Melatonin Promotes the Proliferation of Chicken Sertoli Cells by Activating the ERK/Inhibin Alpha Subunit Signaling Pathway

Abstract
1. Introduction
2. Results
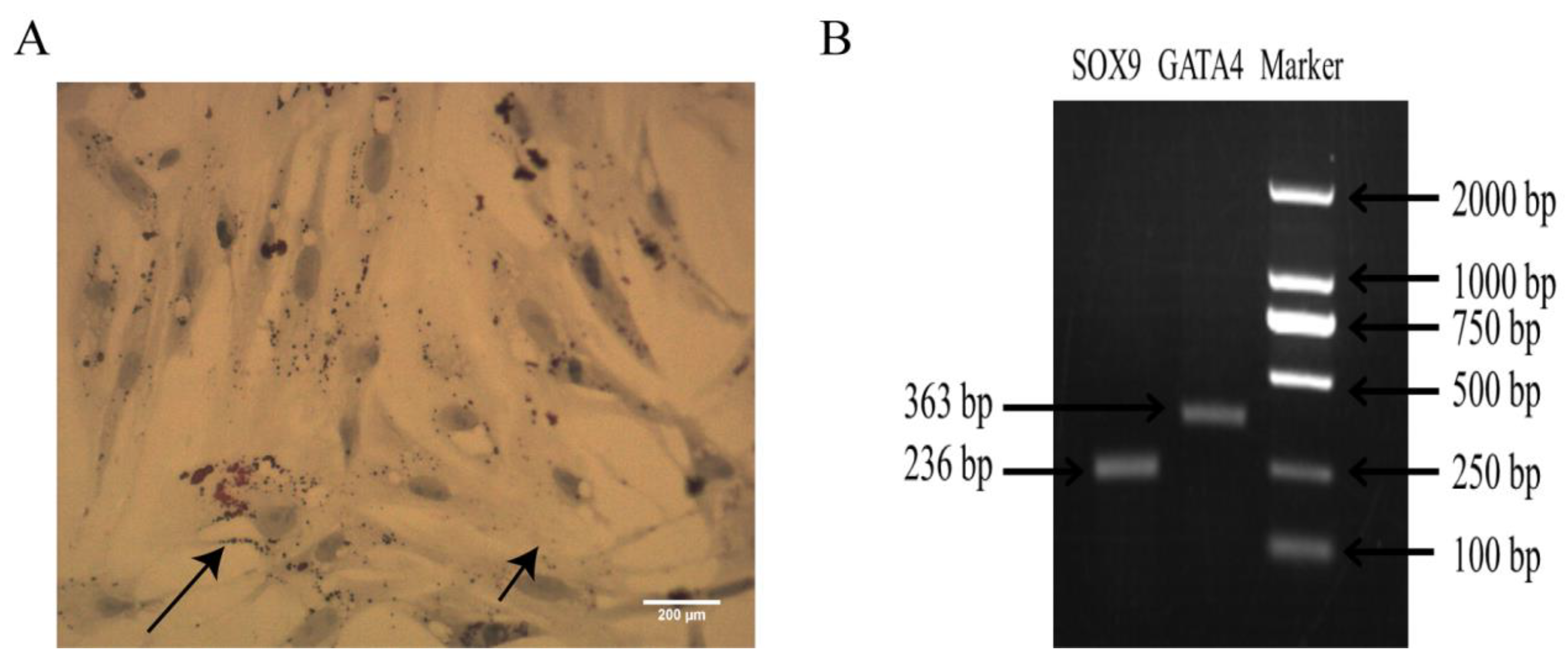
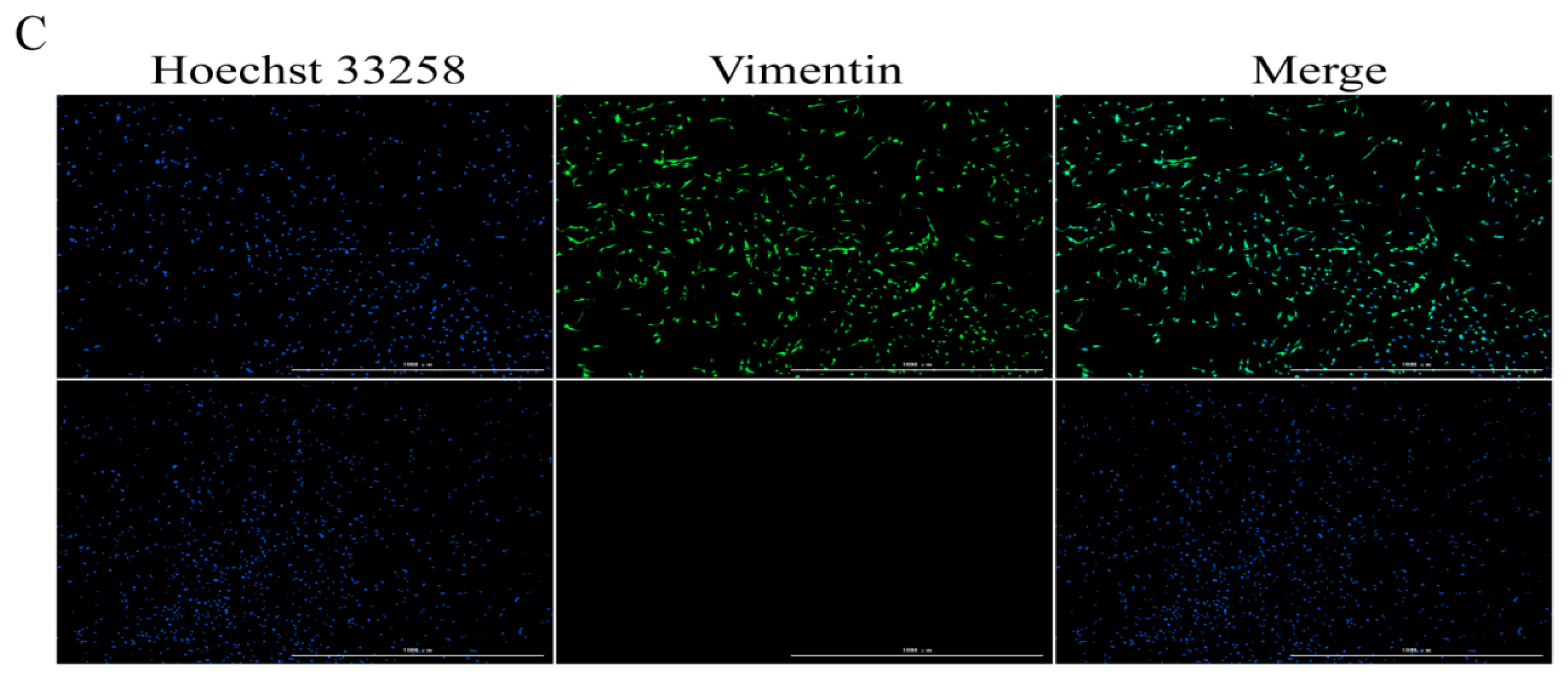
2.1. Identification of Sertoli Cells
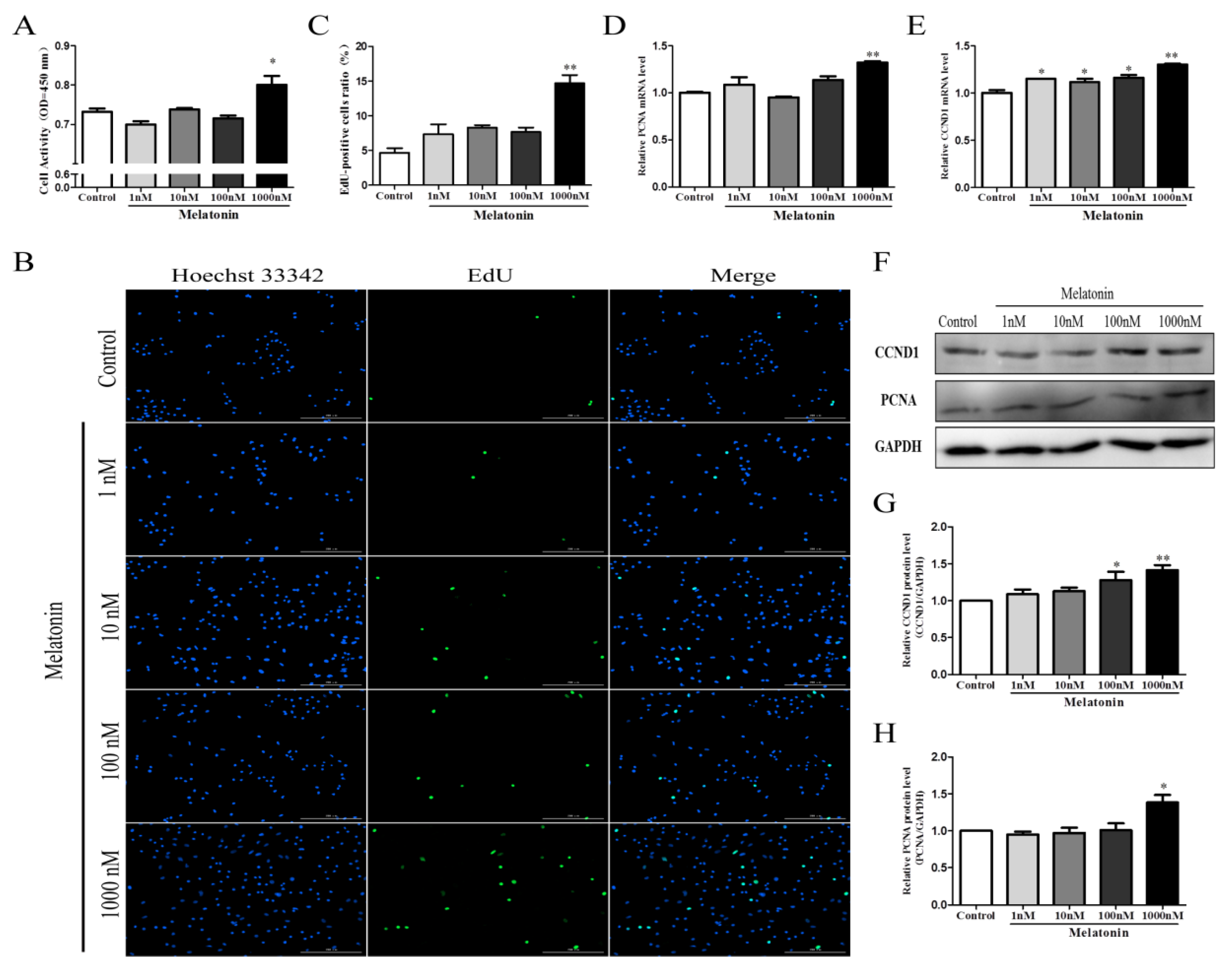
2.2. Melatonin Promoted the Proliferation of Chicken Sertoli Cells
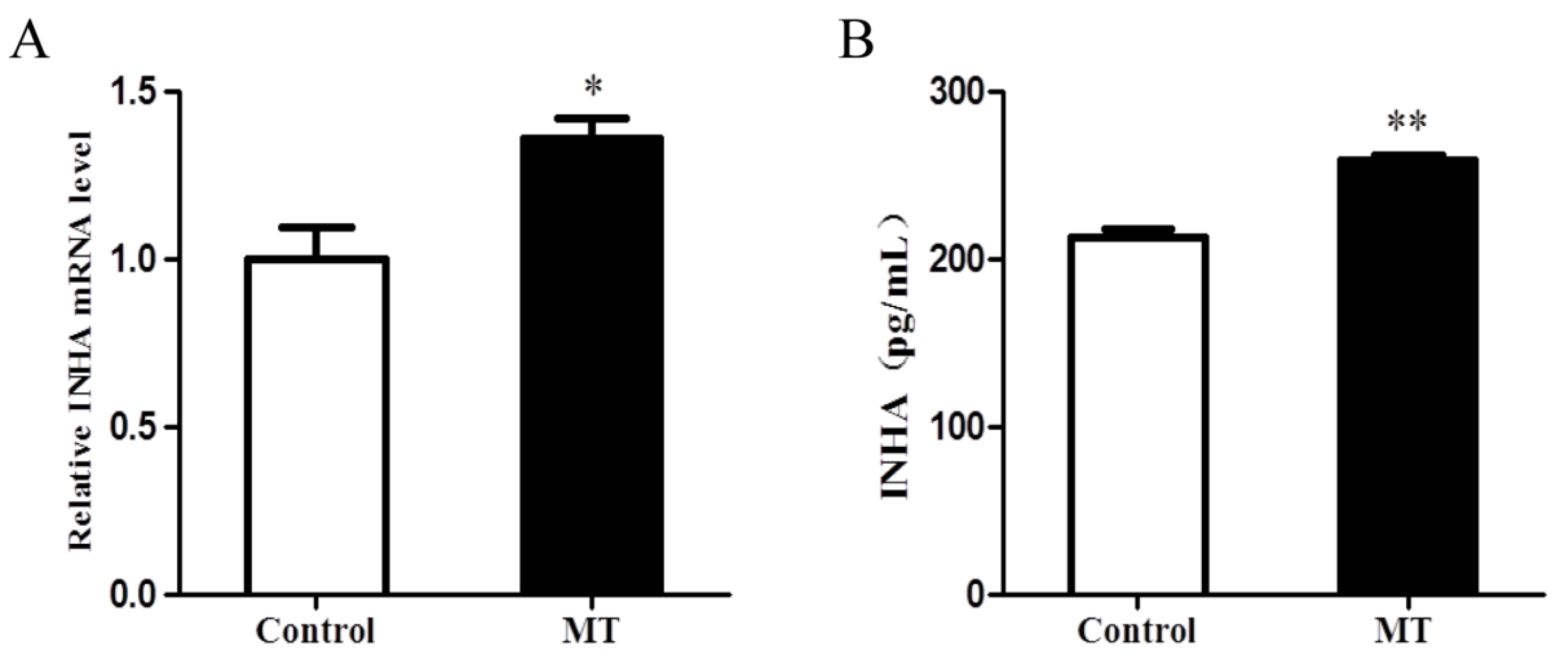
2.3. Melatonin Promoted the Expression of INHA in Chicken Sertoli Cells
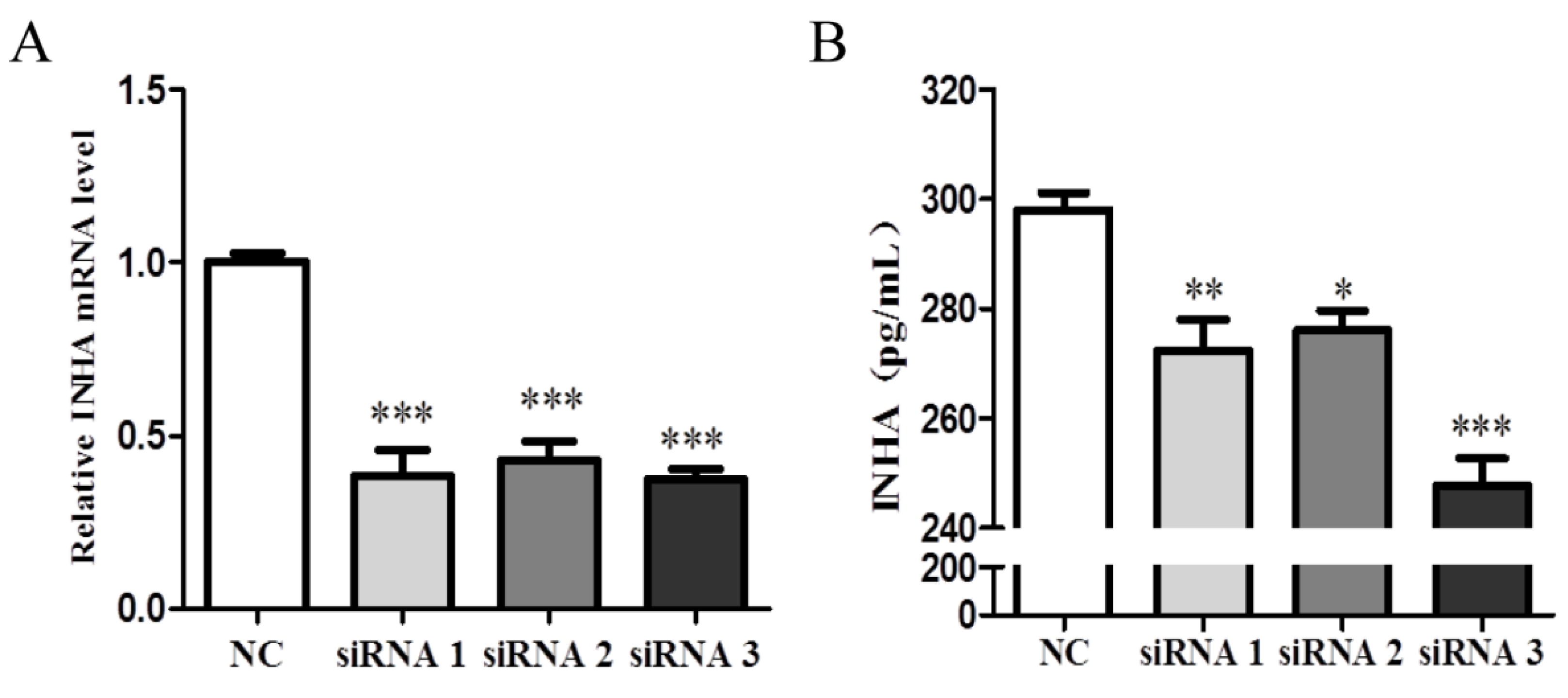
2.4. Identification of the Interference Efficiency of INHA siRNA
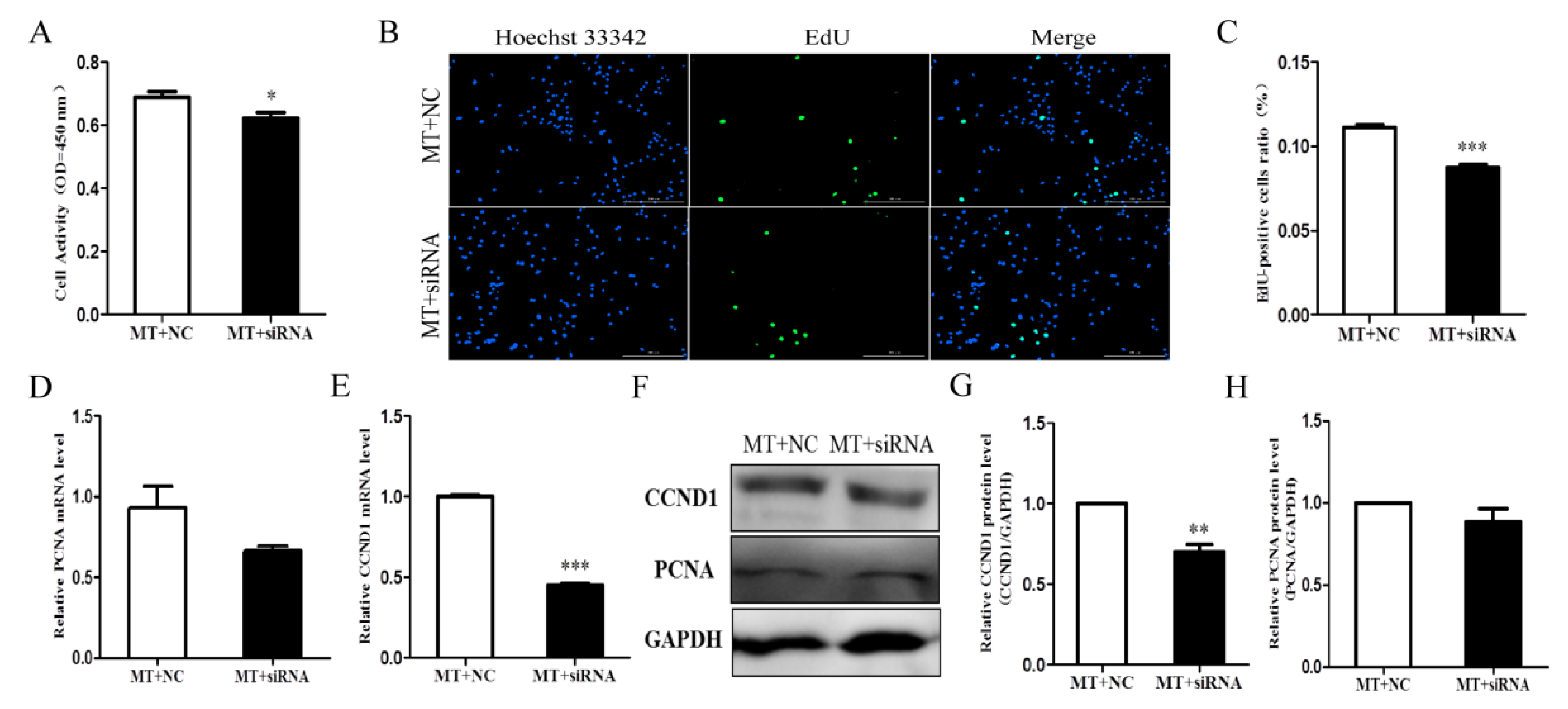
2.5. Melatonin Promoted Cell Proliferation by Affecting INHA in Chicken Sertoli Cells
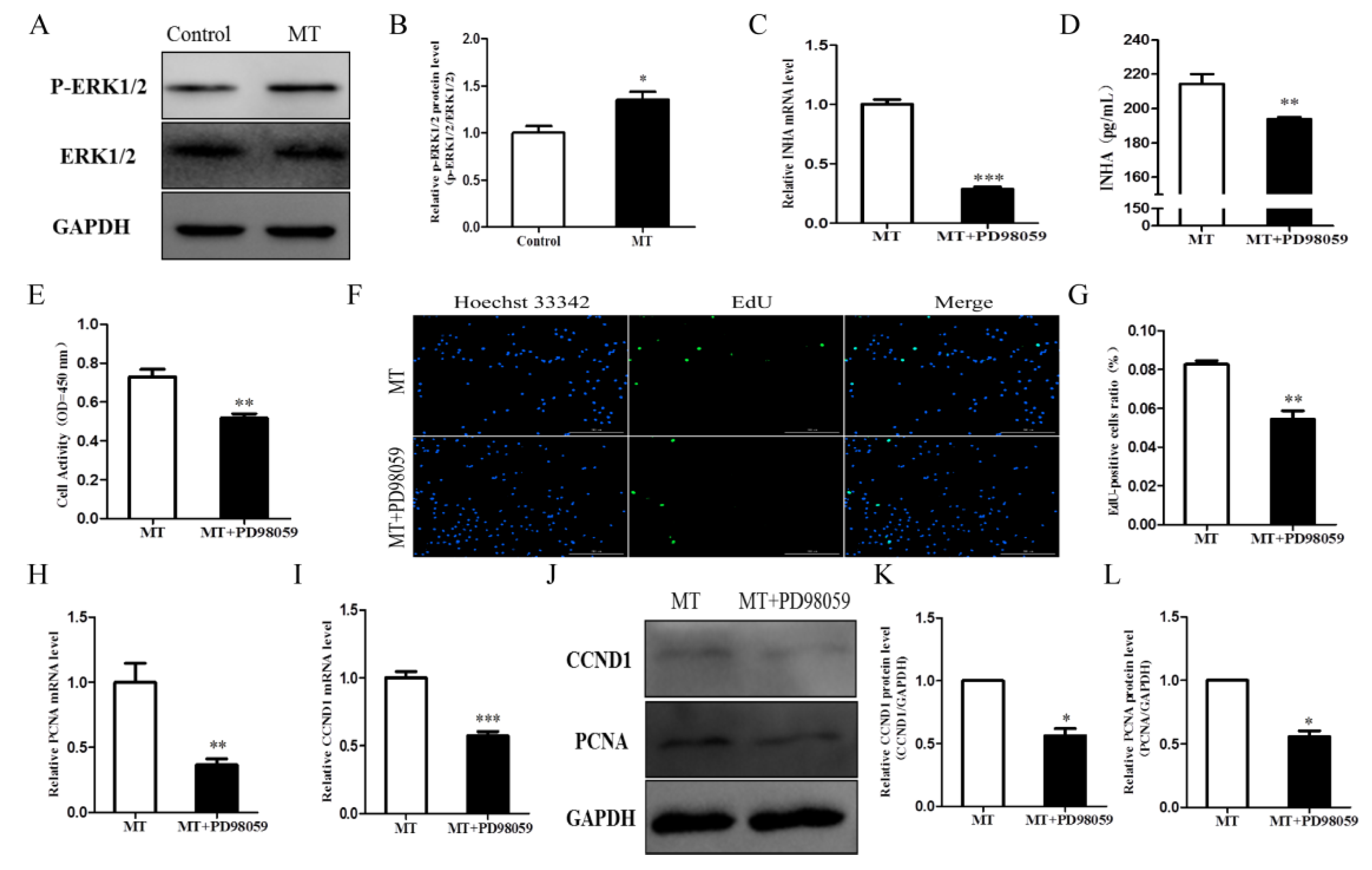
2.6. Melatonin Promotes Cell Proliferation by Activating the ERK Signaling Pathway and Affecting INHA in Chicken Sertoli Cells
3. Discussion
4. Materials and Methods
4.1. Primary Sertoli Cell Culture
4.2. Sertoli Cell Treatment
4.3. Oil Red O Staining
4.4. PCR and Agarose Gel Electrophoresis
4.5. Immunofluorescence Assay
4.6. CCK-8 Assay
4.7. Cell Transfection
4.8. Total RNA Isolation and Real-Time qPCR
4.9. EdU Staining
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Western Blotting Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rato, L.; Alves, M.G.; Socorro, S.; Duarte, A.I.; Cavaco, J.E.; Oliveira, P.F. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.R.; Liu, Y.X. Regulation of spermatogonial stem cell self-renewal and spermatocyte meiosis by Sertoli cell signaling. Reproduction 2015, 149, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Hai, Y.; Hou, J.; Liu, Y.; Liu, Y.; Yang, H.; Li, Z.; He, Z. The roles and regulation of Sertoli cells in fate determinations of spermatogonial stem cells and spermatogenesis. Semin. Cell Dev. Biol. 2014, 29, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Vadala, S.; Dufour, J.M. An overview of a Sertoli cell transplantation model to study testis morphogenesis and the role of the Sertoli cells in immune privilege. Environ. Epigenet 2017, 3, dvx012. [Google Scholar] [CrossRef]
- Jackson, G.L.; Kuehl, D.E. Interactions of photoperiod, testosterone, and naloxone on GnRH and LH pulse parameters in the male sheep. Domest. Anim. Endocrinol. 2000, 18, 97–110. [Google Scholar] [CrossRef]
- Dawson, D.; Encel, N. Melatonin and sleep in humans. J. Pineal Res. 1993, 15, 1–12. [Google Scholar] [CrossRef]
- Lewis, P.D.; Middleton, B.A.; Gous, R.M. Exogenous melatonin modifies rate of sexual maturation in domestic pullets. Poult. Sci. 2006, 85, 117–122. [Google Scholar] [CrossRef]
- Sainz, R.M.; Mayo, J.C.; Rodriguez, C.; Tan, D.X.; Lopez-Burillo, S.; Reiter, R.J. Melatonin and cell death: Differential actions on apoptosis in normal and cancer cells. Cell. Mol. Life Sci. Cmls 2003, 60, 1407–1426. [Google Scholar] [CrossRef]
- Semercioz, A.; Onur, R.; Ogras, S.; Orhan, I. Effects of melatonin on testicular tissue oxide level and antioxidant enzyme activities in experimentally induced left varicocele. Neuro Endocrinol. Lett. 2003, 24, 86–90. [Google Scholar]
- Sarabia, L.; Maurer, I.; Bustos-Obregon, E. Melatonin prevents damage elicited by the organophosphorous pesticide diazinon on mouse sperm DNA. Ecotoxicol. Environ. Saf. 2009, 72, 663–668. [Google Scholar] [CrossRef]
- Rossi, S.P.; Windschuettl, S.; Matzkin, M.E.; Terradas, C.; Ponzio, R.; Puigdomenech, E.; Levalle, O.; Calandra, R.S.; Mayerhofer, A.; Frungieri, M.B. Melatonin in testes of infertile men: Evidence for anti-proliferative and anti-oxidant effects on local macrophage and mast cell populations. Andrology 2014, 2, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Jin, L.; Chen, M.; Zheng, Z.; Lu, W.; Fan, W.; Li, L.; Zheng, F.; Zhu, Q.; Qiu, H.; et al. Ku80 promotes melanoma growth and regulates antitumor effect of melatonin by targeting HIF1-alpha dependent PDK-1 signaling pathway. Redox Biol. 2019, 25, 101197. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.L.; Wang, Z.P.; Jin, C.; Kang, X.L.; Liu, Y.-X. Melatonin promotes sheep Leydig cell testosterone secretion in a co-culture with Sertoli cells. Theriogenology 2017, 106, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Deng, S.L.; Sun, T.C.; Li, Y.Y.; Liu, Y.X. Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules 2018, 23, 447. [Google Scholar] [CrossRef]
- Xiong, J.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Effect of the melatonin nuclear receptor RORalpha on monochromatic light-induced T-lymphocyte proliferation in chicken thymus. Immunol. Lett. 2019, 213, 21–29. [Google Scholar] [CrossRef]
- Yang, W.C.; Tang, K.Q.; Fu, C.Z.; Riaz, H.; Zhang, Q.; Zan, L.S. Melatonin regulates the development and function of bovine Sertoli cells via its receptors MT1 and MT2. Anim. Reprod. Sci. 2014, 147, 10–16. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Melatonin receptor subtypes Mel1a and Mel1c but not Mel1b are associated with monochromatic light-induced B-lymphocyte proliferation in broilers. Domest. Anim. Endocrinol. 2013, 45, 206–215. [Google Scholar] [CrossRef]
- Chen, W.; Chen, L.F.; Zhang, M.B.; Xia, Y.P.; Zhao, Y.H.; Li, G.Z.; Wang, X.L. [Effects of different melatonin treatment regimens on the proliferation of endogenous neural stem cells in neonatal rats with hypoxic-ischemic brain damage]. Zhongguo Dang Dai Er Ke Za Zhi 2019, 21, 830–835. [Google Scholar]
- Niu, B.; Li, B.; Wu, C.; Wu, J.; Yan, Y.; Shang, R.; Bai, C.; Li, G.; Hua, J. Melatonin promotes goat spermatogonia stem cells (SSCs) proliferation by stimulating glial cell line-derived neurotrophic factor (GDNF) production in Sertoli cells. Oncotarget 2016, 7, 77532–77542. [Google Scholar] [CrossRef]
- Anawalt, B.D.; Bebb, R.A.; Matsumoto, A.M.; Groome, N.P.; Illingworth, P.J.; McNeilly, A.S.; Bremner, W.J. Serum inhibin B levels reflect Sertoli cell function in normal men and men with testicular dysfunction. J. Clin. Endocrinol. Metab. 1996, 81, 3341–3345. [Google Scholar]
- Roberts, V.; Meunier, H.; Sawchenko, P.E.; Vale, W. Differential production and regulation of inhibin subunits in rat testicular cell types. Endocrinology 1989, 125, 2350–2359. [Google Scholar] [CrossRef]
- Kaneko, H.; Noguchi, J.; Kikuchi, K.; Akagi, S.; Shimada, A.; Taya, K.; Watanabe, G.; Hasegawa, Y. Production and endocrine role of inhibin during the early development of bull calves. Biol. Reprod. 2001, 65, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, L.; Chen, S.; Chen, L.; Liu, Z.; Hou, X.; Chen, C.; Han, Y.; Wang, C.; Li, C.; et al. Association of inhibin-alpha gene polymorphisms with follicular cysts in large white sows. Theriogenology 2015, 84, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, C.; Chen, S.; Sun, L.; Li, H.; Chen, L.; Zhou, X. Effect of inhibin A on proliferation of porcine granulosa cells in vitro. Theriogenology 2018, 114, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Hua, G.; Ahmad, S.; Liang, A.; Han, L.; Wu, C.; Yang, F.; Yang, L. Action mechanism of inhibin alpha-subunit on the development of Sertoli cells and first wave of spermatogenesis in mice. PLoS ONE 2011, 6, e25585. [Google Scholar] [CrossRef]
- Park, S.; Park, M.Y.; Song, G.; Lim, W. Alpha-solanine inhibits cell proliferation via mitochondrial dysfunction and inhibin synthesis in mouse testis In vitro and In vivo. Chemosphere 2019, 235, 271–279. [Google Scholar] [CrossRef]
- Scott, M.; Bonnefin, P.; Vieyra, D.; Boisvert, F.M.; Young, D.; Bazett-Jones, D.P.; Riabowol, K. UV-induced binding of ING1 to PCNA regulates the induction of apoptosis. J. Cell Sci. 2001, 114 (Pt 19), 3455–3462. [Google Scholar]
- Alao, J.P.; Gamble, S.C.; Stavropoulou, A.V.; Pomeranz, K.M.; Lam, E.W.; Coombes, R.C.; Vigushin, D.M. The cyclin D1 proto-oncogene is sequestered in the cytoplasm of mammalian cancer cell lines. Mol. Cancer 2006, 5, 7. [Google Scholar] [CrossRef]
- Li, C.; Zhu, X.; Chen, S.; Chen, L.; Zhao, Y.; Jiang, Y.; Gao, S.; Wang, F.; Liu, Z.; Fan, R.; et al. Melatonin promotes the proliferation of GC-1 spg cells by inducing metallothionein-2 expression through ERK1/2 signaling pathway activation. Oncotarget 2017, 8, 65627–65641. [Google Scholar] [CrossRef]
- Heo, J.S.; Pyo, S.; Lim, J.Y.; Yoon, D.W.; Kim, B.Y.; Kim, J.H.; Kim, G.J.; Lee, S.G.; Kim, J. Biological effects of melatonin on human adiposederived mesenchymal stem cells. Int. J. Mol. Med. 2019, 44, 2234–2244. [Google Scholar]
- Fu, J.; Zhao, S.D.; Liu, H.J.; Yuan, Q.H.; Liu, S.M.; Zhang, Y.M.; Ling, E.A.; Hao, A.J. Melatonin promotes proliferation and differentiation of neural stem cells subjected to hypoxia in vitro. J. Pineal Res. 2011, 51, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Herlaar, E.; Brown, Z. p38 MAPK signalling cascades in inflammatory disease. Mol. Med. Today 1999, 5, 439–447. [Google Scholar] [CrossRef]
- Ge, J.; Zhou, Q.; Niu, J.; Wang, Y.; Yan, Q.; Wu, C.; Qian, J.; Yang, H. Melatonin Protects Intervertebral Disc from Degeneration by Improving Cell Survival and Function via Activation of the ERK1/2 Signaling Pathway. Oxid Med Cell Longev. 2019, 2019, 5120275. [Google Scholar] [CrossRef] [PubMed]
- Maria, S.; Samsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for MT2 melatonin receptors, MEK1/2, and MEK5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2018, 64. [Google Scholar] [CrossRef] [PubMed]
- Awad, H.; Halawa, F.; Mostafa, T.; Atta, H. Melatonin hormone profile in infertile males. Int. J. Androl. 2006, 29, 409–413. [Google Scholar] [CrossRef]
- Zhang, P.; Zheng, Y.; Lv, Y.; Li, F.; Su, L.; Qin, Y.; Zeng, W. Melatonin protects the mouse testis against heat-induced damage. Mol. Hum. Reprod. 2020, 26, 65–79. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′–3′) | Genebank No. | Size (bp) |
|---|---|---|---|
| SOX9 | F: GCTGTGGAGGCTGCTGAATGAG | NM_204281.1 | 236 |
| R: CGCTGATGCTGGAGGATGACTG | |||
| GATA4 | F: TGTCACCTCGCTTCTCCTTCTCC | XM_004935896.3 | 363 |
| R: AGTGCCCTGTGCCATCTCTCC |
| siRNA | Sequence (5′→3′) |
|---|---|
| Negative control | Sense: UUCUCCGAACGUGUCACGUTT Antisense: ACGUGACACGUUCGGAGAATT |
| INHA siRNA1 | Sense: GCGUCCCUCAACAUCUCUUTT Antisense: AAGAGAUGUUGAGGGACGCTT |
| INHA siRNA2 | Sense: CCACGGGAACUGUGCCGAATT Antisense: UUCGGCACAGUUCCCGUGGTT |
| INHA siRNA3 | Sense: ACCUCUGAUGGUGGCUACUTT Antisense: AGUAGCCACCAUCAGAGGUTT |
| Genes | Primer Sequence (5′–3′) | Genebank No. | Size (bp) |
|---|---|---|---|
| PCNA | F: GCAGATGTTCCTCTCGTTGTGGAG | NM_204170.2 | 95 |
| R: GAGCCTTCCTGCTGGTCTTCAATC | |||
| CCND1 | F: TCGGTGTCCTACTTCAAGTG | NM_205381.1 | 273 |
| R: GGAGTTGTCGGTGTAAATGC | |||
| INHA | F: ACCGCAGAGATGTCCTCGAAGAG | NM_001031257.1 | 95 |
| R: GCACGGCACGTCTGTGGAAG | |||
| GAPDH | F: TAAGCGTGTTATCATCTC | NM_204305.1 | 83 |
| R: GGGACTTGTCATATTTCT |
| Antibodies | Cat NO. | Source | Dilution |
|---|---|---|---|
| PCNA | bs-2006R | Bioss, Beijing, China | 1:300 |
| CCND1 | PAB9944 | Abnova, Taipei, Taiwan, China | 1:300 |
| GAPDH | 60004-1-lg | ProteinTech, Chicago, IL, USA | 1:10,000 |
| ERK1/2 | orb315598 | Biorbyt, California, USA | 1:1000 |
| p-ERK1/2 | orb338969 | Biorbyt, California, USA | 1:1000 |
| Goat Anti-Rabbit IgG | SA00001-2 | ProteinTech, Chicago, IL, USA | 1:10,000 |
| Goat Anti-mouse IgG | SA00001-1 | ProteinTech, Chicago, IL, USA | 1:10,000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, K.; Wang, J.; Liu, H.; Zhao, J.; Lu, W. Melatonin Promotes the Proliferation of Chicken Sertoli Cells by Activating the ERK/Inhibin Alpha Subunit Signaling Pathway. Molecules 2020, 25, 1230. https://doi.org/10.3390/molecules25051230
Xu K, Wang J, Liu H, Zhao J, Lu W. Melatonin Promotes the Proliferation of Chicken Sertoli Cells by Activating the ERK/Inhibin Alpha Subunit Signaling Pathway. Molecules. 2020; 25(5):1230. https://doi.org/10.3390/molecules25051230
Chicago/Turabian StyleXu, Ke, Jun Wang, Hongyu Liu, Jing Zhao, and Wenfa Lu. 2020. "Melatonin Promotes the Proliferation of Chicken Sertoli Cells by Activating the ERK/Inhibin Alpha Subunit Signaling Pathway" Molecules 25, no. 5: 1230. https://doi.org/10.3390/molecules25051230
APA StyleXu, K., Wang, J., Liu, H., Zhao, J., & Lu, W. (2020). Melatonin Promotes the Proliferation of Chicken Sertoli Cells by Activating the ERK/Inhibin Alpha Subunit Signaling Pathway. Molecules, 25(5), 1230. https://doi.org/10.3390/molecules25051230