μg/mL of paclitaxel could inhibit HIV-1 protease activity in a similar manner to the positive control pepstatin A (80 μg/mL) in the in vitro experiment. A combination of paclitaxel and protease inhibitors (indinavir, nelfinavir, or combinations of these agents) at recommended dosages and schedules was used to treat patients of HIV-associated Kaposi’s Sarcoma without enhancing toxicity [75]. In the virtual screening procedure, paclitaxel has been suggested as a therapeutic agent of SARS-CoV-2 based on its higher binding energy (–11.33 kcal/mol) to the active site of SARS-CoV-2 protease than that of lopinavir (–5.36 kcal/mol) and ritonavir (–5.04 kcal/mol) [21]. However, patient conditions for paclitaxel applications should be considered because of its side effects on bone marrow suppression. In addition, two semicochliodinols (semicochliodinol A and B) and didemethylasterriquinone D that were isolated from a microfungus, Chrysosporium merdarium, displayed an inhibitory effect on HIV-1 protease activity [76,77].
Some HIV-1 protease inhibitors have been isolated from certain mushrooms, especially some edible and medicinal mushrooms. Lingzhi mushrooms (Ganoderma species) have been generally acknowledged as a nutritional supplement across the world due to their association with long-term safety and the fact that they possess a vast array of medicinal properties. Various compounds that have exhibited inhibitory effects against HIV-1 protease activity have been identified from Ganoderma lucidum including ganolucidic acid A, 3β-5α-dihydroxy-6β-methoxyergosta-7,22-diene, ganoderic acid A–C, ganoderic acid β, ganodermanondiol, ganodermanontriol and lucidumol B [78,79,80]. Six colossolactones, ganomycin I, and ganomycin B isolated from G. colosum have displayed anti-HIV-1 protease activity [81,82]. Twenty-five metabolites were isolated from the fruiting body of G. sinnense, and it was found that ganoderic acid GS-2, 20-hydroxylucidenic acid N, 20(21)-dehydrolucidenic acid N and ganoderiol F exhibited a high potential to inhibit HIV-1 protease activity [83]. Notably, crude extracts of tiger milk mushroom (Lignosus rhinocerus) displayed inhibitory activity against HIV-1 protease activity on infected cells, while in silico analysis showed that heliantriol F displayed significant binding energy at -12.57 kcal/mol on the active site of HIV-1 protease [84]. Hexane extract fractions obtained from a jelly fungus (Auricularia polytricha) could effectively inhibit HIV-1 protease activity in vitro, while four major compounds, ergosterol, linoleic acid and two triacylglycerols were found to be present [85]. Moreover, adenosine and iso-sinensetin isolated from golden cordycep (Cordycep militaris), and 4.5 kDa protein isolated from Russula paludosa, have been reported as anti-HIV-1 replications by inhibition of HIV-1 protease activity [86,87].
3.2. HCV NS3/4A Protease Inhibitors Isolated from Fungi
Simeprevir, which is a HCV NS3/4A protease inhibitor, has been acknowledged as a highly effective agent of SARS-CoV-2 that can display a higher energy value (–11.33 kcal/mol) for the binding active site of SARS-CoV-2 protease than lopinavir and ritonavir [20]. However, there has been an absence of clinical test support for this outcome. Additionally, patient conditions for simeprevir applications should be considered, because it can commonly cause a rash, nause, and muscle pain, as well as an allergic reaction [11]. In published literature, there are several bioactive compounds isolated from endophytic fungi and mushrooms that have been identified for the inhibition of HCV NS3/4A protease (Table 2 and Figure 4). An aqueous extract with a low molecular weight (< 3 kDa) fraction of the white button mushroom (Agaricus bisporus) has displayed a responsible activity to the indictors of HCV replication [88]. Alternaroil and alternariol derivatives (alternariol-9-methyl ether-3-O-sulphate and alternariol-9-methyl ether) of an endophytic fungus, Alternaria alternate, and their metabolites were explored for the inhibition of HCV NS3-NS4A protease [89,90,91,92]. Hawas et al. [93] found that the most potent HCV NS3/4A protease isolated compound that was obtained from Fusarium equiseti were ω-hydroxyemodin and Griseoxanthone C. Furthermore, mellein, patulin, and H1-A were isolated from Aspergillus ochraceus, Penicillium griseofulvum, and Fusarium oxysporum, respectively. These compounds displayed activity against HCV NS3/4A protease [94,95,96]. Antrodia cinnamomea, a medicinal mushroom, produced antrodins A–E. Antrodins A showed potent inhibitory capabilities of HCV protease activity [97]. Five products that were obtained from the endophytic fungus Emericella nidulans, namely cordycepin, emericellin, ergosterol peroxide, myristic acid, and sterigmatocystin, reported having HCV NS3/4A protease inhibitory properties [98,99,100,101,102]. Moreover, Ahmed et al. [103] isolated the metabolite compounds for HCV NS3/4A protease inhibitors that were obtained from Aspergillus versicolor that possess constituents of (−)-curvularin, cyclo(L-Pro-L-Ile), cyclo(L-Tyr-LPro), cyclo(L-Phe-L-Pro), cyclic tetrapeptide, and cyclo-(Phenylalanyl-pro-Leu-pro). Three metabolites were isolated from an endophytic fungus, P. chrysogenum [104]. These compounds were identified as alatinone, emodin, and ω-hydroxyemodin, and they displayed activities against HCV NS3/4A protease.
Table 2.
Fungal bioactive compounds for HCV NS3/4A protease inhibitor as potential candidates for the treatment of CoVs, particularly SARS-CoV-2.
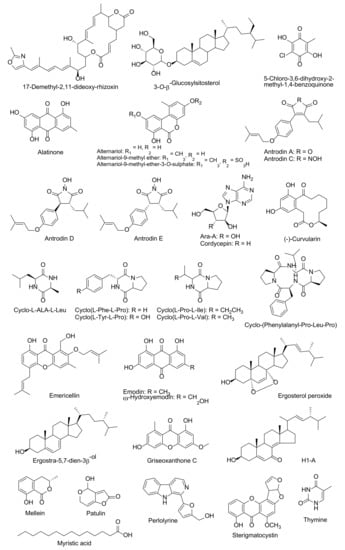
Figure 4.
Fungal bioactive compounds for inhibition of HCV NS3/4A protease.
4. Potential of Fungal Bioactive Compounds for Immunomodulators
Inflammasome is a cytosolic multiprotein oligomer of the innate immune system that is responsible for the activation of inflammatory responses. Inflammasome induction by coronavirus was first reported in porcine reproductive and respiratory syndrome virus [105]. Currently, the transport of Ca2+ by SARS-CoV has been reported to trigger inflammasome activation. It has been suggested that the cytokine storm is associated with cases of pneumonia that were infected by SARS-CoV-2 [106]. Cytokines and chemokines have been recognized for playing an important role in immunity and immunopathology in the body during virus infection. They are an important part of the first barrier of innate immunity that serves as a defense against the viruses. The massive infiltrated inflammatory cells and the elevated proinflammatory cytokines/chemokines can lead to fatal acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) [107,108]. A clinical study of 41 patients infected with SARS-CoV-2 in Wuhan, China showed that 63% of the patients had lymphopenia, 12% had ARDS, all patients had pneumonia, and the intensive care patients reported higher plasma levels of IL-2, IL-7, IL-10, GSCF, IP10, MCP1, MIP1A, and TNF-α than the non-intensive care patients [108]. Researchers also noted that patients with high concentrations of IL-1β, IFN-γ, IP10, and MCP1 were likely associated with activated T-helper-1 (Th1) cell responses.
Immunomodulators are the bioactive substances that can play a role or affect the regulating of the immune system, which is the first barrier against infectious diseases [109]. Clinically, immunomodulators are usually classified into three categories based on their relevant activities including: (1) reducing the stimulation of the immune system or reducing the effectiveness of the immune system (immunosuppressants), (2) promoting the response of the innate immune system (immunostimulants), and (3) enhancing the efficacy of vaccines to stimulate immunity (immunoadjuvants) [109,110]. Many drugs are known to be immunomodulatory substances because they have significant clinical efficacy for altering host responses in the therapy of viral and bacterial infections [111,112,113,114]. Various edible mushrooms have been studied for many years in terms of the effects of their metabolites in boosting immune responses and treating infectious [115,116,117,118]. The principal immunomodulatory effect of active substances derived from mushrooms is to stimulate immune effector cells such as T cells, cytotoxic T lymphocytes (CTL), dendritic cells (DCs), lymphocytes, macrophages and natural killer (NK) cells, resulting in cytokine expression and secretion including interleukins (ILs), tumor necrosis factor-alpha (TNF)-α, and interferon-gamma (INF)-γ [119,120].
Immunomodulators derived from mushrooms are classified into four groups, including lectins, proteins, polysaccharides, and terpenoids [109]. Lectins are carbohydrate-binding proteins that can be found in many organisms and are extracted from mushrooms. They have specific immune cell functions such as antiproliferative, and antitumor activities [108].
Fungal immunomodulatory proteins (FIPs) are small molecular weight proteins, ∼13 kDa and 110–114 amino acids, displaying immunomodulatory activity. They are a type of bioactive substance that can be derived from some edible mushrooms. Meanwhile, mushrooms are an essential source of immunomodulatory polysaccharides, which are long chains of carbohydrate molecules, particularly polymeric carbohydrates, that are composed of monosaccharides linked together by glycosidic bonds [108]. Polysaccharides are responsible for immuno-modulating activities that include stimulating phagocytic activity, acting as inflammatory mediators and in cytokine production [121,122,123]. Terpenes and terpenoids are a large and diverse class of hydrocarbon compounds and typically consist of five-carbon isoprene units [109,124]. Many terpenoids are biologically active and have been widely used for the treatment of many diseases. Simultaneously, they play a diverse role in the fields of cosmetic and food production and have been associated with hormones, medicines, vitamins, etc. [124]. Triterpenoids such as lanostane are the highly oxidized substances that can be isolated from wood-decaying mushrooms, Ganoderma sp. These compounds display immunomodulating and anti-infective effects [125,126,127]. Many species of mushrooms have been found to produce immunomodulators, such as Agaricus bisporus, Agaricus blazei, Amanita pantherina, Boletus satanas, Coprinus cinereus, Cordyceps sinensis, Ga. lucidum, Grifola frondosa, Flammulina velutipes, Ischnoderma resinosum, Lactarius deterrimus, Laetiporus sulphureus, Lentinus tigrinus, Trametes versicolor, and Volvariella volvacea [115,128,129,130], as is detailed in Figure 5 and Table 3.
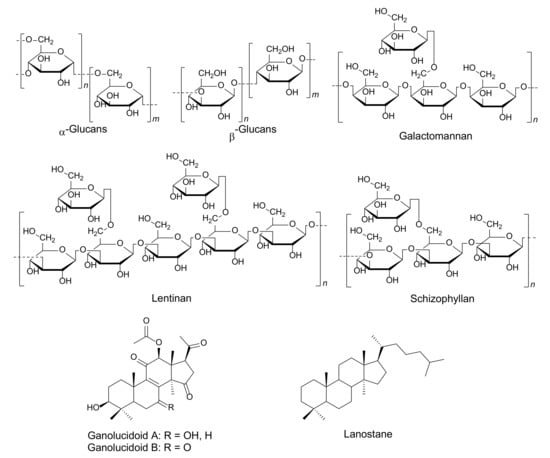
Figure 5.
Fungal bioactive compounds for immunomodulators.
Table 3.
Immunomodulatory activities of mushrooms.
The fungal immunomodulatory protein FIP-fve that was obtained from Flammulina velutipes has been employed to suppress the respiratory syncytial virus (RSV), which is known to cause bronchiolitis. FIP-fve effectively decreased RSV replication, IL-6 expression, and inflammation via inhibition of NF-κB translocation and respiratory pathogenesis in RSV-challenged mice. Interestingly, FIP-fve maymight be seen as a safe substance for viral prevention and disease therapy [133]. Immunomodulators have become useful agents in relieving the pathology that is associated with viral infections going forward [152]. The immunomodulatory mechanisms of mushroom products involve stimulating innate and adaptive immune responses through the activation of macrophages, T lymphocytes, DCs, NK cells, and cytokines. A study of the relationship between the structure and activity of immunomodulators will encourage the development of new therapeutic agents for the treatment of viral infection diseases.
5. Conclusions
The discovery and production of antiviral metabolites from fungi have emerged as part of an exciting field in viral therapeutic and antiviral drug development. Although, CoVs vaccines have been continually developed to alter the occurrence of virally associated diseases, viral protease inhibitors and immunomodulators have become extremely useful agents in this process. The results of the current studies indicate that fungi are an important source of the natural bioactive compounds that have potential as protease inhibitors and immunomodulations. Fungal protease inhibitors reveal strong potential as future candidates in the development of antiviral drugs or alternative and complementary medicals prevention and treatment of CoVs. However, it is of particular interest and concern that fungal protease inhibitors and fungal extracts could have both poisonous and curative effects against CoVs. Presently, there has been a lack of clinical tests that can validate these determinations. Consequently, these circumstances may result in consumers delaying or stopping their pursuit of appropriate medical treatment, which may lead to serious and life-threatening harm to those individuals. Therefore, laboratory assays and clinical tests are needed to fully understand the level of toxicity and pharmacokinetic profile of these viral protease inhibitors and immunomodulators. The important research must be done before the application of these fungal compounds can be used for the prevention and treatment of CoVs in the future, particularly with regard to SARS-CoV-2.
Author Contributions
The project approach was conceptually designed by N.S., J.K., C.S., S.L.; writing and original draft preparation, N.S., J.K., K.S., T.P., C.S.; chemical structure drawing, K.S.; the research was supervised by N.S., S.L.; All authors have read and agreed to the published version of the manuscript.
Funding
This research work was partially supported by Chiang Mai University.
Acknowledgments
We are grateful to Russell K. Hollis for the English proofreading of this manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Cauchemez, S.; Van Kerkhove, M.D.; Riley, S.; Donnelly, C.A.; Fraser, C.; Ferguson, N.M. Transmission scenarios for middle east respiratory syndrome coronavirus (MERS-CoV) and how to tell them apart. Eurosurveillance 2013, 18, 18. [Google Scholar]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Genet. 2018, 17, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. New Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, S.; Teng, T.; Abdalla, A.E.; Zhu, W.; Xie, L.; Wang, Y.; Guo, X. Systematic comparison of two animal-to-human transmitted human coronaviruses: SARS-CoV-2 and SARS-CoV. Viruses 2020, 12, 244. [Google Scholar] [CrossRef]
- World Health Organization. Clinical management of severe acute respiratory infection when novel coronavirus (2019-nCoV) infection is suspected; WHO: Geneva, Switzerland, 2020; Available online: https://www.who.int/ (accessed on 8 March 2020).
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.; Lau, E.H.; Wong, J.Y.; et al. Early transmission dynamics in Wuhan, China, of novel coronavirus–infected pneumonia. New Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- Fernández-Montero, J.V.; Barreiro, P.; Soriano, V. HIV Protease inhibitors: recent clinical trials and recommendations on use. Expert Opin. Pharmacother. 2009, 10, 1615–1629. [Google Scholar] [CrossRef]
- Pokorná, J.; Machala, L.; Řezáčová, P.; Konvalinka, J. Current and novel inhibitors of HIV protease. Viruses 2009, 1, 1209–1239. [Google Scholar] [CrossRef]
- Goetz, D.H.; Choe, Y.; Hansell, E.; Chen, Y.T.; McDowell, M.; Jonsson, C.B.; Roush, W.R.; McKerrow, J.; Craik, C.S. Substrate specificity profiling and identification of a new class of inhibitor for the major protease of the SARS coronavirus. Biochemistry 2007, 46, 8744–8752. [Google Scholar] [CrossRef]
- Anderson, J.; Schiffer, C.; Lee, S.K.; Swanstrom, R. Viral Protease Inhibitors. In Pharmacology and Therapeutics of Cough; Springer Science and Business Media LLC: New York, NY, USA, 2009; Vol. 189, pp. 85–110. [Google Scholar]
- Hosseini, F.S.; Amanlou, M. Simeprevir, Potential candidate to repurpose for coronavirus infection: virtual screening and molecular docking study. Preprints 2020. [Google Scholar]
- Ziebuhr, J.; Gorbalenya, A.E.; Snijder, E.J. Virus-encoded proteinases and proteolytic processing in the Nidovirales. J. Gen. Virol. 2000, 81, 853–879. [Google Scholar] [CrossRef]
- Mielech, A.M.; Kilianski, A.; Baez-Santos, Y.M.; Mesecar, A.D.; Baker, S.C. MERS-CoV papain-like protease has delSGylating and deubiquitinating activities. Virology 2014, 450–451, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, T.P.; Sims, A.C.; Graham, R.L.; Menachery, V.D.; Gralinski, L.E.; Case, J.B.; Leist, S.R.; Pyrc, K.; Feng, J.Y.; Trantcheva, I.; et al. Broad-spectrum antiviral GS-5734 inhibits both epidemic and zoonotic coronaviruses. Sci. Transl. Med. 2017, 9, 3653. [Google Scholar] [CrossRef] [PubMed]
- Dayer, M.R.; Taleb-Gassabi, S.; Dayer, M.S. Lopinavir; A potent drug against coronavirus infection: insight from molecular docking study. Arch. Clin. Infect. Dis. 2017, 12, 13823. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.-J. Potential inhibitors for 2019-nCoV coronavirus M protease from clinically approved medicines. J. Genet. Genom. 2020. [Google Scholar] [CrossRef]
- Lim, J.; Jeon, S.; Shin, H.Y.; Kim, M.J.; Seong, Y.M.; Lee, W.J.; Choe, K.-W.; Kang, Y.M.; Lee, B.; Park, S.J. Case of the index patient who caused tertiary transmission of coronavirus disease 2019 in Korea: the application of lopinavir/ritonavir for the treatment of COVID-19 pneumonia monitored by quantitative RT-PCR. J. Korean Med. Sci. 2020, 35, 79. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Chang, Y.C.; Tung, Y.A.; Lee, K.H.; Chen, T.F.; Hsiao, Y.C.; Chang, H.C.; Hsieh, T.T.; Su, C.H.; Chan-Hung, S.; Su-Shia, W.; et al. Potential therapeutic agents for COVID-19 based on the analysis of protease and RNA polymerase docking. Preprints 2020. [Google Scholar] [CrossRef]
- Contini, A. Virtual screening of an FDA approved drugs database on two COVID-19 coronavirus proteins. ChemRxiv 2020. [Google Scholar] [CrossRef]
- Del Rio, C.; Malani, P.N. COVID-19—New insights on a rapidly changing epidemic. JAMA 2020. [Google Scholar] [CrossRef]
- Mulangu, S.; Dodd, L.E.; Davey, R.T.; Mbaya, O.T.; Proschan, M.; Mukadi, D.; Manzo, M.L.; Nzolo, D.; Oloma, A.T.; Ibanda, A.; et al. A randomized, controlled trial of ebola virus disease therapeutics. New Engl. J. Med. 2019, 381, 2293–2303. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Leist, S.R.; Schäfer, A.; Won, J.; Brown, A.J.; Montgomery, S.A.; Hogg, A.; Babusis, D.; Clarke, M.O.; et al. Comparative therapeutic efficacy of remdesivir and combination lopinavir, ritonavir, and interferon beta against MERS-CoV. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Shi, N.; Shan, F.; Zhang, Z.; Shen, J.; Lu, H.; Ling, Y.; Jiang, Y.; Shi, Y. Emerging 2019 novel coronavirus (2019-nCoV) pneumonia. Radiology 2020, 295, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Collado, J.; Singh, S.; Jayasuriya, H.; Dewey, R.; Polishook, J.D.; Dombrowski, A.W.; Zink, D.L.; Platas, G.; Pelaez, F.; et al. Isolation, structure, and HIV-1-integrase inhibitory activity of structurally diverse fungal metabolites. J. Ind. Microbiol. Biotechnol. 2003, 30, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Berkaew, P.; Intereya, K.; Komwijit, S.; Sathitkunanon, T. Antiplasmodial and antiviral cyclohexadepsipeptides from the endophytic fungus Pullularia sp. BCC 8613. Tetrahedron 2007, 63, 6855–6860. [Google Scholar] [CrossRef]
- Linnakoski, R.; Reshamwala, D.; Veteli, P.; Cortina-Escribano, M.; Vanhanen, H.; Marjomäki, V.S. Antiviral agents from fungi: diversity, mechanisms and potential applications. Front. Microbiol. 2018, 9, 2325. [Google Scholar] [CrossRef]
- Kim, Y.S.; Eo, S.K.; Oh, K.W.; Lee, C.; Han, S.-S. Antiherpetic activities of acidic protein bound polysacchride isolated from Ganoderma lucidum alone and in combinations with interferons. J. Ethnopharmacol. 2000, 72, 451–458. [Google Scholar] [CrossRef]
- Wang, H.X.; Ng, T. Isolation of a novel ubiquitin-like protein from Pleurotus ostreatus mushroom with anti-human immunodeficiency virus, translation-inhibitory, and ribonuclease activities. Biochem. Biophys. Res. Commun. 2000, 276, 587–593. [Google Scholar] [CrossRef]
- Gu, C.Q.; Li, J.W.; Chao, F.; Jin, M.; Wang, X.; Shen, Z.Q. Isolation, identification and function of a novel anti-HSV-1 protein from Grifola frondosa. Antivir. Res. 2007, 75, 250–257. [Google Scholar] [CrossRef]
- Faccin, L.C.; Benati, F.; Rincao, V.P.; Mantovani, M.S.; Soares, S.A.; Gonzaga, M.L.; Nozawa, C.; Carvalho Linhares , R.E. Antiviral activity of aqueous and ethanol extracts and of an isolated polysaccharide from Agaricus brasiliensis against poliovirus type 1. Lett. Appl. Microbiol. 2007, 45, 24–28. [Google Scholar] [CrossRef]
- Zhang, D.; Tao, X.; Chen, R.; Liu, J.; Li, L.; Fang, X.; Yu, L.; Dai, J. Pericoannosin A, a Polyketide synthase–nonribosomal peptide synthetase hybrid metabolite with new carbon skeleton from the endophytic fungus Periconia sp. Org. Lett. 2015, 17, 4304–4307. [Google Scholar] [CrossRef]
- Roy, B.G. Potential of small-molecule fungal metabolites in antiviral chemotherapy. Antivir. Chem. Chemother. 2017, 25, 20–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, X.; Lü, X.; Xu, F.; Wan, J.; Lin, X.; Zhou, X.F.; Liao, S.; Yang, B.; Tu, Z.; et al. Eight new polyketide metabolites from the fungus Pestalotiopsis vaccinii endogenous with the mangrove plant Kandelia candel (L.) Druce. Tetrahedron 2014, 70, 9695–9701. [Google Scholar] [CrossRef]
- Fang, W.; Lin, X.; Zhou, X.; Wan, J.; Lu, X.; Yang, B.; Ai, W.; Zhang, T.; Tu, Z.; Liu, Y. Cytotoxic and antiviral nitrobenzyl sesquiterpenoids from the marine-derived fungus Aspergillus ochraceus Jcma1F17. Med. Chem. Communn. 2014, 5, 701–705. [Google Scholar] [CrossRef]
- Jia, Y.L.; Guan, F.F.; Ma, J.; Wang, C.Y.; Shao, C.L. Pestalotiolide A, a new antiviral phthalide derivative from a soft coral-derived fungus Pestalotiopsis sp. Nat. Prod. Sci. 2015, 21, 227. [Google Scholar] [CrossRef]
- Pang, X.; Lin, X.; Tian, Y.; Liang, R.; Wang, J.; Yang, B.; Zhou, X.F.; Kaliyaperumal, K.; Luo, X.W.; Tu, Z.; et al. Three new polyketides from the marine sponge-derived fungus Trichoderma sp. SCSIO41004. Nat. Prod. Res. 2017, 32, 105–111. [Google Scholar] [CrossRef]
- Zhang, S.P.; Huang, R.; Li, F.F.; Wei, H.X.; Fang, X.W.; Xie, X.S.; Lin, D.G.; Wu, S.; He, J. Antiviral anthraquinones and azaphilones produced by an endophytic fungus Nigrospora sp. from Aconitum carmichaeli. Fitoterapia 2016, 112, 85–89. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, J.; Shen, Y.; Tan, Z.; Zhang, M.; Chen, R.; Zhao, J.; Zhang, D.; Yu, L.; Dai, J. Stachybotrysams A–E, prenylated isoindolinone derivatives with anti-HIV activity from the fungus Stachybotrys chartarum. Phytochem. Lett. 2017, 20, 289–294. [Google Scholar] [CrossRef]
- Sarkar, S.; Koga, J.; Whitley, R.J.; Chatterjee, S. Antiviral effect of the extract of culture medium of Lentinus edodes mycelia on the replication of herpex simplex virus type I. Antiviral Res. 1993, 20, 293–303. [Google Scholar] [CrossRef]
- Razumov, I.A.; Kosogova, T.A.; Kazachinskaia, E.I.; Puchkova, L.I.; Shcherbakova, N.S.; Gorbunova, I.A.; Mikhaĭlovskaia, I.N.; Loktev, V.B.; Tepliakova, T.V. Antiviral activity of aqueous extracts and polysaccharide fractions from mycelium and fruit bodies of higher fungi. Antibiot. chemoterapy 2010, 55, 14–18. [Google Scholar]
- Yamamoto, K.A.; Galhardi, L.C.F.; Rincao, V.P.; Soares, S.; Vieira, I.G.; Ricardo, N.M.P.S.; Nozawa, C.; Linhares, R.E.C. Antiherpetic activity of an Agaricus brasiliensis polysaccharide, its sulfated derivative and fractions. Int. J. Boil. Macromol. 2013, 52, 9–13. [Google Scholar] [CrossRef]
- Puente, X.S.; Sanchez, L.M.; Overall, C.M.; Lopez-Otin, C. Human and mouse proteases: A comparative genomic approach. Nat. Rev. 2003, 4, 544–558. [Google Scholar] [CrossRef] [PubMed]
- Drag, M.; Salvesen, G.S. Emerging principles in protease-based drug discovery. Nat. Rev. Drug Discov. 2010, 9, 690–701. [Google Scholar] [CrossRef]
- Turk, B. Targeting proteases: Successes, failures and future prospects. Nat. Rev. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Brik, A.; Wong, C.H. HIV-1 protease: Mechanism and drug discovery. Org. Biomol. Chem. 2003, 1, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.L.; Morgenstern, K.A.; Lin, C.; Fox, T.; Dwyer, M.D.; Landro, J.A.; Chambers, S.P.; Markland, W.; Lepre, C.A.; O’Malley, E.T.; et al. Crystal structure of the hepatitis C virus NS3 protease domain complexed with a synthetic NS4A cofactor peptide. Cell 1996, 87, 343–355. [Google Scholar] [CrossRef]
- Love, R.A.; Parge, H.E.; Wickersham, J.A.; Hostomsky, Z.; Habuka, N.; Moomaw, E.; Adachi, T.; Hostomska, Z. The crystal structure of hepatitis C virus NS3 proteinase reveals a trypsin-like fold and a structural zinc binding site. Cell 1996, 87, 331–342. [Google Scholar] [CrossRef]
- Urbani, A.; Bianchi, E.; Narjes, F.; Tramontano, A.; de Francesco, R.; Steinkuhler, C.; Pessi, A. Substrate specificity of the hepatitis C virus serine protease NS3. J. Biol. Chem. 1997, 272, 9204–9209. [Google Scholar] [CrossRef]
- Xue, X.; Yu, H.; Yang, H.; Xue, F.; Wu, Z.; Shen, W.; Li, J.; Zhou, Z.; Ding, Y.; Zhao, Q.; et al. Structures of two coronavirus main proteases: implications for substrate binding and antiviral drug design. J. Virol. 2008, 82, 2515–2527. [Google Scholar] [CrossRef]
- Anand, K.; Ziebuhr, J.; Wadhwani, P.; Mesters, J.R.; Hilgenfeld, R. Coronavirus main proteinase (3CLpro) structure: basis for design of anti-SARS drugs. Science 2003, 300, 1763–1767. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S. Peptidomimetic therapeutic agents targeting the protease enzyme of the human immunodeficiency virus and hepatitis C virus. Acc. Chem. Res. 2008, 41, 1252–1263. [Google Scholar] [CrossRef]
- Clercq, E.D.; Li, G. Approved antiviral drugs over the past 50 years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, J.E.; Elowe, N.H.; Huitema, C.; Fortin, P.D.; Cechetto, J.D.; Eltis, L.D.; Brown, E.D. High-throughput screening identifies inhibitors of the SARS coronavirus main proteinase. Chem. Boil. 2004, 11, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Yang, R.; Yoshinaka, Y.; Amari, S.; Nakano, T.; Cinatl, J.; Rabenau, H.; Doerr, H.W.; Hunsmann, G.; Otaka, A.; et al. HIV protease inhibitor nelfinavir inhibits replication of SARS-associated coronavirus. Biochem. Biophys. Res. Commun. 2004, 318, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Booth, C.; Matukas, L.M.; Tomlinson, G.; Rachlis, A.R.; Rose, D.B.; Dwosh, H.A.; Walmsley, S.; Mazzulli, T.; Avendano, M.; Derkach, P.; et al. Clinical features and short-term outcomes of 144 patients with SARS in the greater toronto area. JAMA 2003, 289, 2801. [Google Scholar] [CrossRef] [PubMed]
- Loutfy, M.R.; Blatt, L.M.; Siminovitch, K.A.; Ward, S.; Wolff, B.; Lho, H.; Pham, D.H.; Deif, H.; LaMere, E.A.; Chang, M.; et al. Interferon alfacon-1 plus corticosteroids in severe acute respiratory syndrome. JAMA 2003, 290, 3222. [Google Scholar] [CrossRef]
- Chu, C.M.; Cheng, V.C.C.; Hung, I.F.N.; Wong, M.M.L.; Chan, K.; Kao, R.Y.; Poon, L.L.M.; Wong, C.L.P.; Guan, Y.; Peiris, J.S.M.; et al. Role of lopinavir/ritonavir in the treatment of SARS: initial virological and clinical findings. Thorax 2004, 59, 252–256. [Google Scholar] [CrossRef]
- Stockman, L.J.; Bellamy, R.; Garner, P. SARS: Systematic review of treatment effects. Plos Med. 2006, 3, 343. [Google Scholar] [CrossRef]
- Momattin, H.; Al-Ali, A.Y.; Al-Tawfiq, J.A. A systemtic review of therapeutic agents for the treatment of the Middle East Respiratory Syndrome Coronaavirus (MERS-CoV). Travel. Med. Infect. Dis. 2019, 30, 9–18. [Google Scholar] [CrossRef]
- Stierlé, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef]
- Caruso, M.; Colombo, A.L.; Fedeli, L.; Pavesi, A.; Quaroni, S.; Saracchi, M.; Ventrella, G. Isolation of endophytic fungi and actinomycetes taxane producers. Ann. Microbiol. 2000, 50, 3–13. [Google Scholar]
- Chen, Y.J.; Zhang, Z.; Wang, Y.; Su, Y.; Zhang, R. Screening endophytic fungus to produce taxol from Taxus yunnanensis. Biotechnology 2003, 13, 10–11. [Google Scholar]
- Hoffman, A. Methods for obtaining taxanes. U.S. Patent 6638742, 2003. [Google Scholar]
- Kumaran, R.S.; Muthumary, J.; Hur, B.K. Production of Taxol from Phyllosticta spinarum, an endophytic fungus of Cupressus sp. Eng. Life Sci. 2008, 8, 438–446. [Google Scholar] [CrossRef]
- Kumaran, R.S.; Muthumary, J.; Hur, B.-K. Isolation and identification of an anticancer drug, taxol from Phyllosticta tabernaemontanae, a leaf spot fungus of an angiosperm, Wrightia tinctoria. J. Microbiol. 2009, 47, 40–49. [Google Scholar] [CrossRef]
- Flores-Bustamante, Z.R.; Rivera-Orduña, F.N.; Martínez-Cárdenas, A.; Flores-Cotera, L.B. Microbial paclitaxel: advances and perspectives. J. Antibiot. 2010, 63, 460–467. [Google Scholar] [CrossRef]
- Zhao, K.; Sun, L.X.; Wang, X.; Zhou, D. Screening of high taxol producing fungi by mutagenesis and construction of subtracted cDNA library by Suppression substracted hybridization for differentially expressed genes. Acta. Microbiol. Sin. 2011, 51, 923–933. [Google Scholar]
- Xiong, Z.; Yang, Y.Y.; Zhao, N.; Wang, Y. Diversity of endophytic fungi and screening of fungal paclitaxel producer from Anglojap yew, Taxus x media. Bmc Microbiol. 2013, 13, 71. [Google Scholar] [CrossRef]
- Heinig, U.; Scholz, S.; Jennewein, S. Getting to the bottom of Taxol biosynthesis by fungi. Fungal Divers. 2013, 60, 161–170. [Google Scholar] [CrossRef]
- Naik, B.S. Developments in taxol production through endophytic fungal biotechnology: a review. Orient. Pharm. Exp. Med. 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Priyadarshini, K.; Keerthi, A.U. Paclitaxel Against Cancer: A Short Review. Med. Chem. 2012, 2, 7. [Google Scholar] [CrossRef]
- Tew, W.P. Ovarian cancer in the older woman. J. Geriatr. Oncol. 2016, 7, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Ryang, J.; Yan, Y.; Song, Y.; Liu, F.; Ng, T.B. Anti-HIV, antitumor and immunomodulatory activities of paclitaxel from fermentation broth using molecular imprinting technique. Amb Express 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mary, C.; Sandra, L.; Jamie, V.R.; Michelle, A.R.; Bruce, J.D.; Susan, E.K.; Joseph, A.S. Pilot study evaluating the interaction between paclitaxel and protease inhibitors in patients with human immunodeficiency virus-associated kaposi’s sarcoma: An eastern cooperative oncology group (ECOG) and AIDS malignancy consortium (AMC) trial. Cancer Chemther. Pharmacol. 2011, 68, 827–833. [Google Scholar]
- Casella, T.M.; Eparvier, V.; Mandavid, H.; Bendelac, A.; Odonne, G.; Dayan, L.; Duplais, C.; Espindola, L.S.; Stien, D. Antimicrobial and cytotoxic secondary metabolites from tropical leaf endophytes: Isolation of antibacterial agent pyrrocidine C from Lewia infectoria SNB-GTC2402. Phytochemisty 2013, 96, 370–377. [Google Scholar] [CrossRef]
- Fredenhagen, A.; Petersen, F.; Tintelnot-Blomley, M.; Rosel, J.; Mett, H.; Hug, P. Semicochiodinol A and B: inhibitors of HIV-1 protease and EGF-R protein tyrosine kinase related to asterriquinones produced by the fungus Chrysosporium merdarium. J. Antibiot. 1997, 50, 395–401. [Google Scholar] [CrossRef]
- El-Mekkawy, S.; Meselhy, M.R.; Nakamura, N.; Tezuka, Y.; Hattori, M.; Kakiuchi, N.; Shimotohno, K.; Kawahata, T.; Otake, T. Anti-HIV-1 and anti-HIV-1-protease substances from Ganoderma lucidum. Phytochemistry 1998, 49, 1651–1657. [Google Scholar] [CrossRef]
- Min, B.S.; Nakamura, N.; Miyashiro, H.; Bae, K.W.; Hattori, M. Triterpenes from the spores of Ganoderma lucidum and their inhibitory activity against HIV-1 protease. Chem. Pharm. Bull. 1998, 46, 1607–1612. [Google Scholar] [CrossRef]
- Martinez-Montemayor, M.; Ling, T.; Suárez-Arroyo, I.J.; Ortiz-Soto, G.; Santiago-Negrón, C.L.; Lacourt-Ventura, M.Y.; Valentín-Acevedo, A.; Lang, W.H.; Rivas, F. Identification of biologically Active Ganoderma lucidum compounds and synthesis of improved derivatives That confer anti-cancer activities in vitro. Front. Pharmacol. 2019, 10, 115. [Google Scholar] [CrossRef]
- El Dine, R.S.; Halawany, A.M.E.; Ma, C.M.; Hattori, M. Anti-HIV1- protease activity of lanostane triterpenes from the Vienamese mushroom Ganoderma colossum. J. Nat. Prod. 2008, 71, 1022–1026. [Google Scholar] [CrossRef]
- El Dine, R.S.; El-Halawany, A.; Ma, C.M.; Hattori, M. Inhibition of the dimerization and active site of HIV-1 protease by secondary metabolites from the Vietnamese Mushroom Ganoderma colossum. J. Nat. Prod. 2009, 72, 2019–2023. [Google Scholar] [CrossRef]
- Sato, N.; Zhang, Q.; Ma, C.-M.; Hattori, M. Anti-human immunodeficiency virus-1 protease activity of new lanostane-type triterpenoids from Ganoderma sinense. Chem. Pharm. Bull. 2009, 57, 1076–1080. [Google Scholar] [CrossRef] [PubMed]
- Sillapachaiyaporn, C.; Chuchawankul, S. HIV-1 protease and reverse transcriptase inhibition by tiger milk mushroom (Lignosus rhinocerus) sclerotium extracts: In vitro and in silico studies. J. Tradit. Complement. Med. 2019. [CrossRef]
- Sillapachaiyaporn, C.; Nilkhet, S.; Ung, A.T.; Chuchawankul, S. Anti-HIV-1 protease activity of the crude extracts and isolated compounds from Auricularia polytricha. Bmc Complement. Altern. Med. 2019, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Ng, T. A peptide with HIV-1 reverse transcriptase inhibitory activity from the medicinal mushroom Russula paludosa. Peptides 2007, 28, 560–565. [Google Scholar] [CrossRef]
- Jiang, Y.; Wong, J.; Fu, M.; Ng, T.B.; Liu, Z.; Wang, C.; Li, N.; Qiao, W.; Wen, T.; Liu, F. Isolation of adenosine, iso-sinensetin and dimethylguanosine with antioxidant and HIV-1 protease inhibiting activities from fruiting bodies of Cordyceps militaris. Phytomedicine 2011, 18, 189–193. [Google Scholar] [CrossRef]
- Gallego, P.; Rojas, A.; Falcón, G.; Carbonero, P.; García-Lozano, M.R.; Gil, A.; Grande, L.; Cremades, O.; Romero-Gómez, M.; Bautista, J.D.; et al. Water-soluble extracts from edible mushrooms (Agaricus bisporus) as inhibitors of hepatitis C viral replication. Food Funct. 2019, 10, 3758–3767. [Google Scholar] [CrossRef]
- Hawas, U.W.; El Desouky, S.; El Kassem, L.A.; Elkhateeb, W. Alternariol derivatives from Alternaria alternata, an endophytic fungi residing in red sea soft coral, inhibit HCV NS3/4A protease. Appl. Biochem. Microbiol. 2015, 51, 579–584. [Google Scholar] [CrossRef]
- Schmutz, C.; Cenk, E.; Marko, D. The Alternaria mycotoxin alternariol triggers the immune response of IL-1β-stimulated, differentiated Caco-2 cells. Mol. Nutr. Food Res. 2019, 63, 1900341. [Google Scholar] [CrossRef]
- Grover, S.; Lawrence, C.B. The Alternaria alternata mycotoxin Alternariol suppresses lipopolysaccharide-induced inflammation. Int. J. Mol. Sci. 2017, 18, 1577. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, M.H.; Wang, X.B.; Li, T.X.; Kong, L. Bioactive metabolites from the endophytic fungus Alternaria alternata. Fitoterapia 2014, 99, 153–158. [Google Scholar] [CrossRef]
- Hawas, U.W.; Al Farawati, R.; El Kassem, L.T.A.; Turki, A.J. Different culture metabolites of the red sea fungus Fusarium equiseti optimize the inhibition of hepatitis C virus NS3/4A protease (HIV PR). Mar. Drugs 2016, 14, 190. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.Y.; Lin, J.; Zhou, B.; Liu, Y.; Zhu, B.Q. H1-A, a compound isolated from Fusarium oxysporum inhibits hepatitis C virus (HCV) NS3 serine protease. Chin. J. Nat. Med. 2016, 14, 299–302. [Google Scholar] [CrossRef]
- Chu, M.; Mierzwa, R.; He, L.; King, A.; Patel, M.; Pichardo, J.; Hart, A.; Butkiewicz, N.; Puar, M.S. Isolation and structure (HCV) NS3 protease inhibitor from the fungus Penicillum griseofulvum. Bioorg. Med. Chem. Lett. 1999, 9, 1949–1952. [Google Scholar] [CrossRef]
- Dai, J.R.; Carte, B.K.; Sidebottom, P.J.; Yew, A.L.S.; Ng, S.B.; Huang, Y.; Butler, M.S. Circumdatin G, a new alkaloid from the fungus Aspergillus ochraceus. J. Nat. Prod. 2001, 64, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Phuong, D.T.; Ma, C.M.; Hattori, M.; Jin, J.S. Inhibitory effects of antrodins A-E from Antrodia cinnamomea and their metabolites on hepatitis C virus protease. Phytother. Res. 2009, 23, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Hawas, U.W.; El Kassem, L.T.A.; Ahmed, E.F.; Emam, M. In-vitro bioassays on the metabolites of the fungus Emericella nidulans isolated from the Egyptian red sea algae. Egypt. Pharmaceut. J. 2012, 11, 124–128. [Google Scholar]
- Lee, D.; Lee, W.Y.; Jung, K.; Kwon, Y.S.; Kim, D.; Hwang, G.S.; Kim, C.E.; Lee, S.; Kang, K.S. The inhibitory effect of cordycepin on the proliferation of MCF-7 breast cancer cells, and its mechanism: An investigation using network pharmacology-based analysis. Biomolecules 2019, 9, 414. [Google Scholar] [CrossRef]
- Xu, J.C.; Zhou, X.P.; Wang, X.A.; Xu, M.D.; Chen, T.; Chen, T.Y.; Zhou, P.H.; Zhang, Y.Q. Cordycepin induces apoptosis and G2/M phase arrest through the ERK pathways in esophageal cancer cells. J. Cancer 2019, 10, 2415. [Google Scholar] [CrossRef]
- Wu, H.Y.; Yang, F.L.; Li, L.H.; Rao, Y.K.; Ju, T.C.; Wong, W.T.; Hsieh, C.Y.; Pivkin, M.V.; Hua, K.F.; Wu, S.H. Ergosterol peroxide from marine fungus Phoma sp. induces ROS-dependent apoptosis and autophagy in human lung adenocarcinoma cells. Sci. Rep. 2018, 8, 17956. [Google Scholar] [CrossRef]
- Harada, H.; Yamashita, U.; Kurihara, H.; Fukushi, E.; Kawabata, J.; Kamei, Y. Antitumer activity of palmitic acid found as a selective cytotoxic substance in a marine red alga. Anticancer Res. 2002, 22, 2587–2590. [Google Scholar]
- Ahmed, E.F.; Rateb, M.E.; El Kassem, L.T.A.; Hawas, U.W. Anti-HCV protease of diketopiperazines produced by the red sea sponge-saaociated fungus Aspergillus versicolor. Appl. Biochem. Microbiol. 2017, 53, 101–106. [Google Scholar] [CrossRef]
- Hawas, U.; El-Halawany, A.; Ahmede, E.F. Hepatitis C Virus NS3-NS4A protease inhibitors from the endophytic Penicillium chrysogenum isolated from the red alga Liagora viscida. Z. Für Nat. C 2013, 68, 355–366. [Google Scholar] [CrossRef]
- Zhang, K.; Hou, Q.; Zhong, Z.; Li, X.; Chen, H.; Li, W.; Wen, J.; Wang, L.; Liu, W.; Zhong, F. Porcine reproductive and respiratory syndrome virus activates inflammasomes of porcine alveolar macrophages via its small envelope protein E. Virology 2013, 442, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Peiris, J.M.; Lai, S.; Poon, L.L.; Guan, Y.; Yam, L.; Lim, W.; Nicholls, J.M.; Yee, W.; Yan, W.; Cheung, M.; et al. Coronavirus as a possible cause of severe acute respiratory syndrome. Lancet 2003, 361, 1319–1325. [Google Scholar] [CrossRef]
- Channappanavar, R.; Perlman, S. Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. Semin. Immunopathol. 2017, 39, 529–539. [Google Scholar] [CrossRef]
- Enshasy, H.; Hatti-Kaul, R. Mushroom immunomodulators: unique molecules with unlimited applications. Trends Biotechnol. 2013, 31, 668–677. [Google Scholar] [CrossRef]
- Kumar, D.; Arya, V.; Kaur, R.; Bhat, Z.A.; Gupta, V.K.; Kumar, V. A review of immunomodulators in the Indian traditional health care system. J. Microbiol. Immunol. Infect. 2012, 45, 165–184. [Google Scholar] [CrossRef]
- González-Navajas, J.M.; Lee, J.; David, M.; Raz, E. Immunomodulatory functions of type I interferons. Nat. Rev. Immunol. 2012, 12, 125–135. [Google Scholar] [CrossRef]
- Kak, V.; Sundareshan, V.; Modi, J.; Khardori, N.M. Immunotherapies in infectious diseases. Med. Clin. North. Am. 2012, 96, 455–474. [Google Scholar] [CrossRef]
- Labro, M.T. Immunomodulatory effects of antimicrobial agents. Part I: antibacterial and antiviral agents. Expert Rev. Anti-Infect. Ther. 2012, 10, 319–340. [Google Scholar] [CrossRef] [PubMed]
- Zapater, P.; González-Navajas, J.M.; Such, J.; Francés, R. Immunomodulating effects of antibiotics used in the prophylaxis of bacterial infections in advanced cirrhosis. World J. Gastroenterol. 2015, 21, 11493–11501. [Google Scholar] [CrossRef] [PubMed]
- Guggenheim, A.G.; Wright, K.M.; Zwickey, H.L. Immune modulation from five major Mushrooms: application to integrative oncology. Integr. Med. (Encinitas, Calif.) 2014, 13, 32–44. [Google Scholar]
- Li, Q.; Wang, X.F.; Zhou, X.W. Recent status and prospects of the fungal immunomodulatory protein family. Crit. Rev. Biotechnol. 2011, 31, 365–375. [Google Scholar] [CrossRef]
- Mallard, B.; Leach, D.N.; Wohlmuth, H.; Tiralongo, J. Synergistic immuno-modulatory activity in human macrophages of a medicinal mushroom formulation consisting of Reishi, Shiitake and Maitake. Plos One 2019, 14, e0224740. [Google Scholar] [CrossRef]
- Shao, K.D.; Mao, P.W.; Li, Q.Z.; Li, L.D.J.; Wang, Y.L.; Zhou, X.W. Characterization of a novel fungal immunomodulatory protein, FIP-SJ75 shuffled from Ganoderma lucidum, Flammulina velutipes and Volvariella volvacea. Food Agric. Immunol. 2019, 30, 1253–1270. [Google Scholar] [CrossRef]
- Lull, C.; Wichers, H.J.; Savelkoul, H. Antiinflammatory and immunomodulating properties of fungal metabolites. Mediat. Inflamm. 2005, 2005, 63–80. [Google Scholar] [CrossRef]
- Moradali, M.F.; Mostafavi, H.; Ghods, S.; Hedjaroude, G.A. Immunomodulating and anticancer agents in the realm of macromycetes fungi (macrofungi). Int. Immunopharmacol. 2007, 7, 701–724. [Google Scholar] [CrossRef]
- Brown, G.D.; Herre, J.; Williams, D.L.; Willment, J.A.; Marshall, A.; Gordon, S. Dectin-1 mediates the biological effects of β-glucans. J. Exp. Med. 2003, 197, 1119–1124. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vashishta, A.; Saraswat-Ohri, S.; Vetvickova, J. Immunological effects of yeast- and mushroom-derived β-glucans. J. Med. Food. 2008, 11, 615–622. [Google Scholar] [CrossRef]
- Wasser, S. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [PubMed]
- Perveen, S. Introductory Chapter: Terpenes and Terpenoids. In Terpenes and Terpenoids; InTech Open: London, UK, 2018; pp. 1–12. [Google Scholar] [CrossRef]
- Jeong, Y.T.; Yang, B.K.; Jeong, S.C.; Kim, S.M.; Song, C.H. Ganoderma applanatum: a promising mushroom for antitumor and immunomodulating activity. Phytother. Res. 2008, 22, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Ren, W.; Zhou, Y.; Ma, J.; Ruan, Y.; Wen, C.N. Triterpenoids from the spores of Ganoderma lucidum. North. Am. J. Med. Sci. 2011, 3, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Su, H.G.; Peng, X.R.; Shi, Q.Q.; Huang, Y.J.; Zhou, L.; Qiu, M.H. Lanostane triterpenoids with anti-inflammatory activities from Ganoderma lucidum. Phytochemistry 2020, 173, 112256. [Google Scholar] [CrossRef]
- Li, Q.Z.; Zheng, Y.Z.; Zhou, X.W. Fungal immunomodulatory proteins: characteristic, potential antitumor activities and their molecular mechanisms. Drug Discov. Today 2019, 24, 307–314. [Google Scholar] [CrossRef]
- Sonawane, H.; Bhosle, H.; Bapat, G.; Vikram, G. Pharmaceutical metabolites with potent bioactivity from mushrooms. J. Phar. Res. 2014, 8, 969–972. [Google Scholar]
- Sze, S.; Ho, J.; Liu, W. Volvariella volvacea lectin activates mouse T lymphocytes by a calcium dependent pathway. J. Cell. Biochem. 2004, 92, 1193–1202. [Google Scholar] [CrossRef]
- Švajger, U.; Pohleven, J.; Kos, J.; Strukelj, B.; Jeras, M. CNL, a ricin B-like lectin from mushroom Clitocybe nebularis, induces maturation and activation of dendritic cells via the toll-like receptor 4 pathway. Immunol. 2011, 134, 409–418. [Google Scholar] [CrossRef]
- Wang, H.X.; Ng, T.B.; Ooi, V.E.; Liu, W.K.; Chang, S.T. Action of lectin from the mushroom Trichoderma mongolicum on macrophages, splenocytes and life-span in sarcoma-bearing mice. Anticancer Res. 1997, 17, 419–429. [Google Scholar]
- Chang, Y.C.; Chow, Y.H.; Sun, H.; Liu, Y.F.; Lee, Y.T.; Lue, K.H.; Ko, J.L. Alleviation of respiratory syncytial virus replication and inflammation by fungal immunomodulatory protein FIP-fve from Flammulina velutipes. Antivir. Res. 2014, 110, 124–131. [Google Scholar] [CrossRef]
- Paaventhan, P.; Joseph, J.S.; Seow, S.V.; Vaday, S.; Robinson, H.; Chua, K.Y.; Kolatkar, P. A 1.7A structure of Fve, a member of the new fungal immunomodulatory protein family. J. Mol. Boil. 2003, 332, 461–470. [Google Scholar] [CrossRef]
- Xu, H.; Kong, Y.Y.; Chen, X.; Guo, M.; Bai, X.H.; Lu, Y.J.; Li, W.; Zhou, X.W. Recombinant FIP-gat, a fungal immunomodulatory protein from Ganoderma atrum, induces growth inhibition and cell death in breast cancer cells. J. Agric. Food Chem. 2016, 64, 2690–2698. [Google Scholar] [CrossRef] [PubMed]
- Li, J.R.; Cheng, C.L.; Yang, W.J.; Yang, C.R.; Ou, Y.C.; Wu, M.J.; Ko, J.L. FIP-gts potentiate autophagic cell death against cisplatin-resistant urothelial cancer cells. Anticancer. Res. 2014, 34, 2973–2983. [Google Scholar] [PubMed]
- Gao, Y.; Wáng, Y.; Wu, Y.; Chen, H.; Yang, R.; Bao, D. Protective function of novel fungal immunomodulatory proteins Fip-lti1 and Fip-lti2 from Lentinus tigrinus in concanavalin A induced liver oxidative injury. Oxidative Med. Cell. Longev. 2019, 2019, 1–15. [Google Scholar] [CrossRef]
- Li, S.Y.; Shi, L.J.; Ding, Y.; Nie, Y.; Tang, X. Identification and functional characterization of a novel fungal immunomodulatory protein from Postia placenta. Food Chem. Toxicol. 2015, 78, 64–70. [Google Scholar] [CrossRef]
- Hsu, H.-C.; Hsu, C.I.; Lin, R.H.; Kao, C.L.; Lin, J.Y. Fip-vvo, a new fungal immunomodulatory protein isolated from Volvariella volvacea. Biochem. J. 1997, 323, 557–565. [Google Scholar] [CrossRef]
- Hsin, I.L.; Ou, C.C.; Wu, M.F.; Jan, M.S.; Hsiao, Y.M.; Lin, C.H.; Ko, J.L. GMI, an immunomodulatory protein from Ganoderma microsporum, potentiates cisplatin-induced apoptosis via autophagy in lung cancer cells. Mol. Pharm. 2015, 12, 1534–1543. [Google Scholar] [CrossRef]
- Haak-Frendscho, M.; Kino, K.; Sone, T.; Jardieu, P. Ling Zhi-8: A novel T cell mitogen induces cytokine production and upregulation of ICAM-1 expression. Cell. Immunol. 1993, 150, 101–113. [Google Scholar] [CrossRef]
- Wang, S.Y.; Hsu, M.L.; Hsu, H.C.; Tzeng, C.H.; Lee, S.S.; Shiao, M.S.; Ho, C.K. The anti-tumor effect of Ganoderma lucidum is mediated by cytokines released from activated macrophages and T lymphocytes. Int. J. Cancer. 1997, 70, 699–705. [Google Scholar] [CrossRef]
- Yeh, C.H.; Chen, H.C.; Yang, J.J.; Chuang, W.I.; Sheu, F. Polysaccharides PS-G and protein LZ-8 from Reishi (Ganoderma lucidum) exhibit diverse functions in regulating murine aacrophages and T lymphocytes. Agric. Food Chem. 2010, 58, 8535–8544. [Google Scholar] [CrossRef]
- Kozarski, M.; Klaus, A.; Nikšić, M.; JakovljeviĆ, D.; Helsper, J.P.; Van Griensven, L.J.L.D. Antioxidative and immunomodulating activities of polysaccharide extracts of the medicinal mushrooms Agaricus bisporus, Agaricus brasiliensis, Ganoderma lucidum and Phellinus linteus. Food Chem. 2011, 129, 1667–1675. [Google Scholar] [CrossRef]
- Adachi, Y.; Okazaki, M.; Ohno, N.; Yadomae, T. Enhancement of cytokine production by macrophages stimulated with (l→3)-β-D-glucan, grifolan (GRN), isolated from Grifola frondosa. Biologic. Pharma. Bul. 1994, 17, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.R.; Patel, D.K.; Shin, W.C.; Sim, W.S.; Lee, O.H.; Lim, K.T. Structural elucidation and immune-enhancing effects of novel polysaccharide from Grifola frondosa. Biomed. Res. Int. 2019, 2019, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.L.; Chen, Y.; Wang, S.S.; Kai, G.Q.; Fang, Y.M. Isolation, partial characterisation and immunomodulatory activities of polysaccharide from Morchella esculenta. J. Sci. Food Agric. 2011, 91, 2180–2185. [Google Scholar] [CrossRef] [PubMed]
- Su, C.A.; Xu, X.Y.; Liu, D.Y.; Wu, M.; Zeng, F.Q.; Zeng, M.Y.; Wei, W.; Jiang, N.; Luo, X. Isolation and characterization of exopolysaccharide with immunomodulatory activity from fermentation broth of Morchella conica. Daru J. Pharm. Sci. 2013, 21, 5. [Google Scholar] [CrossRef]
- Murata, Y.; Shimamura, T.; Tagami, T.; Takatsuki, F.; Hamuro, J. The skewing to Th1 induced by lentinan is directed through the distinctive cytokine production by macrophages with elevated intracellular glutathione content. Int. Immunopharmacol. 2002, 2, 673–689. [Google Scholar] [CrossRef]
- Hobbs, C.R. The chemistry, nutritional value, immunopharmacology, and safety of the traditional food of medicinal split-gill fungus Schizophyllum commune Fr.:Fr. (Schizophyllaceae). A Literature Review. Int. J. Med. Mushrooms 2005, 7, 127–140. [Google Scholar] [CrossRef]
- Kim, K.H.; Moon, E.; Choi, S.U.; Kim, S.Y.; Lee, K.R. Lanostane triterpenoids from the mushroom Naematoloma fasciculare. J. Nat. Prod. 2013, 76, 845–851. [Google Scholar] [CrossRef]
- Malemud, C.J. Immunomodulators in autoimmunity and viral infections. J. Clin. Cell. Immunol. 2018, 9, 1–4. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).