
The Effects of Houttuynia cordata Thunb and Piper ribesioides Wall Extracts on Breast Carcinoma Cell Proliferation, Migration, Invasion and Apoptosis



Abstract
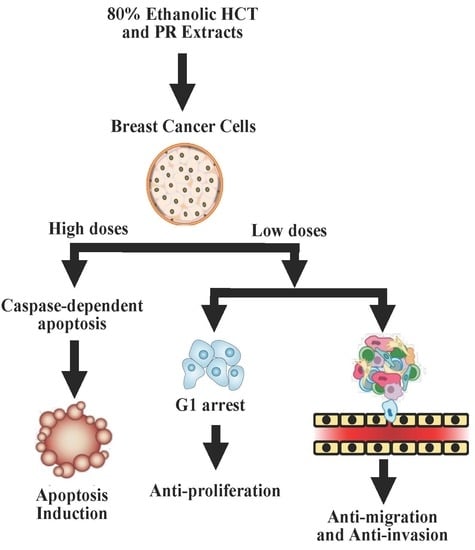
1. Introduction
2. Results
2.1. Identification of Phytochemical Compositions in H. cordata and P. ribesioides Extracts
2.2. Cytotoxic Effects of H. cordata and P. ribesioides Extracts on Breast Cancer Cells, PBMCs and MCF-10A
2.3. Effects of H. cordata and P. ribesioides Extracts on Growth Inhibition in Breast Cancer Cells
2.4. Effect of H. cordata and P. ribesioides on Breast Cancer Migration and Invasion
2.5. Effects of H. cordata and P. ribesioides Extracts on Apoptosis Induction in Breast Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Plant Sample Preparation
4.3. Measurement of Total Phenolic Compounds
4.4. Determination of Total Flavonoid Compound
4.5. Determination of Anti-OxidantActivity by 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Assay
4.6. Determination of Phenolic Compounds by Reversed Phase—High Performance Liquid Chromatography (HPLC)
4.7. Characterization of Phytochemicals by Using GC-MS Technique
4.8. Cell Culture
4.9. Cell Viability and Proliferation Assay
4.10. Apoptosis Determination by Flow Cytometry
4.11. Determination of Caspase 9, -8 and -3 Activities
4.12. Western Blotting
4.13. Colony Formation Assay
4.14. Cell Cycle Assay
4.15. Wound-Healing Assay
4.16. Gelatin Zymography for MMP-2 and MMP-9 Secretions Assay
4.17. Transwell-Migration and Invasion Assay
4.18. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA A Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Scully, O.J.; Bay, B.-H.; Yip, G.; Yu, Y. Breast Cancer Metastasis. Cancer Genom.-Proteom. 2012, 9, 311–320. [Google Scholar] [PubMed]
- Weigelt, B.; Peterse, J.L.; van’t Veer, L.J. Breast cancer metastasis: Markers and models. Nat. Rev. Cancer 2005, 5, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, B.P. Snail: More than EMT. Cell Adh Migr. 2010, 4, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77. [Google Scholar] [CrossRef]
- Sayers, T.J. Targeting the extrinsic apoptosis signaling pathway for cancer therapy. Cancer Immunol. Immunother 2011, 60, 1173–1180. [Google Scholar] [CrossRef]
- Kumar, M.; Prasad, S.K.; Hemalatha, S. A current update on the phytopharmacological aspects of Houttuynia cordata Thunb. Pharmacogn Rev. 2014, 8, 22–35. [Google Scholar] [CrossRef]
- Salleh, W.M.N.H.W.; Ahmad, F.; Khong, H.Y. Chemical composition of Piper stylosum Miq. and Piper ribesioides Wall. essential oils, and their antioxidant, antimicrobial and tyrosinase inhibition activities. B Latinoam Caribe Pl 2014, 13, 488–497. [Google Scholar]
- Kumnerdkhonkaen, P.; Saenglee, S.; Asgar, M.A.; Senawong, G.; Khongsukwiwat, K.; Senawong, T. Antiproliferative activities and phenolic acid content of water and ethanolic extracts of the powdered formula of Houttuynia cordata Thunb. fermented broth and Phyllanthus emblica Linn. fruit. BMC Complement Altern Med. 2018, 18, 130. [Google Scholar] [CrossRef]
- Cohen, Z.; Maimon, Y.; Samuels, N.; Berger, R. Role of reactive oxygen species in the anticancer activity of botanicals: Comparing sensitivity profiles. Oncol. Lett. 2017, 13, 2642–2648. [Google Scholar] [CrossRef][Green Version]
- Chen, Y.-F.; Yang, J.-S.; Chang, W.-S.; Tsai, S.-C.; Peng, S.-F.; Zhou, Y.-R. Houttuynia cordata Thunb extract modulates G 0/G 1 arrest and Fas/CD95-mediated death receptor apoptotic cell death in human lung cancer A549 cells. J. Biomes. Sci. 2013, 20, 18. [Google Scholar] [CrossRef]
- Tian, Y.; Wan, H.; Tan, G. Cell cycle-related kinase in carcinogenesis. Oncol. Lett. 2012, 4, 601–606. [Google Scholar] [CrossRef]
- Skomedal, H.; Kristensen, G.B.; Lie, A.K.; Holm, R. Aberrant expression of the cell cycle associated proteins TP53, MDM2, p21, p27, cdk4, cyclin D1, RB, and EGFR in cervical carcinomas. Gynecol. Oncol. 1999, 73, 223–228. [Google Scholar] [CrossRef]
- Lai, K.-C.; Chiu, Y.-J.; Tang, Y.-J.; Lin, K.-L.; Chiang, J.-H.; Jiang, Y.-L.; Jen, H.-F.; Kuo, Y.-H.; Agamaya, S.; Chung, J.-G. Houttuynia cordata Thunb extract inhibits cell growth and induces apoptosis in human primary colorectal cancer cells. Anticancer Res. 2010, 30, 3549–3556. [Google Scholar]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Khan, T.; Sharifi-Rad, J.; Ozleyen, A.; Turkdonmez, E.; et al. Piper Species: A Comprehensive Review on Their Phytochemistry, Biological Activities and Applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef]
- Yeh, Y.-C.; Liu, T.-J.; Lai, H.-C. Shikonin induces apoptosis, necrosis, and premature senescence of human A549 lung cancer cells through upregulation of p53 expression. Evid Based Complement Alternat. Med. 2015, 2015. [Google Scholar] [CrossRef]
- Grütter, M.G. Caspases: Key players in programmed cell death. Curr. Opin. Struc. Biol. 2000, 10, 649–655. [Google Scholar] [CrossRef]
- Jänicke, R.U.; Ng, P.; Sprengart, M.L.; Porter, A.G. Caspase-3 is required for α-fodrin cleavage but dispensable for cleavage of other death substrates in apoptosis. J. Biol. Chem. 1998, 273, 15540–15545. [Google Scholar] [CrossRef]
- Zhao, Y.; Sui, X.; Ren, H. From procaspase-8 to caspase-8: Revisiting structural functions of caspase-8. J. Cell Physiol. 2010, 225, 316–320. [Google Scholar] [CrossRef]
- Würstle, M.L.; Laussmann, M.A.; Rehm, M. The central role of initiator caspase-9 in apoptosis signal transduction and the regulation of its activation and activity on the apoptosome. Exp. Cell Res. 2012, 318, 1213–1220. [Google Scholar] [CrossRef]
- Yan, N.; Shi, Y. Mechanisms of apoptosis through structural biology. Annu. Rev. Cell Dev. Biol. 2005, 21, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol 2013, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef]
- Meng, J.; Leung, K.S.-Y.; Jiang, Z.; Dong, X.; Zhao, Z.; Xu, L.-J. Establishment of HPLC-DAD-MS fingerprint of fresh Houttuynia cordata. Chem. Pharm. Bull 2005, 53, 1604–1609. [Google Scholar] [CrossRef]
- Xin-Yi, D.; Yi, J.; Li-Ping, Y. Systematic analysis of components and contents in Houttuynia cordata Thunb. Tradit. Med. Res. 2017, 2, 176–188. [Google Scholar]
- Ross, D.T.; Perou, C.M. A comparison of gene expression signatures from breast tumors and breast tissue derived cell lines. Dis. Markers 2001, 17, 99–109. [Google Scholar] [CrossRef]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.-P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef]
- J. Deka, S.; Gorai, S.; Manna, D.; Trivedi, V. Evidence of PKC binding and translocation to explain the anticancer mechanism of chlorogenic acid in breast cancer cells. Curr. Mol. Med. 2017, 17, 79–89. [Google Scholar] [CrossRef]
- Scott, I.M.; Jensen, H.R.; Philogène, B.J.; Arnason, J.T. A review of Piper spp.(Piperaceae) phytochemistry, insecticidal activity and mode of action. Phytochem. Rev. 2008, 7, 65. [Google Scholar] [CrossRef]
- Scott, I.M.; Puniani, E.; Jensen, H.; Livesey, J.F.; Poveda, L.; Sánchez-Vindas, P.; Durst, T.; Arnason, J.T. Analysis of Piperaceae germplasm by HPLC and LCMS: A method for isolating and identifying unsaturated amides from Piper spp extracts. J. Agric. Food Chem. 2005, 53, 1907–1913. [Google Scholar] [CrossRef]
- Abdelhamed, S.; Yokoyama, S.; Refaat, A.; Ogura, K.; Yagita, H.; Awale, S.; Saiki, I. Piperine enhances the efficacy of TRAIL-based therapy for triple-negative breast cancer cells. Anticancer Res. 2014, 34, 1893–1899. [Google Scholar] [PubMed]
- Greenshields, A.L.; Doucette, C.D.; Sutton, K.M.; Madera, L.; Annan, H.; Yaffe, P.B.; Knickle, A.F.; Dong, Z.; Hoskin, D.W. Piperine inhibits the growth and motility of triple-negative breast cancer cells. Cancer Lett. 2015, 357, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Leung, K.S.-Y.; Jiang, Z.; Dong, X.; Zhao, Z. Establishment of GC-MS fingerprint of fresh Houttuynia cordata. Chem. Pharm. Bull. 2005, 53, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Qi, M.; Zhang, C.; Zhou, S.; Fu, R.; Huang, J. Gas chromatography–mass spectrometry analysis of volatile compounds from Houttuynia cordata Thunb after extraction by solid-phase microextraction, flash evaporation and steam distillation. Anal. Chim. Acta 2005, 531, 97–104. [Google Scholar] [CrossRef]
- Lu, H.; Liang, Y.; Yi, L.; Wu, X. Anti-inflammatory effect of Houttuynia cordata injection. J. Ethnopharmacol. 2006, 104, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-L.; Zhang, Z.-J.; Choksi, S.; Shan, S.; Lu, Z.; Croce, C.M.; Alnemri, E.S.; Korngold, R.; Huang, Z. Cell permeable Bcl-2 binding peptides: A chemical approach to apoptosis induction in tumor cells. Cancer Res. 2000, 60, 1498–1502. [Google Scholar]
- Zafaryab, M.; Fakhri, K.U.; Khan, M.A.; Hajela, K.; Rizvi, M.M.A. In vitro assessment of cytotoxic and apoptotic potential of palmitic acid for breast cancer treatment. Int. J. Life Sci. Res. 2019, 7, 166–174. [Google Scholar]
- Pietenpol, J.A.; Stewart, Z.A. Cell cycle checkpoint signaling: Cell cycle arrest versus apoptosis. Toxicology 2002, 475–481. [Google Scholar] [CrossRef]
- Oakman, C.; Viale, G.; Di Leo, A. Management of triple negative breast cancer. The Breast 2010, 19, 312–321. [Google Scholar] [CrossRef]
- Huang, L.; Li, A.; Liao, G.; Yang, F.; Yang, J.; Chen, X.; Jiang, X. Curcumol triggers apoptosis of p53 mutant triple-negative human breast cancer MDA-MB 231 cells via activation of p73 and PUMA. Oncol. Lett. 2017, 14, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Shao, Z.; Wu, J. WAF1/CIP1/p21 gene in wild type p53 and mutant p53 human breast cancer cell lines in relation to its cytobiological features. Zhonghua Zhong Liu Za Zhi 1998, 20, 181–184. [Google Scholar] [PubMed]
- Rachakhom, W.; Khaw-On, P.; Pompimon, W.; Banjerdpongchai, R. Dihydrochalcone Derivative Induces Breast Cancer Cell Apoptosis via Intrinsic, Extrinsic, and ER Stress Pathways but Abolishes EGFR/MAPK Pathway. Biomed. Res. Int. 2019, 2019, 7298539. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Hwang, I.-H.; Jang, I.-S.; Kim, M.; Bang, I.S.; Park, S.J.; Chung, Y.-J.; Joo, J.-C.; Lee, M.-G. Houttuynia cordata Thunb promotes activation of HIF-1A–FOXO3 and MEF2A pathways to induce apoptosis in human HepG2 hepatocellular carcinoma cells. Integr. Cancer Ther. 2017, 16, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Ullmannová, V.; Stöckbauer, P.; Hradcová, M.; Souček, J.; Haškovec, C. Relationship between cyclin D1 and p21Waf1/Cip1 during differentiation of human myeloid leukemia cell lines. Leukemia Res. 2003, 27, 1115–1123. [Google Scholar] [CrossRef]
- Davidson, B.; Reich, R.; Risberg, B.; Nesland, J.M. The biological role and regulation of matrix metalloproteinases (MMP) in cancer. Arkhiv Patologii 2002, 64, 47–53. [Google Scholar]
- Liotta, L.; Stetler-Stevenson, W. Metalloproteinases and cancer invasion. Semin. Cancer Biol. 1990, 1, 99–106. [Google Scholar]
- Stetler-Stevenson, W.G. Type IV collagenases in tumor invasion and metastasis. Cancer Metastasis Rev. 1990, 9, 289–303. [Google Scholar] [CrossRef]
- Kim, D.; Rhee, S. Matrix metalloproteinase-2 regulates MDA-MB-231 breast cancer cell invasion induced by active mammalian diaphanous-related formin 1. Mol. Med. Rep. 2016, 14, 277–282. [Google Scholar] [CrossRef]
- Degterev, A.; Boyce, M.; Yuan, J. A decade of caspases. Oncogene 2003, 22, 8543. [Google Scholar] [CrossRef]
- Gillissen, B.; Essmann, F.; Graupner, V.; Stärck, L.; Radetzki, S.; Dörken, B.; Schulze-Osthoff, K.; Daniel, P.T. Induction of cell death by the BH3-only Bcl-2 homolog Nbk/Bik is mediated by an entirely Bax-dependent mitochondrial pathway. Embo. J. 2003, 22, 3580–3590. [Google Scholar] [CrossRef] [PubMed]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, T.; Strasser, A.; Jost, P.J. Fas death receptor signalling: Roles of Bid and XIAP. Cell Death Differ. 2012, 19, 42. [Google Scholar] [CrossRef] [PubMed]
- Kantari, C.; Walczak, H. Caspase-8 and Bid: Caught in the act between death receptors and mitochondria. BBA-Mol. Cell Res. 2011, 1813, 558–563. [Google Scholar] [CrossRef]
- Green, D.R. Apoptotic pathways: The roads to ruin. Cell 1998, 94, 695–698. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Walsh, J.G.; Cullen, S.P.; Sheridan, C.; Lüthi, A.U.; Gerner, C.; Martin, S.J. Executioner caspase-3 and caspase-7 are functionally distinct proteases. Proc. Natl. Acad. Sci. USA 2008, 105, 12815–12819. [Google Scholar] [CrossRef]
- Lakhani, S.A.; Masud, A.; Kuida, K.; Porter, G.A.; Booth, C.J.; Mehal, W.Z.; Inayat, I.; Flavell, R.A. Caspases 3 and 7: Key mediators of mitochondrial events of apoptosis. Science 2006, 311, 847–851. [Google Scholar] [CrossRef]
- Fukumoto, L.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Baskar, R.; Shrisakthi, S.; Sathyapriya, B.; Shyampriya, R.; Nithya, R.; Poongodi, P. Antioxidant potential of peel extracts of banana varieties (Musa sapientum). Food Sci. Nutr. 2011, 2, 1128. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pintha, K.; Yodkeeree, S.; Pitchakarn, P.; Limtrakul, P. Anti-invasive activity against cancer cells of phytochemicals in red jasmine rice (Oryza sativa L.). Asian Pac. J. Cancer Prev. 2014, 15, 4601–4607. [Google Scholar] [CrossRef] [PubMed]
- Promprom, W.; Chatan, W. GC-MS Analysis and antioxidant activity of Bauhinia nakhonphanomensis leaf ethanolic extract. Pharmacogn Res. 2017, 9. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112. [Google Scholar] [CrossRef] [PubMed]
- Wlodkowic, D.; Skommer, J.; Darzynkiewicz, Z. Flow cytometry-based apoptosis detection. In Apoptosis; Springer: New York, NY, USA, 2009; pp. 19–32. [Google Scholar]
- Zhang, M.; Chen, J.; Jia, Z.; Zhou, H.; Zhang, X. Effects of chrysotile fibers on apoptosis of human bronchial epithelial cell line (BEAS-2B) in vitro. Joem 2009, 26, 155–158. [Google Scholar]
- Wudtiwai, B.; Pitchakarn, P.; Banjerdpongchai, R. Alpha-mangostin, an active compound in Garcinia mangostana, abrogates anoikis-resistance in human hepatocellular carcinoma cells. Toxico. Vitro 2018, 53, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Clair, D.K.S.; Fang, F.; Warren, G.W.; Rangnekar, V.M.; Crooks, P.A.; Clair, W.H.S. The radiosensitization effect of parthenolide in prostate cancer cells is mediated by nuclear factor-κB inhibition and enhanced by the presence of PTEN. Molecules 2007, 6, 2477–2486. [Google Scholar]
- Ascione, F.; Caserta, S.; Guido, S. The wound healing assay revisited: A transport phenomena approach. Chemistry 2017, 160, 200–209. [Google Scholar] [CrossRef]
- Toth, M.; Sohail, A.; Fridman, R. Assessment of gelatinases (MMP-2 and MMP-9) by gelatin zymography. In Metastasis Research Protocols; Springer: New York, NY, USA, 2012; pp. 121–135. [Google Scholar]
- Xu, J.; Zhang, W.; Tang, L.; Chen, W.; Guan, X. Epithelial-mesenchymal transition induced PAI-1 is associated with prognosis of triple-negative breast cancer patients. Gene 2018, 670, 7–14. [Google Scholar] [CrossRef]
Sample Availability: Not available. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Total Phenolic Content (mg Gallic Acid/g Extract) | Total Flavonoid (mg Catechin/g Extract) | DPPH-Radical Scavenging Activity (IC50 (μg/mL)) |
|---|---|---|---|
| H. cordata | 119.8 ± 3.0 | 93.8 ± 1.3 | 234.6 ± 11.9 |
| P. ribesioides | 203.8 ± 10.8 | 95.4 ± 2.5 | 153.8 ± 4.4 |
| Retention Times (Rt) | H. cordata | P. ribesioides | Compounds |
|---|---|---|---|
| 3.186 | 0.435 ± 0.01 | 0.827 ± 0.08 | Gallic acid |
| 7.908 | 1.64 ± 0.43 | 1.128 ± 0.13 | Catechin |
| 9.012 | 25.5 ± 3.41 | n.d. | Chlorogenic acid |
| 10.904 | 0.844 ± 0.14 | 0.543 ± 0.03 | Vanilic acid |
| 13.132 | 0.621 ± 0.02 | 1.199 ± 0.02 | Ferulic acid |
| 13.990 | 0.116 ± 0.02 | n.d. | p-Courmaric acid |
| 17.167 | 44.00 ± 5.61 | n.d. | Rutin |
| 18.288 | 1.49 ± 0.04 | 1.89 ± 0.01 | Rosmarinic acid |
| 21.361 | 0.196 ± 0.05 | 0.465 ± 0.05 | Quercetin |
| 22.329 | n.d. | 0.124 ± 0.01 | Apigenin |
| No. | Retention Time | Name of Compounds | % of Total |
|---|---|---|---|
| 1 | 7.926 | Decanoic acid | 9.42 |
| 2 | 10.329 | Dodecanoic acid | 2.57 |
| 3 | 12.578 | Tetradecanoic acid | 0.54 |
| 4 | 12.979 | Octadecane | 0.9 |
| 5 | 14.558 | n-Hexadecanoic acid | 10.5 |
| 6 | 14.878 | Eicosane | 0.59 |
| 7 | 16.034 | 9,12-Octadecadienoic acid | 1.78 |
| 8 | 16.091 | 9,12-Octadecatrienoic acid | 11.67 |
| 9 | 16.257 | Octadecanic acid | 2.93 |
| 10 | 21.716 | 9,17-Octadecadienal | 0.78 |
| 11 | 33.629 | alpha-Tocopherol | 5.33 |
| 12 | 34.711 | Sesamin | 2.01 |
| 13 | 42.287 | Cholest-5-en-3-ol | 14.74 |
| No. | Retention Time | Name of Compounds | % of Total |
|---|---|---|---|
| 1 | 10.324 | Dodecanoic acid | 0.05 |
| 2 | 12.584 | Tetradecanoic acid | 0.43 |
| 3 | 14.581 | n-Hexadecanoic acid | 6.82 |
| 4 | 16.051 | 9,12-Octadecanoic acid | 3.84 |
| 5 | 16.103 | cis-Vaccenic acid | 5.1 |
| 6 | 16.269 | Octadecanoic acid | 1.94 |
| 7 | 26.540 | Piperine | 49.51 |
| 8 | 42.292 | β-Sitrosterol | 3.46 |
| MCF-7 | ||||||
| H. cordataExtract (µg/mL) | P. ribesioidesExtract (µg/mL) | |||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| IC20 | 129.3 ± 6.9 | 82.4 ± 7.1 | 129.7 ± 3.5 | 93.2 ± 7.2 | 56.4 ± 9.7 | 66.4 ± 12.2 |
| IC50 | 347.4 ± 10.8 | 187.9 ± 18.2 | 210.8 ± 3.1 | 167.0 ± 11.0 | 99.5 ± 12.0 | 98.4 ± 14.1 |
| MDA-MB-231 | ||||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| IC20 | 184.6 ± 14.0 | 66.9 ± 5.3 | 64.8 ± 3.8 | 174.6 ± 13.8 | 146.3 ± 5.6 | 149.5 ± 8.5 |
| IC50 | 667.8 ± 11.7 | 279.6 ± 4.4 | 290.7 ± 11.9 | 484.5 ± 14.3 | 264.1 ± 12.2 | 254.1 ± 8.0 |
| PBMCs | ||||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| IC20 | >1000 µg/mL | |||||
| IC50 | >1000 µg/mL | |||||
| MCF-10A | ||||||
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| IC20 | 514.6 ± 5.3 | 372.3 ± 9.1 | 275.9 ± 2.1 | 353.1 ± 4.5 | 256.3 ± 6.5 | 264.5 ± 2.5 |
| IC50 | >1000 µg/mL | 893.8 ± 8.4 | 679.5 ± 7.9 | 879.5 ± 2.6 | 523.0 ± 1.7 | 442.9 ± 1.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subhawa, S.; Chewonarin, T.; Banjerdpongchai, R. The Effects of Houttuynia cordata Thunb and Piper ribesioides Wall Extracts on Breast Carcinoma Cell Proliferation, Migration, Invasion and Apoptosis. Molecules 2020, 25, 1196. https://doi.org/10.3390/molecules25051196
Subhawa S, Chewonarin T, Banjerdpongchai R. The Effects of Houttuynia cordata Thunb and Piper ribesioides Wall Extracts on Breast Carcinoma Cell Proliferation, Migration, Invasion and Apoptosis. Molecules. 2020; 25(5):1196. https://doi.org/10.3390/molecules25051196
Chicago/Turabian StyleSubhawa, Subhawat, Teera Chewonarin, and Ratana Banjerdpongchai. 2020. "The Effects of Houttuynia cordata Thunb and Piper ribesioides Wall Extracts on Breast Carcinoma Cell Proliferation, Migration, Invasion and Apoptosis" Molecules 25, no. 5: 1196. https://doi.org/10.3390/molecules25051196
APA StyleSubhawa, S., Chewonarin, T., & Banjerdpongchai, R. (2020). The Effects of Houttuynia cordata Thunb and Piper ribesioides Wall Extracts on Breast Carcinoma Cell Proliferation, Migration, Invasion and Apoptosis. Molecules, 25(5), 1196. https://doi.org/10.3390/molecules25051196