Interactive Effect of Melatonin and UV-C on Phenylpropanoid Metabolite Production and Antioxidant Potential in Callus Cultures of Purple Basil (Ocimum basilicum L. var purpurascens)

 ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
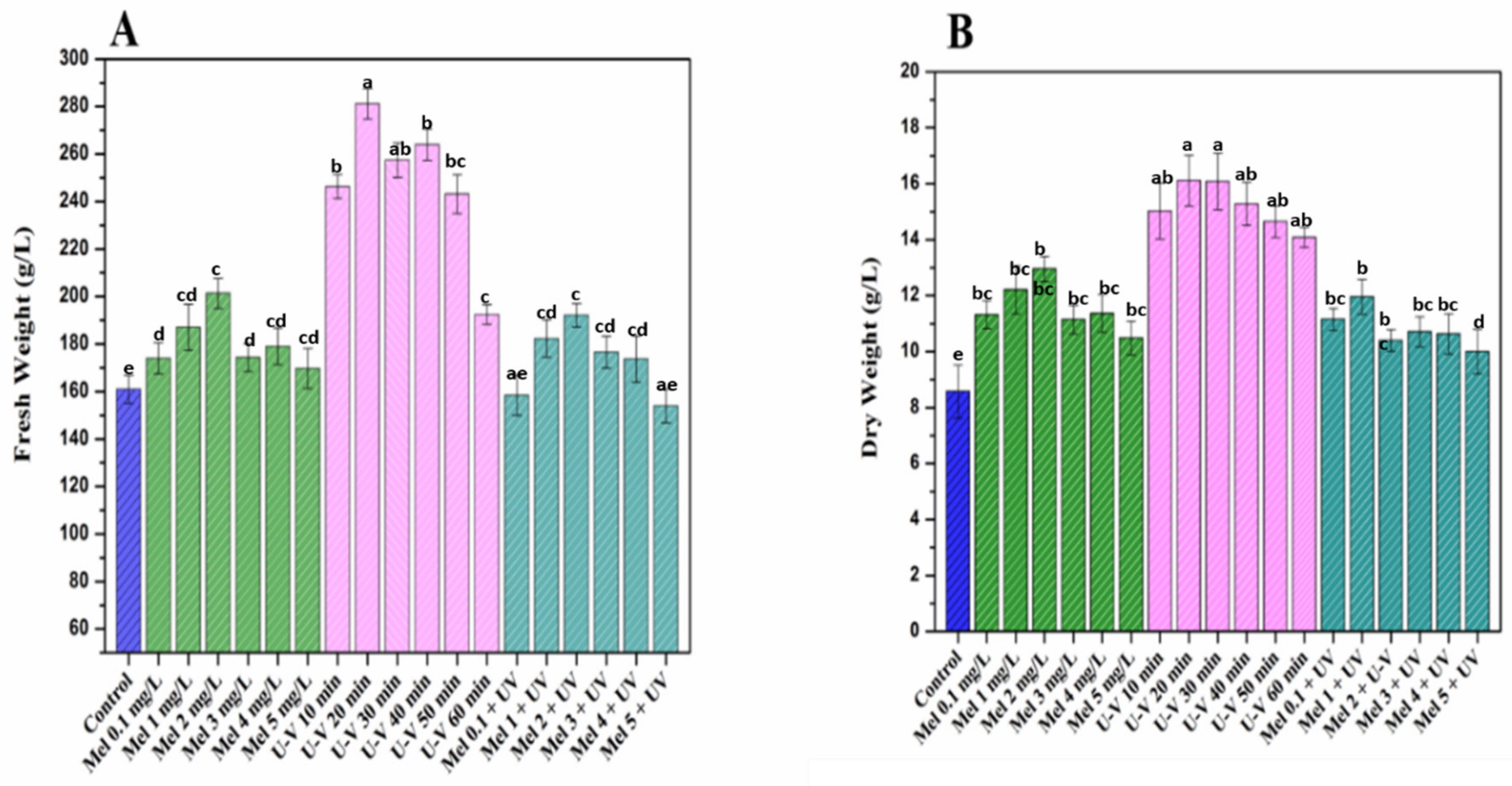
2.1. Biomass Accumulation under Melatonin and UV-C Treatments
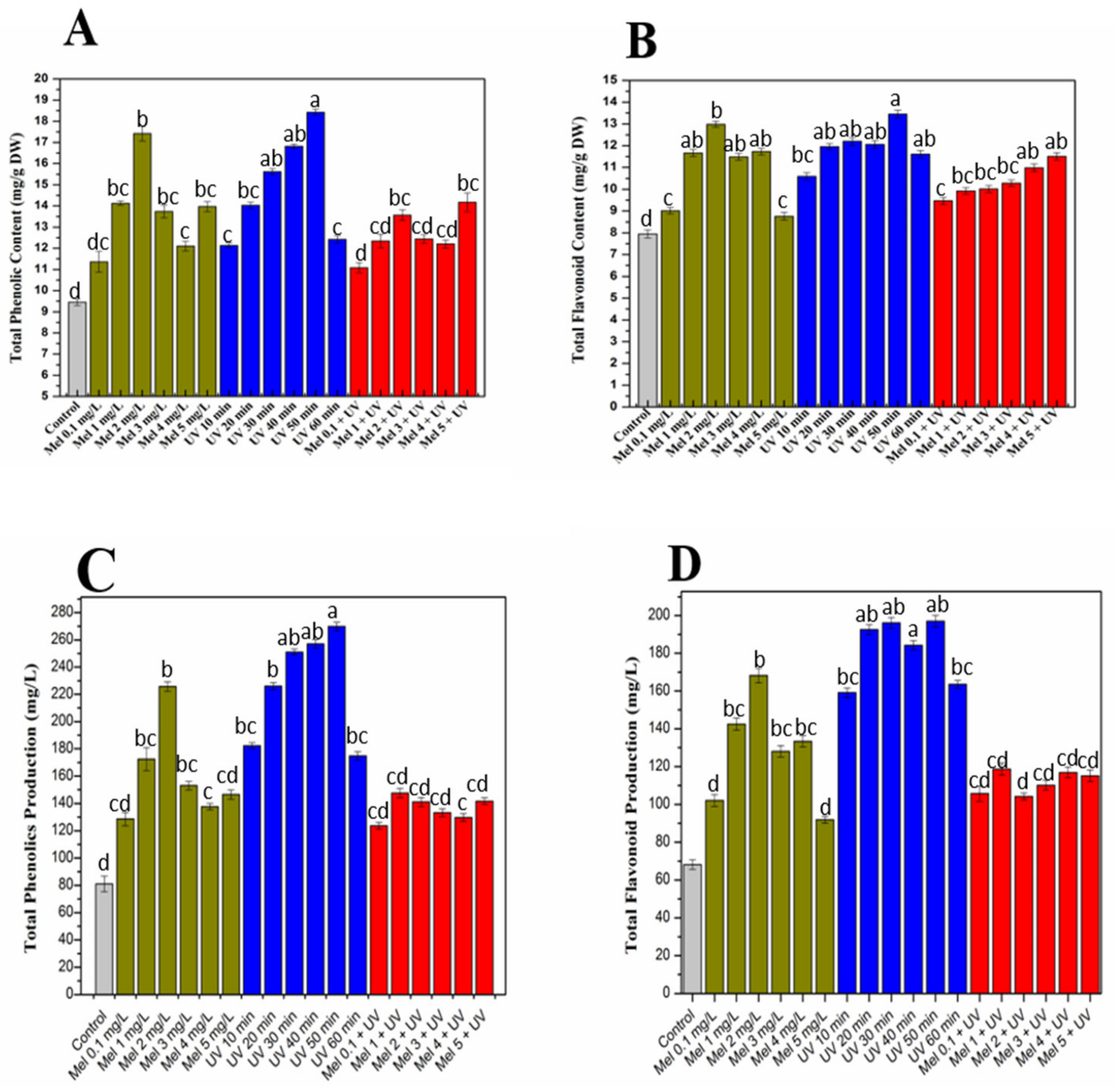
2.2. Influence UV-C radiations and Melatonin on Total Phenolic and Flavonoid Productions
2.3. Quantification of Phenylpropanoid Metabolites Via HPLC
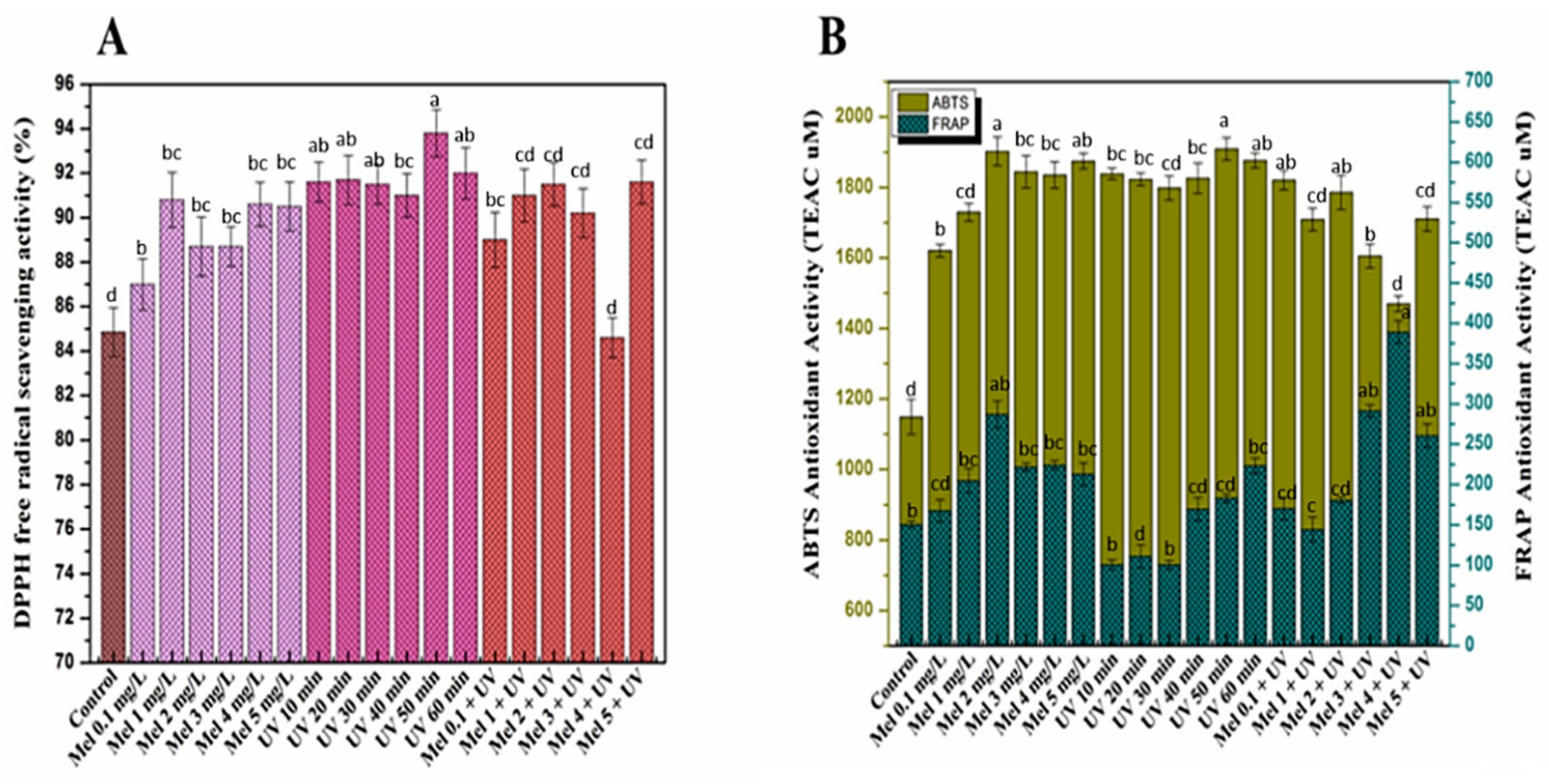
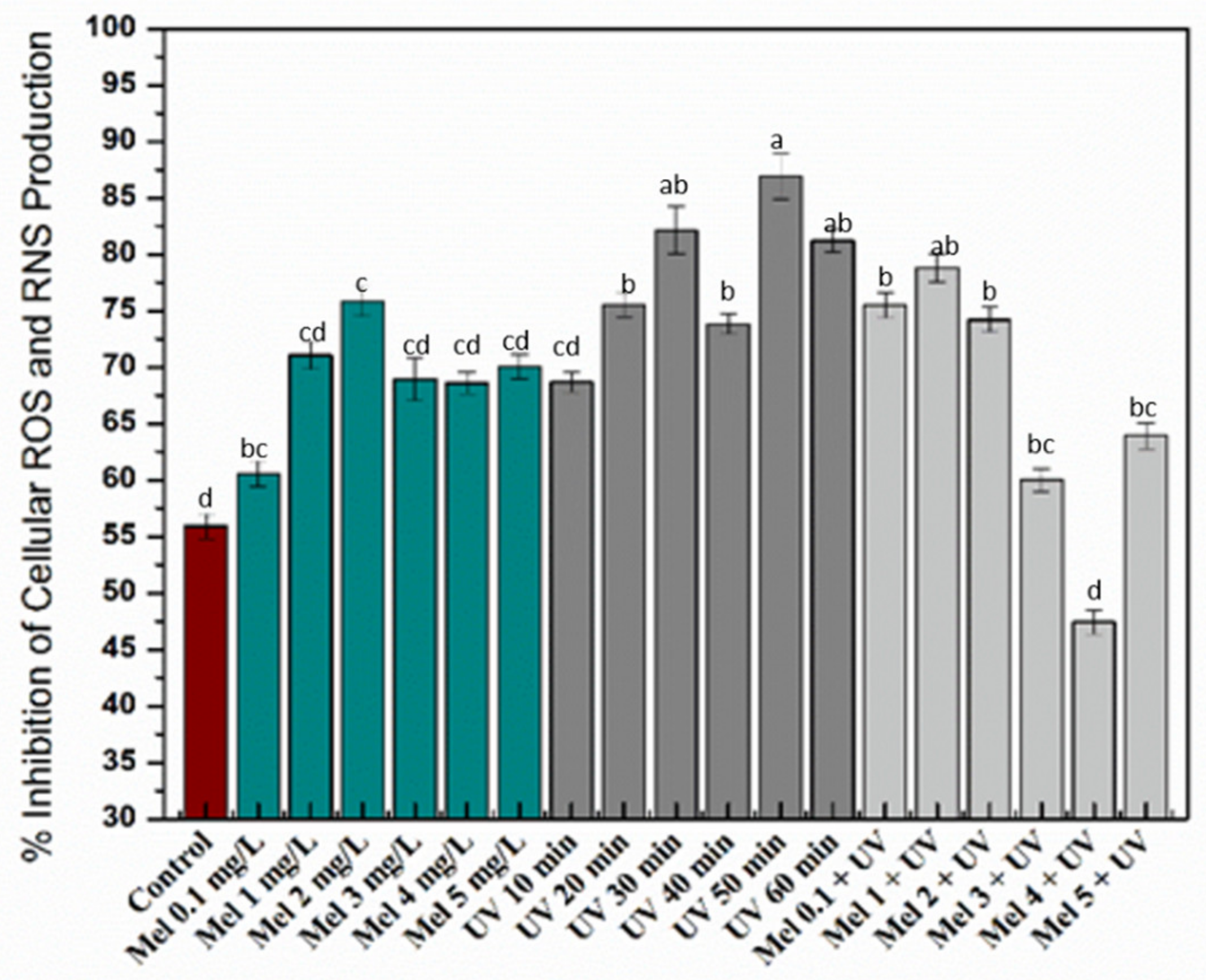
2.4. Antioxidant Activities of Purple Basil Callus Culture under UV-C light and Melatonin Treatments
3. Materials and Methods
3.1. Seed Germination and Callogenesis
3.2. Elicitors Treatment on Callus Culture
3.2.1. UV-C Treatment
3.2.2. Melatonin Treatment
3.2.3. Combined Melatonin + UV-C Treatment
3.3. Preparation of Sample Extracts
3.4. Total Flavonoid and Phenolic Estimation
3.5. Metabolic Quantification by HPLC Analysis
3.6. Estimation of Antioxidant Potential
3.6.1. DPPH Radical Scavenging Activity (%)
3.6.2. Ferric-Reducing Antioxidant Power (FRAP) Assay
3.6.3. Antioxidant ABTS Assay
3.6.4. Cellular Antioxidant Assay
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ABTS | 2,2-Azinobis 3-ethylbenzthiazoline-6-sulphonic acid |
| CA | Caffeic acid |
| CHS | Chalcone synthase |
| DHR-123 | Dihydrorhodamine-123 |
| DMSO | Dimethyl sulfoxide |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| DW | Dry weight |
| ET | Electron transfer |
| FRAP | Ferric-reducing antioxidant power |
| FW | Fresh weight |
| HAT | Hydrogen atom transfer |
| HPLC | High-performance liquid chromatography |
| Mel | Melatonin |
| MS | Murashige and Skoog |
| NAA | α-naphthalene acetic acid |
| PAL | Phenylalanine ammonia-lyase |
| RA | Rosmarinic acid |
| ROS | Reactive oxygen species |
| RNS | Reactive nitrogen species |
| TEAC | Trolox C equivalent antioxidant capacity |
| TFC: | Total flavonoid content |
| TFP | Total flavonoid production |
| TPP | Total phenolic production |
| TPC | Total phenolic content |
| UV-C | Ultraviolet radiation C |
References
- Kasem, M. Micropropagation and In Vitro Secondary Metabolites Production of Ocimum Species. J. Plant Prod. 2017, 8, 473–484. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, S.Y. Antioxidant activity and phenolic compounds in selected herbs. J. Agric. Food Chem. 2001, 49, 5165–5170. [Google Scholar] [CrossRef]
- Javanmardi, J.; Khalighi, A.; Kashi, A.; Bais, H.; Vivanco, J. Chemical characterization of basil (Ocimum basilicum L.) found in local accessions and used in traditional medicines in Iran. J. Agric. Food Chem. 2002, 50, 5878–5883. [Google Scholar] [CrossRef] [PubMed]
- Shan, B.; Cai, Y.Z.; Sun, M.; Corke, H. Antioxidant capacity of 26 spice extracts and characterization of their phenolic constituents. J. Agric. Food Chem. 2005, 53, 7749–7759. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Scagel, C.F. Chicoric acid found in basil (Ocimum basilicum L.) leaves. Food Chem. 2009, 115, 650–656. [Google Scholar] [CrossRef]
- Bowen-Forbes, C.S.; Zhang, Y.; Nair, M.G. Anthocyanin content, antioxidant, anti-inflammatory and anticancer properties of blackberry and raspberry fruits. J. Food Compos. Anal. 2010, 23, 554–560. [Google Scholar] [CrossRef]
- La Camera, S.; Gouzerh, G.; Dhondt, S.; Hoffmann, L.; Fritig, B.; Legrand, M.; Heitz, T. Metabolic reprogramming in plant innate immunity: The contributions of phenylpropanoid and oxylipin pathways. Immunol. Rev. 2004, 198, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Nazir, M.; Tungmunnithum, D.; Bose, S.; Drouet, S.; Garros, L.; Giglioli-Guivarc’h, N.; Abbasi, B.H.; Hano, C. Differential production of phenylpropanoid metabolites in callus cultures of Ocimum basilicum L. with distinct in vitro antioxidant activities and in vivo protective effects against UV stress. J. Agric. Food Chem. 2019, 67, 1847–1859. [Google Scholar] [CrossRef]
- Gaj, M.D. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Zhong, J.-J. Biochemical engineering of the production of plant-specific secondary metabolites by cell suspension cultures. In Plant Cells; Springer: Berlin, Heidelberg, 2001; pp. 1–26. [Google Scholar]
- Dörnenburg, H.; Knorr, D. Strategies for the improvement of secondary metabolite production in plant cell cultures. Enzym. Microb. Technol. 1995, 17, 674–684. [Google Scholar] [CrossRef]
- Roberts, S.C.; Shuler, M.L. Large-scale plant cell culture. Curr. Opin. Biotechnol. 1997, 8, 154–159. [Google Scholar] [CrossRef]
- Mulabagal, V.; Tsay, H.-S. Plant cell cultures-an alternative and efficient source for the production of biologically important secondary metabolites. Int. J. Appl. Sci. Eng. 2004, 2, 29–48. [Google Scholar]
- Yue, W.; Ming, Q.-L.; Lin, B.; Rahman, K.; Zheng, C.-J.; Han, T.; Qin, L.-P. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and its relationship to plant hormones. Ann. Bot. 2017, 121, 195–207. [Google Scholar] [CrossRef]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef]
- Arnao, M.; Hernández-Ruiz, J.; DeMello, J. Melatonin: Synthesis from tryptophan and its role in higher plants. In Amino Acids in Higher Plants; CAB eBooks, 2015; pp. 390–435. [Google Scholar]
- Nawaz, M.A.; Huang, Y.; Bie, Z.; Ahmed, W.; Reiter, R.J.; Niu, M.; Hameed, S. Melatonin: Current status and future perspectives in plant science. Front. Plant Sci. 2016, 6, 1230. [Google Scholar] [CrossRef]
- Meng, J.F.; Yu, Y.; Shi, T.C.; Fu, Y.S.; Zhao, T.; Zhang, Z.W. Melatonin treatment of pre-veraison grape berries modifies phenolic components and antioxidant activity of grapes and wine. Food Sci. Technol. 2019, 39, 35–42. [Google Scholar] [CrossRef]
- Coskun, Y.; Duran, R.E.; Kilic, S. Striking effects of melatonin on secondary metabolites produced by callus culture of rosemary (Rosmarinus officinalis L.). Plant Cell, Tissue Organ Cult. 2019, 138, 1–7. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Huhman, D.V.; Farag, M.A.; Smith, J.T.; May, G.D.; Mendes, P.; Dixon, R.A.; Sumner, L.W. Metabolic profiling of Medicago truncatula cell cultures reveals the effects of biotic and abiotic elicitors on metabolism. J. Exp. Bot. 2004, 56, 323–336. [Google Scholar] [CrossRef]
- Searles, P.S.; Kropp, B.R.; Flint, S.D.; Caldwell, M.M. Influence of solar UV-B radiation on peatland microbial communities of southern Argentinia. New Phytol. 2001, 152, 213–221. [Google Scholar] [CrossRef]
- Yin, X.; Singer, S.D.; Qiao, H.; Liu, Y.; Jiao, C.; Wang, H.; Li, Z.; Fei, Z.; Wang, Y.; Fan, C. Insights into the mechanisms underlying ultraviolet-C induced resveratrol metabolism in grapevine (V. amurensis Rupr.) Cv.“Tonghua-3”. Front. Plant Sci. 2016, 7, 503. [Google Scholar]
- Gangopadhyay, M.; Gantait, S.; Palchoudhury, S.; Ali, M.N.; Mondal, C.; Pal, A.K. UVC-priming mediated modulation of forskolin biosynthesis key genes against Macrophomina root rot of Coleus forskohlii A tissue culture based sustainable approach. Phytochem. Lett. 2016, 17, 36–44. [Google Scholar] [CrossRef]
- Li, D.; Luo, Z.; Mou, W.; Wang, Y.; Ying, T.; Mao, L. ABA and UV-C effects on quality, antioxidant capacity and anthocyanin contents of strawberry fruit (Fragaria ananassa Duch.). Postharvest Biol. Technol. 2014, 90, 56–62. [Google Scholar] [CrossRef]
- Freitas, A.; Moldão-Martins, M.; Costa, H.S.; Albuquerque, T.G.; Valente, A.; Sanches-Silva, A. Effect of UV-C radiation on bioactive compounds of pineapple (Ananas comosus L. Merr.) by-products. J. Sci. Food Agric. 2015, 95, 44–52. [Google Scholar] [CrossRef]
- Shah, M.; Ullah, M.A.; Drouet, S.; Younas, M.; Tungmunnithum, D.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Interactive Effects of Light and Melatonin on Biosynthesis of Silymarin and Anti-Inflammatory Potential in Callus Cultures of Silybum marianum (L.) Gaertn. Molecules. 2019, 24, 1207. [Google Scholar] [CrossRef]
- Ullah, M.A.; Tungmunnithum, D.; Garros, L.; Drouet, S.; Hano, C.; Abbasi, B.H. Effect of Ultraviolet-C Radiation and Melatonin Stress on Biosynthesis of Antioxidant and Antidiabetic Metabolites Produced in In Vitro Callus Cultures of Lepidium sativum L. Int. J. Mol. Sci. 2019, 20, 1787. [Google Scholar] [CrossRef]
- Gai, Q.-Y.; Jiao, J.; Luo, M.; Wang, W.; Zhao, C.-J.; Fu, Y.-J.; Ma, W. UV elicitation for promoting astragaloside production in Astragalus membranaceus hairy root cultures with transcriptional expression of biosynthetic genes. Ind. Crop. Prod. 2016, 84, 350–357. [Google Scholar] [CrossRef]
- Nawkar, G.; Maibam, P.; Park, J.; Sahi, V.; Lee, S.; Kang, C. UV-induced cell death in plants. Int. J. Mol. Sci. 2013, 14, 1608–1628. [Google Scholar] [CrossRef]
- Xu, A.; Zhan, J.-C.; Huang, W.-D. Effects of ultraviolet C, methyl jasmonate and salicylic acid, alone or in combination, on stilbene biosynthesis in cell suspension cultures of Vitis vinifera L. cv. Cabernet Sauvignon. Plant Cell Tissue Organ Cult. 2015, 122, 197–211. [Google Scholar] [CrossRef]
- Xu, A.; Zhan, J.-C.; Huang, W.-D. Combined elicitation of chitosan and ultraviolet C enhanced stilbene production and expression of chitinase and β-1, 3-glucanase in Vitis vinifera cell suspension cultures. Plant Cell Tissue Organ Cult. 2016, 124, 105–117. [Google Scholar] [CrossRef]
- Anjum, S.; Abbasi, B.H.; Doussot, J.; Favre-Reguillon, A.; Hano, C. Effects of photoperiod regimes and ultraviolet-C radiations on biosynthesis of industrially important lignans and neolignans in cell cultures of Linum usitatissimum L. (Flax). J. Photochem. Photobiol. B: Biol. 2017, 167, 216–227. [Google Scholar] [CrossRef]
- Khan, T.; Ullah, M.A.; Garros, L.; Hano, C.; Abbasi, B.H. Synergistic effects of melatonin and distinct spectral lights for enhanced production of anti-cancerous compounds in callus cultures of Fagonia indica. J. Photochem. Photobiol. B: Biol. 2018, 190, 163–171. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N.; Ali, M. Exogenous melatonin trigger biomass accumulation and production of stress enzymes during callogenesis in medicinally important Prunella vulgaris L. (Selfheal). Physiol. Mol. Biol. Plants. 2018, 24, 1307–1315. [Google Scholar] [CrossRef]
- Sarropoulou, V.N.; Therios, I.N.; Dimassi-Theriou, K.N. Melatonin promotes adventitious root regeneration in in vitro shoot tip explants of the commercial sweet cherry rootstocks CAB-6P (Prunus cerasus L.), Gisela 6 (P. cerasus× P. canescens), and MxM 60 (P. avium× P. Mahaleb). J. Pineal Res. 2012, 52, 38–46. [Google Scholar] [CrossRef]
- Guidi, L.; Brunetti, C.; Fini, A.; Agati, G.; Ferrini, F.; Gori, A.; Tattini, M. UV radiation promotes flavonoid biosynthesis, while negatively affecting the biosynthesis and the de-epoxidation of xanthophylls: Consequence for photoprotection? Environ. Exp. Bot. 2016, 127, 14–25. [Google Scholar] [CrossRef]
- Escobar, A.L.; de Oliveira Silva, F.M.; Acevedo, P.; Nunes-Nesi, A.; Alberdi, M.; Reyes-Díaz, M. Different levels of UV-B resistance in Vaccinium corymbosum cultivars reveal distinct backgrounds of phenylpropanoid metabolites. Plant Physiol. Biochem. 2017, 118, 541–550. [Google Scholar] [CrossRef]
- Nigro, F.; Ippolito, A.; Lattanzio, V.; Di Venere, D.; Salerno, M. Effect of ultraviolet-C light on postharvest decay of strawberry. J. Plant Pathol. 2000, 29–37. [Google Scholar]
- Tiecher, A.; de Paula, L.A.; Chaves, F.C.; Rombaldi, C.V. UV-C effect on ethylene, polyamines and the regulation of tomato fruit ripening. Postharvest Biol. Technol. 2013, 86, 230–239. [Google Scholar] [CrossRef]
- Chen, Y.E.; Mao, J.J.; Sun, L.Q.; Huang, B.; Ding, C.B.; Gu, Y.; Liao, J.Q.; Hu, C.; Zhang, Z.W.; Yuan, S. Exogenous melatonin enhances salt stress tolerance in maize seedlings by improving antioxidant and photosynthetic capacity. Physiol. Plant. 2018, 164, 349–363. [Google Scholar] [CrossRef]
- Liang, D.; Shen, Y.; Ni, Z.; Wang, Q.; Lei, Z.; Xu, N.; Deng, Q.; Lin, L.; Wang, J.; Lv, X. Exogenous melatonin application delays senescence of kiwifruit leaves by regulating the antioxidant capacity and biosynthesis of flavonoids. Front. Plant Sci. 2018, 9, 426. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Awadi, M.E. Alleviation of salinity stress on Vicia faba L. plants via seed priming with melatonin. Acta Biologica Colomb. 2015, 20, 223–235. [Google Scholar] [CrossRef]
- Szafrańska, K.; Glinska, S.; Janas, K.M. Changes in the nature of phenolic deposits after re-warming as a result of melatonin pre-sowing treatment of Vigna radiata seeds. J. Plant Physiol. 2012, 169, 34–40. [Google Scholar] [CrossRef]
- Rea, G.; Antonacci, A.; Lambreva, M.; Pastorelli, S.; Tibuzzi, A.; Ferrari, S.; Fischer, D.; Johanningmeier, U.; Oleszek, W.; Doroszewska, T. Integrated plant biotechnologies applied to safer and healthier food production: The Nutra-Snack manufacturing chain. Trends Food Sci. Technol. 2011, 22, 353–366. [Google Scholar] [CrossRef]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation: A tool for enriching the bioactive composition of foods. Molecules. 2014, 19, 13541–13563. [Google Scholar] [CrossRef]
- Muhitch, M.J.; Fletcher, J.S. Influence of culture age and spermidine treatment on the accumulation of phenolic compounds in suspension cultures. Plant Physiol. 1985, 78, 25–28. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell. 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Chappel, J.; Hahlbrock, K. Transcription of plant defence genes in response to UV light or fungal elicitor. Nature 1984, 311, 76–78. [Google Scholar] [CrossRef]
- Lavola, A. Accumulation of flavonoids and related compounds in birch induced by UV-B irradiance. Tree Physiol. 1998, 18, 53–58. [Google Scholar] [CrossRef]
- Duran, R.E.; Kilic, S.; Coskun, Y. Melatonin influence on in vitro callus induction and phenolic compound production in sweet basil (Ocimum basilicum L.). In Vitro Cell. Dev. Biol. Plant. 2019, 55, 468–475. [Google Scholar] [CrossRef]
- Guo, J.; Han, W.; Wang, M. Ultraviolet and environmental stresses involved in the induction and regulation of anthocyanin biosynthesis: A review. Afr. J. Biotechnol. 2008, 7, 25. [Google Scholar]
- Khan, T.; Abbasi, B.H.; Afridi, M.S.; Tanveer, F.; Ullah, I.; Bashir, S.; Hano, C. Melatonin-enhanced biosynthesis of antimicrobial AgNPs by improving the phytochemical reducing potential of a callus culture of Ocimum basilicum L. var. thyrsiflora. RSC advances. 2017, 7, 38699–38713. [Google Scholar]
- Christie, J.M.; Jenkins, G.I. Distinct UV-B and UV-A/blue light signal transduction pathways induce chalcone synthase gene expression in Arabidopsis cells. Plant Cell. 1996, 8, 1555–1567. [Google Scholar]
- Lavola, A.; Aphalo, P.J.; Lahti, M.; Julkunen-Tiitto, R. Nutrient availability and the effect of increasing UV-B radiation on secondary plant compounds in Scots pine. Environ. Exp. Bot. 2003, 49, 49–60. [Google Scholar] [CrossRef]
- Kataoka, I.; Sugiyama, A.; Beppu, K. Role of ultraviolet radiation in accumulation of anthocyanin in berries of’Gros Colman’grapes (Vitis vinifera L.). J. Jpn. Soc. Hortic. Sci. 2003, 72, 1–6. [Google Scholar] [CrossRef]
- Hirner, A.A.; Seitz, H.U. Isoforms of chalcone synthase in Daucus carota L. and their differential expression in organs from the European wild carrot and in ultraviolet-A-irradiated cell cultures. Planta. 2000, 210, 993–998. [Google Scholar] [CrossRef]
- Toguri, T.; Umemoto, N.; Kobayashi, O.; Ohtani, T. Activation of anthocyanin synthesis genes by white light in eggplant hypocotyl tissues, and identification of an inducible P-450 cDNA. Plant Mol. Biol. 1993, 23, 933–946. [Google Scholar] [CrossRef]
- Riley, P.A. Free radicals in biology: Oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Biol. 1994, 65, 27–33. [Google Scholar] [CrossRef]
- Leon, J.; Acuna-Castroviejo, D.; Escames, G.; Tan, D.X.; Reiter, R.J. Melatonin mitigates mitochondrial malfunction. J. Pineal Res. 2005, 38, 1–9. [Google Scholar] [CrossRef]
- Martin, M.; Macias, M.; Leon, J.; Escames, G.; Khaldy, H.; Acuna-Castroviejo, D. Melatonin increases the activity of the oxidative phosphorylation enzymes and the production of ATP in rat brain and liver mitochondria. Int. J. Biochem. Cell Biol. 2002, 34, 348–357. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules. 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Ali, M.B.; Khatun, S.; Hahn, E.-J.; Paek, K.-Y. Enhancement of phenylpropanoid enzymes and lignin in Phalaenopsis orchid and their influence on plant acclimatisation at different levels of photosynthetic photon flux. Plant Growth Regul. 2006, 49, 137–146. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Eichholz, I.; Huyskens-Keil, S.; Keller, A.; Ulrich, D.; Kroh, L.W.; Rohn, S. UV-B-induced changes of volatile metabolites and phenolic compounds in blueberries (Vaccinium corymbosum L.). Food Chem. 2011, 126, 60–64. [Google Scholar] [CrossRef]
- Li, C.; Wang, P.; Wei, Z.; Liang, D.; Liu, C.; Yin, L.; Jia, D.; Fu, M.; Ma, F. The mitigation efects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to stress combination in tomato plants: New insights in the protective role of melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef]
- Grzegorczyk, I.; Bilichowski, I.; Mikiciuk-Olasik, E.; Wysokinska, H. In vitro cultures of Salvia officinalis L. as a source of antioxidant compounds. Acta Societatis Botanicorum Poloniae 2005, 74, 1. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef]
- Ul-Haq, I.; Ullah, N.; Bibi, G.; Kanwal, S.; Ahmad, M.S.; Mirza, B. Antioxidant and cytotoxic activities and phytochemical analysis of Euphorbia wallichii root extract and its fractions. Iran. J. Pharm. Res. 2012, 11, 241. [Google Scholar]
- Velioglu, Y.; Mazza, G.; Gao, L.; Oomah, B. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Khan, M.; Guo, B.; Bokhari, S.A.; Khan, M.A. Efficient regeneration and antioxidative enzyme activities in Brassica rapa var. turnip. Plant Cell Tissue Organ Cult. 2011, 105, 337–344. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
Sample Availability: Plant materials and chemical standards used in the present study are available upon request to corresponding authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
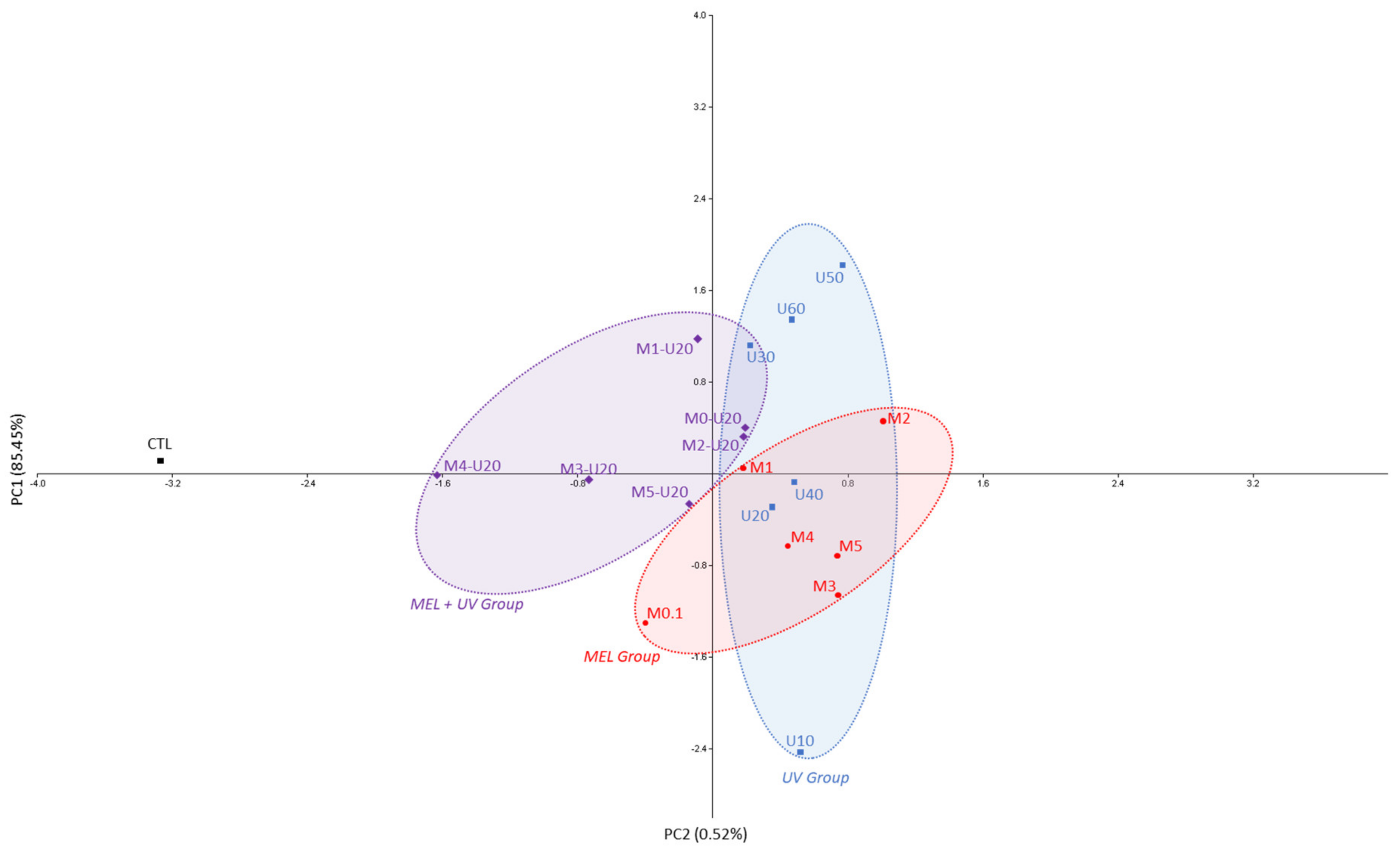{kind=link}
| Treatment | Conc. | Phenylpropanoid Compounds (mg/g DW) | ||||
|---|---|---|---|---|---|---|
| Rosmarinic Acid | Chicoric Acid | Caffeic Acid | Cynadin | Peonidin | ||
| Control | NAA | 56.30 ± 3.85 d,e | 12.33 ± 1.05 e | 0.28 ± 0.08 d | 0.12 ± 0.006 e | 0.11 ± 0.001 e |
| Melatonin (mg/L) | 0.1 | 62.27 ± 1.94 d,e | 32.63 ± 1.41 c | 0.52 ± 0.01 a | 0.36 ± 0.07 c | 0.18 ± 0.004 c |
| 1 | 79.49 ± 2.52 c,d | 37.31 ± 1.89 bc | 0.47 ± 0.03 ab | 0.44 ± 0.04 ab | 0.19 ± 0.006 c | |
| 2 | 77.68 ± 3.04 c,d | 39.99 ± 0.55 b | 0.41 ± 0.06 bc | 0.45 ± 0.009 ab | 0.22 ± 0.002 b,c | |
| 3 | 74.10 ± 4.23 c,d | 36.98 ± 0.34 b,c | 0.46 ± 0.07 b | 0.41 ± 0.06 b | 0.20 ± 0.001 c | |
| 4 | 65.95 ± 3.14 d,e | 36.18 ± 1.14 b,c | 0.45 ± 0.04 b | 0.40 ± 0.02 b | 0.21 ± 0.003 c | |
| 5 | 78.25 ± 2.42 c,d | 37.75 ± 1.31 b,c | 0.44 ± 0.02 b | 0.42 ± 0.08 b | 0.21 ± 0.007 c | |
| UV-C (min) | 10 | 134.01 ± 3.95 a | 37.01 ± 1.19 b,c | 0.44 ± 0.05 b | 0.42 ± 0.09 b | 0.21 ± 0.001 b,c |
| 20 | 77.81 ± 2.77 c,d | 40.65 ± 0.88 b | 0.41 ± 0.03 b,c | 0.40 ± 0.01 b | 0.24 ± 0.009 b | |
| 30 | 82.32 ± 2.11 c,d | 43.34 ± 0.97 a,b | 0.36 ± 0.06 c | 0.49 ± 0.03 a | 0.25 ± 0.007 b | |
| 40 | 89.24 ± 4.62 c | 43.13 ± 1.05 a,b | 0.32 ± 0.02 c,d | 0.50 ± 0.06 a | 0.25 ± 0.003 b | |
| 50 | 110.8 ± 6.39 b | 51.52 ± 2.57 a | 0.29 ± 0.08 c,d | 0.50 ± 0.02 a | 0.30 ± 0.005 a | |
| 60 | 97.61 ± 1.48 b,c | 39.77 ± 0.78 b | 0.041 ± 0.01 b,c | 0.45 ± 0.08 ab | 0.23 ± 0.002 b,c | |
| Melatonin + UV-C | 0.1 + 20 | 74.94 ± 2.35 c,d | 41.40 ± 0.72 b | 0.42 ± 0.01 b,c | 0.46 ± 0.04 a,b | 0.24 ± 0.005 b |
| 1 + 20 | 86.26 ± 1.43 b,c | 42.96 ± 0.79 b | 0.40 ± 0.05 b,c | 0.43 ± 0.01 a,b | 0.23 ± 0.002 b,c | |
| 2 + 20 | 75.65 ± 2.55 c,d | 39.99 ± 1.58 b | 0.41 ± 0.08 b,c | 0.44 ± 0.03 a,b | 0.24 ± 0.008 b | |
| 3 + 20 | 53.93 ± 3.28 d,e | 30.88 ± 1.07 c | 0.50 ± 0.01 a,b | 0.36 ± 0.05 c | 0.18 ± 0.004 c | |
| 4 + 20 | 49.56 ± 2.82 e | 23.67 ± 0.98 d | 0.54 ± 0.07 a | 0.28 ± 0.04 d | 0.14 ± 0.006 e | |
| 5 + 20 | 68.49 ± 6.03 d | 33.76 ± 1.43 bc | 0.47 ± 0.09 a,b | 0.38 ± 0.07 c | 0.19 ± 0.005 c | |
| Variables | Ros-A | Chi-A | Caff-A | Cyan | Peon | DPPH | ABTS | FRAP | cRO/NS |
|---|---|---|---|---|---|---|---|---|---|
| Ros-A | |||||||||
| Chi-A | 0.613 ** | ||||||||
| Caff-A | −0.412 ns | −0.203 ns | |||||||
| Cyan | 0.575 ** | 0.959 *** | −0.152 ns | ||||||
| Peon | 0.622 ** | 0.965 *** | −0.331 ns | 0.895 *** | |||||
| DPPH | 0.669 ** | 0.792 *** | −0.337 ns | 0.760 *** | 0.792 *** | ||||
| ABTS | 0.546 * | 0.873 *** | −0.047 ns | 0.908 *** | 0.780 *** | 0.686 ** | |||
| FRAP | −0.546 * | −0.352 ns | 0.302 ns | −0.253 ns | −0.437 ns | −0.425 ns | −0.130 ns | ||
| cRO/NS | 0.626 ** | 0.870 *** | −0.545 * | 0.799 *** | 0.896 *** | 0.767 *** | 0.681 ** | −0.554 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazir, M.; Asad Ullah, M.; Mumtaz, S.; Siddiquah, A.; Shah, M.; Drouet, S.; Hano, C.; Abbasi, B.H. Interactive Effect of Melatonin and UV-C on Phenylpropanoid Metabolite Production and Antioxidant Potential in Callus Cultures of Purple Basil (Ocimum basilicum L. var purpurascens). Molecules 2020, 25, 1072. https://doi.org/10.3390/molecules25051072
Nazir M, Asad Ullah M, Mumtaz S, Siddiquah A, Shah M, Drouet S, Hano C, Abbasi BH. Interactive Effect of Melatonin and UV-C on Phenylpropanoid Metabolite Production and Antioxidant Potential in Callus Cultures of Purple Basil (Ocimum basilicum L. var purpurascens). Molecules. 2020; 25(5):1072. https://doi.org/10.3390/molecules25051072
Chicago/Turabian StyleNazir, Munazza, Muhammad Asad Ullah, Sadia Mumtaz, Aisha Siddiquah, Muzamil Shah, Samantha Drouet, Christophe Hano, and Bilal Haider Abbasi. 2020. "Interactive Effect of Melatonin and UV-C on Phenylpropanoid Metabolite Production and Antioxidant Potential in Callus Cultures of Purple Basil (Ocimum basilicum L. var purpurascens)" Molecules 25, no. 5: 1072. https://doi.org/10.3390/molecules25051072
APA StyleNazir, M., Asad Ullah, M., Mumtaz, S., Siddiquah, A., Shah, M., Drouet, S., Hano, C., & Abbasi, B. H. (2020). Interactive Effect of Melatonin and UV-C on Phenylpropanoid Metabolite Production and Antioxidant Potential in Callus Cultures of Purple Basil (Ocimum basilicum L. var purpurascens). Molecules, 25(5), 1072. https://doi.org/10.3390/molecules25051072