1. Introduction
Over the past decades, drugs derived from plants have continued to have beneficial health effects in the prevention and treatment of many diseases. Natural products and plants have made an enormous impact on the discovery of anticancer drugs. Approximately 60% of all cancer drugs that are used clinically are either natural products, analogues of the parent natural products, or mimics [
1]. Although plants have been identified as a sound source of anticancer agents, the promise of combinational chemistry and modern synthetic technologies has overshadowed natural product research as a source of new drugs [
2]. Plants historically stand at the core of medicine, and they are still a major source of prospective new drug leads. Although plants represent a superb source of the most effective anticancer drugs, such as vinca alkaloids, taxanes, podophyllotoxin derivatives and others, a large number of plant-derived compounds have barely been studied and still need to be investigated [
3].
The Berberidaceae family is a rich source of isoquinoline alkaloids of various structural types, including bisbenzylisoquinoline alkaloids, which are characterized by having the benzylisoquinoline skeleton dimerized by ether linkages or carbon-carbon bonds. Their diversity is due to the different substituents on the aromatic rings. They are biosynthetically obtained from benzyltetrahydroisoquinoline units through phenolic oxidative coupling [
4]. The representative of this structural type is alkaloid berbamine from
Berberis vulgaris [
5]. Since berbamine showed exquisite potency against cancer cell lines, there is a dire need to investigate and bring to light the evidence for possible anticancer activity of hitherto unexplored novel bisbenzylisoquinoline alkaloid bersavine (
Figure 1).
Bersavine, a natural bisbenzylisoquinoline alkaloid, has been recently isolated for the first time from the root bark
Berberis vulgaris L. (Berberidaceae) [
5]. Bersavine, in structural analogy with berbamine, incorporates two tetrahydrobenzylisoquinoline moieties combined through two ether linkages. Berbamine has been known for its promising cytotoxic activity on human cancer cell lines in vitro for a long time, and it has also been shown to stimulate normal hematopoiesis and enhance immune function in cancer patients [
6]. Berbamine also exerts anti-inflammatory effects by inhibiting nuclear factor-kappaB (NF-κB) and mitogen-activated protein kinase (MAPK) signalling pathways [
7]. Its ability to inhibit proliferation in cancer cells was described repetitively in many experimental studies [
8,
9,
10,
11,
12,
13]. Zhang and coworkers observed the apoptosis-inducing effect of berbamine in colorectal cancer cells by activating p53-dependent apoptotic signalling pathways [
14]. Additionally, another recent study showed enhanced antimetastatic and antitumorigenic efficacy of berbamine-loaded lipid nanoparticles in vivo [
15]. In the study of Jia et al., berbamine and paclitaxel were tested for their synergistic antitumor effects via the reactive oxygen species ROS/Akt pathway in glioma cells; berbamine has been shown to be a promising adjuvant to conventional chemotherapy of malignant glioma [
7].
While berbamine has been shown to possess multiple biological activities, only one study on bersavine bioactivity has been published so far. Hošťálková et al. described the neuroprotective activity of bersavine, including the capability to inhibit human erythrocyte acetylcholinesterase (
hAChE), human serum butyrylcholinesterase (
hBuChE) and prolyl oligopeptidase (POP; E.C. 3.4.21.26) activities [
5]. However, as far as we know, no evidence supporting the antiproliferative, cytotoxic and proapoptotic activity of bersavine has yet been described. In this study, we aimed to elucidate the antiproliferative and cytotoxic activity of bersavine in mini panels with cancer cells. To comprehensively reveal the anticancer potential of bersavine, the evidence for an apoptosis-mediated cell death, impairment in cell cycle progression, including the underpinning molecular mechanisms involved, were also investigated in human leukemic cells.
3. Discussion
This work is the first report focused on exploring the antiproliferative, cytotoxic and apoptosis-inducing effects of bersavine. In previous studies, the structurally related alkaloid berbamine was clearly confirmed as a promising anticancer drug candidate, which was supported by experimental evidence of strong antiproliferative, cytotoxic and proapoptotic activity [
6,
7,
8,
12]. Moreover, berbamine is well-known in Chinese medicine and it has been used as a treatment option for decades [
11]. On the other hand, nothing so far is known about the antiproliferative, cytotoxic and apoptosis-inducing effect of bersavine.
In the study of Wang et al., 24 h of treatment with berbamine negatively affected HepG2 hepatocellular cancer cells, resulting in a decrease in cell viability and proliferation in a dose-dependent manner, with an IC
50 value of 34.5 ± 0.5 µM [
11]. The higher impact of berbamine treatment was observed in work oriented on glioma cancer, where the U-87 human glioblastoma cell line was used as an experimental model. By analysing U-87 glioblastoma cell proliferation at a dosing interval of 48 h, it was observed that berbamine inhibited the growth of cancer cells in a dose-dependent manner with an IC
50 value of 9.8 ± 0.6 µM [
7]. Berbamine-treated lung carcinoma cells A549 showed a dose-dependent decrease of viability after 48 h of treatment, with an estimated IC
50 value of 34.63 ± 1.12 µM [
15]. Another recent study by Zhang et al. described the suppression of proliferation of colorectal cancer cells HCT116 and SW480 by a 48 h treatment with berbamine. The IC
50 values determined were 12.3 ± 1.02 µM for the HCT116 cell line and 16.4 ± 0.89 µM for the SW480 cell line, respectively [
14]. For the first time, our study showed that a new
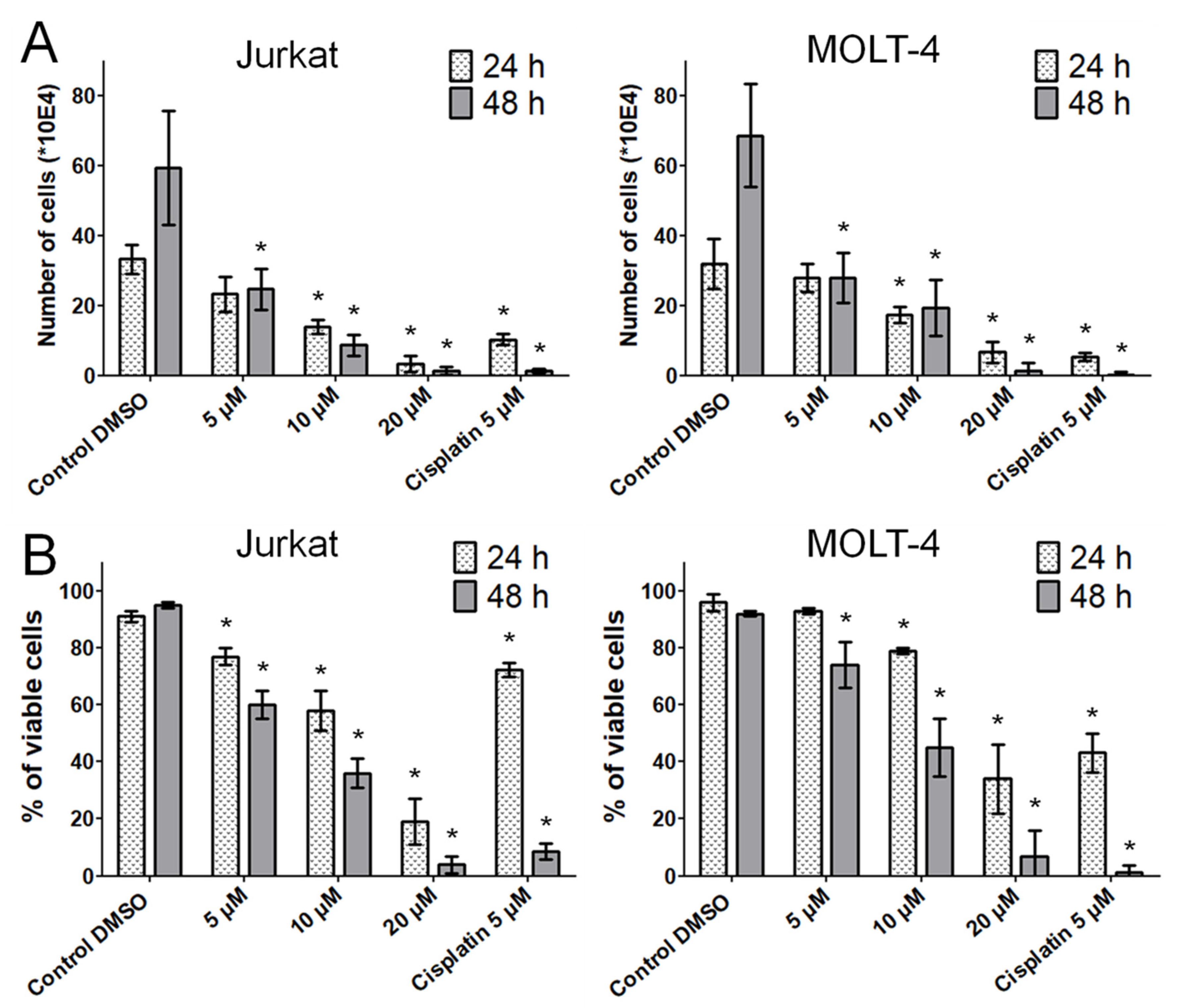
Berberis vulgaris active constituent bersavine applied for 48 h at 5 μM inhibited both the proliferation and the viability of human leukemic Jurkat and MOLT-4 cells in vitro. The coassayed IC
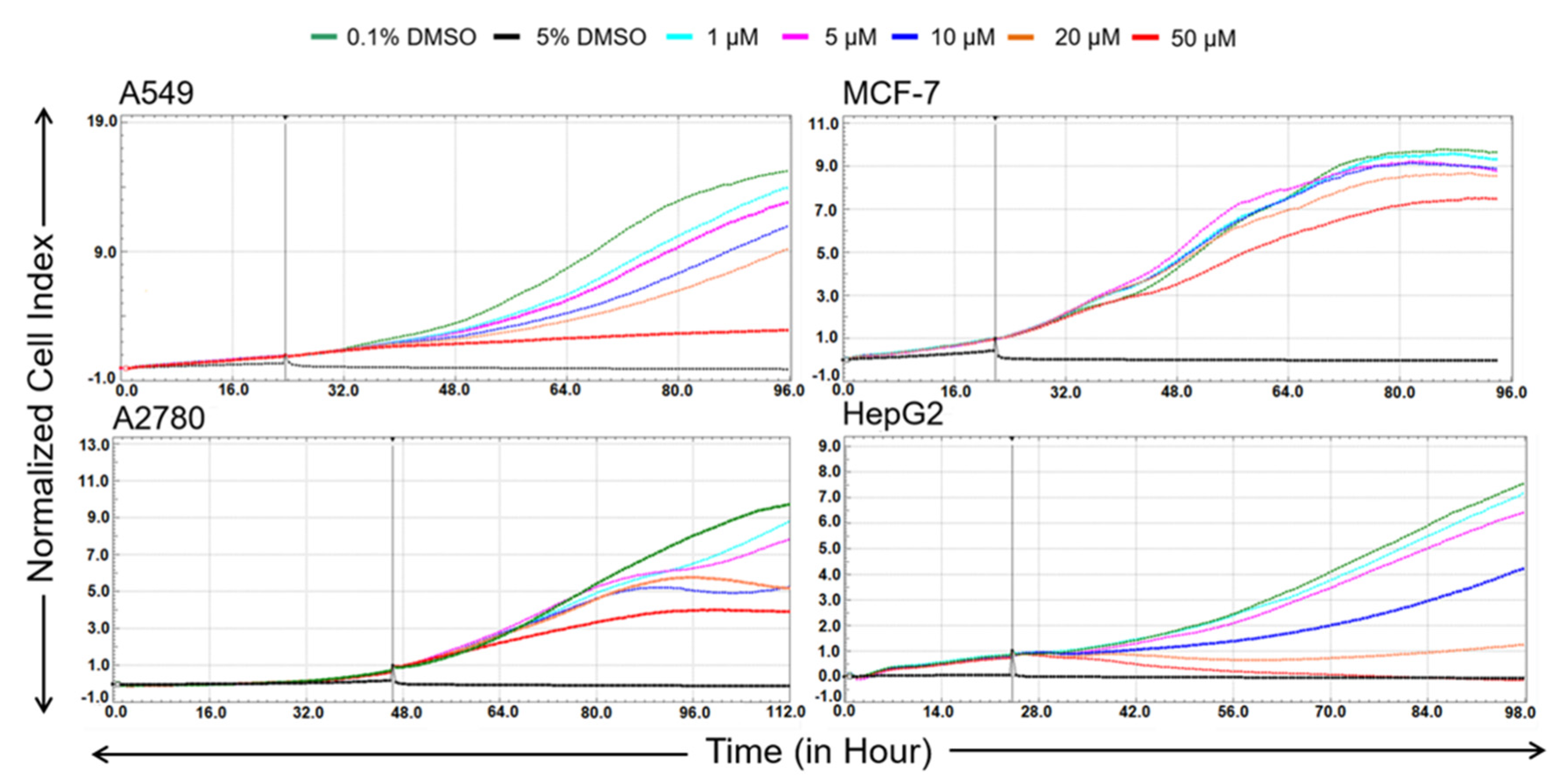
50 values of bersavine for the most sensitive human cancer cells in the initial cytotoxicity screen ranged between 8.1 ± 1.7 and 11.0 ± 1.2 µM. In addition, a real-time cell adhesion, proliferation and cytotoxicity xCELLigence system confirmed the high antiproliferative activity of bersavine against cancer cell lines derived from a lung (A549), ovarian (A2780), breast (MCF-7) and hepatocellular (HepG2) cancer during 72 h of continuous monitoring. These results suggest that bersavine has a similar cytotoxic and antiproliferative effect to berbamine, as has been described in the aforementioned studies.
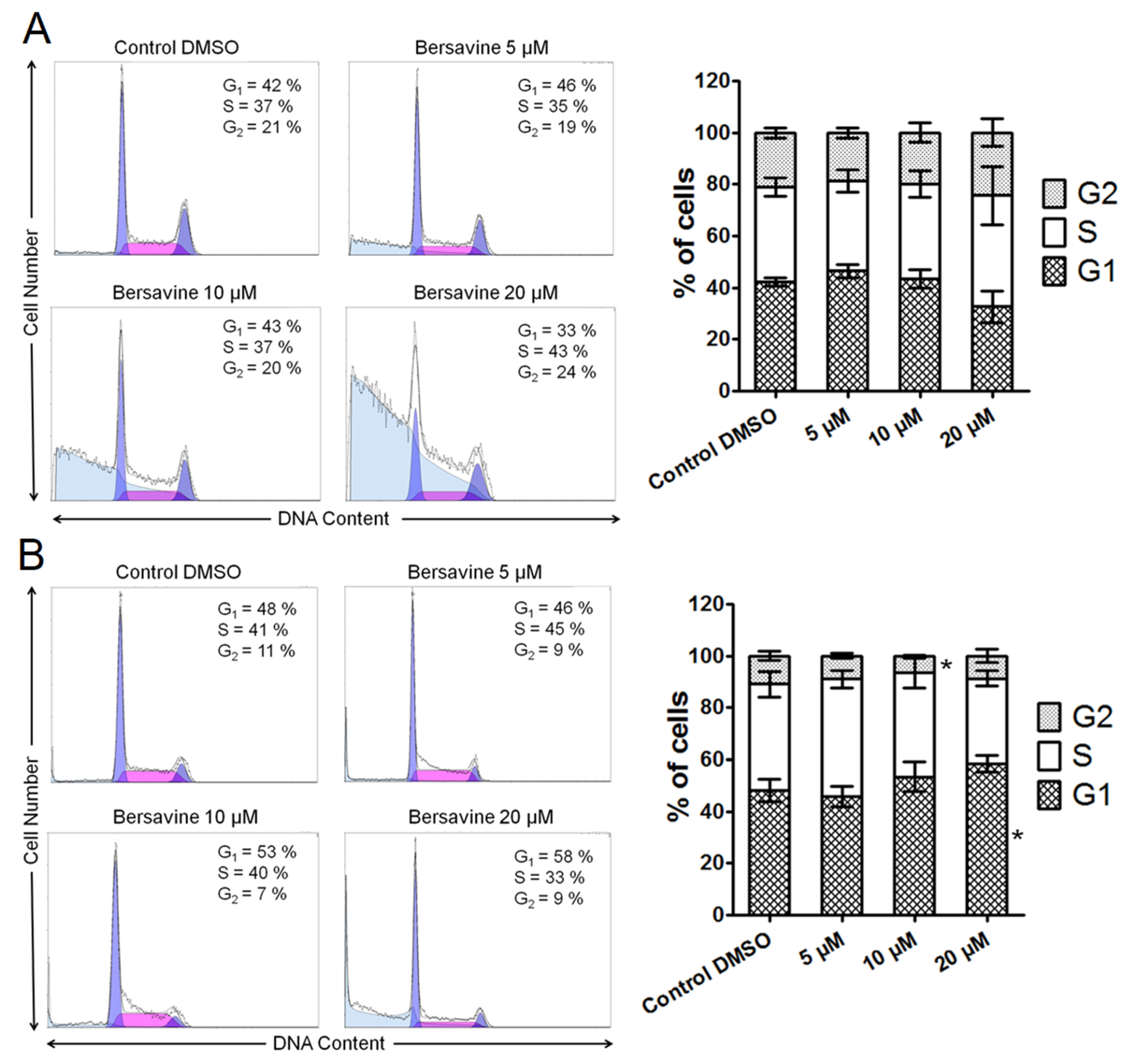
Mechanism studies showed that berbamine exerts its cytostatic and cytotoxic potency against cancer cells by perturbing the cell cycle and inducing apoptosis. Since the chemical structure of berbamine is closely related to bersavine, it is interesting to compare these observations to our results. According to Zhang et al., berbamine treatment at 20 µg/mL (≈33 µM) for 48 h not only significantly increased the percentage of apoptotic cells in both HCT116 and SW480 cells, but also significantly increased the cell population at the G0/G1 phase and decreased the cell population at the G2/M phase in both cell lines [
14]. Also, bersavine in our experiments showed similar effects on MOLT-4 cells because the concentration of 10 µM of bersavine significantly decreased the percentage of leukemic cells in the G2/M phase and 20 µM of treatment significantly increased the percentage of cells in the G1 phase. In another study, berbamine applied to SMMC7721 hepatocellular carcinoma cells caused cell cycle arrest in the G0/G1 phase, induced a loss of mitochondrial membrane potential (Δ
ψm) and induced the activation of caspase-3 and caspase-9 [
16]. Moreover, the authors provided experimental evidence for the involvement of caspases in apoptosis triggered by berbamine, since the berbamine-induced apoptosis could be blocked by pretreatment with the broad caspase inhibitor Z-VAD-FMK [
16]. Berbamine also effectively inhibits the growth of RPMI 8226 human multiple myeloma cells, which is associated with the activation of the GADD45/JNK signalling pathway, apoptosis and upregulation of p53, p21 and GADD45gamma mRNA [
17]. On the other hand, Wang et al. described the proapoptotic effects of berbamine, which upregulated the FasL expression in HepG2 cells, activated caspase-8, induced a loss of mitochondrial membrane potential (Δ
ψm), with subsequent activation of caspase-9 and -3, which ultimately resulted in the disassembly of HepG2 cells [
11]. This study suggests that berbamine induces apoptosis in HepG2 cells through a type II Fas signalling pathway [
11].
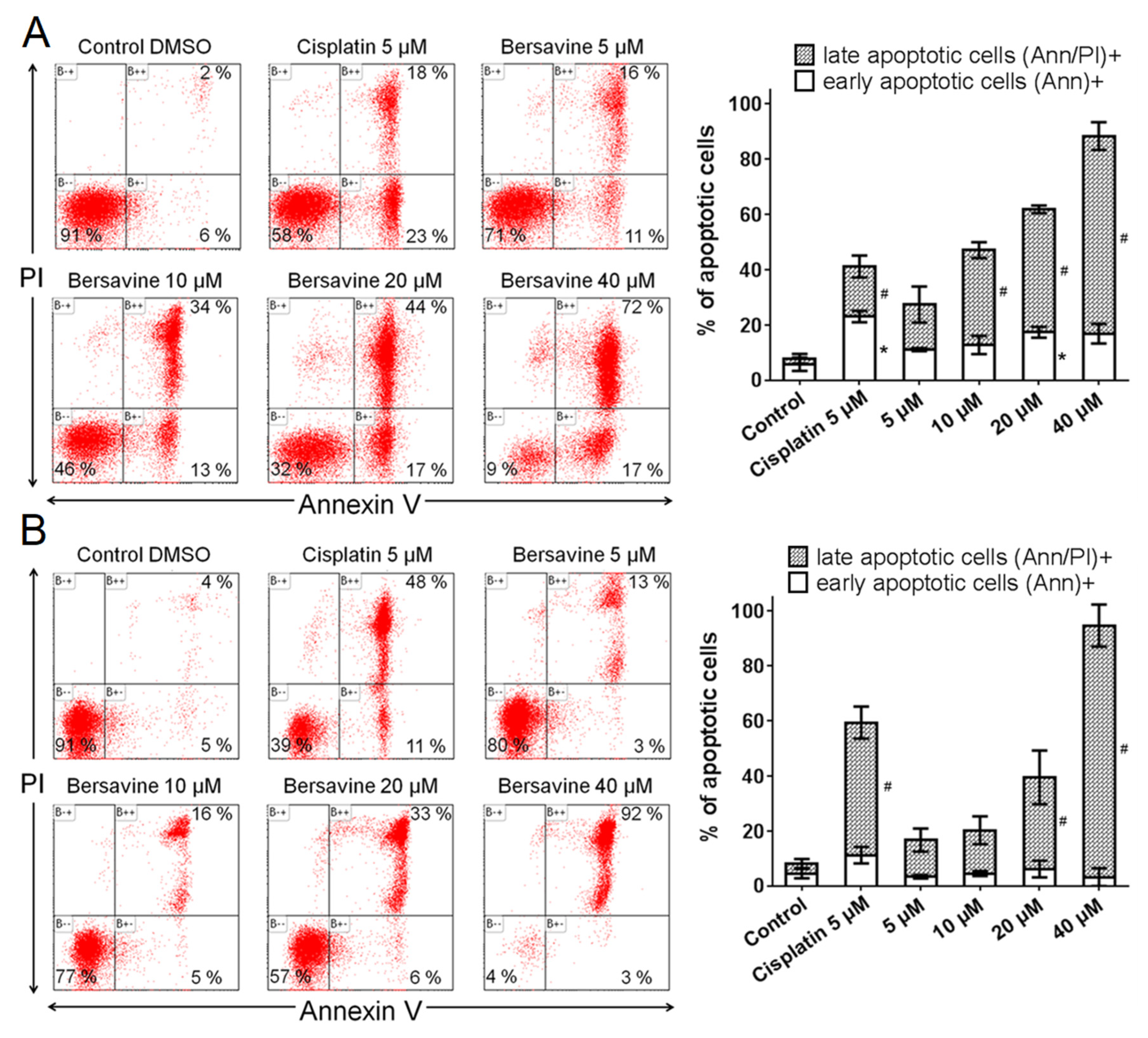
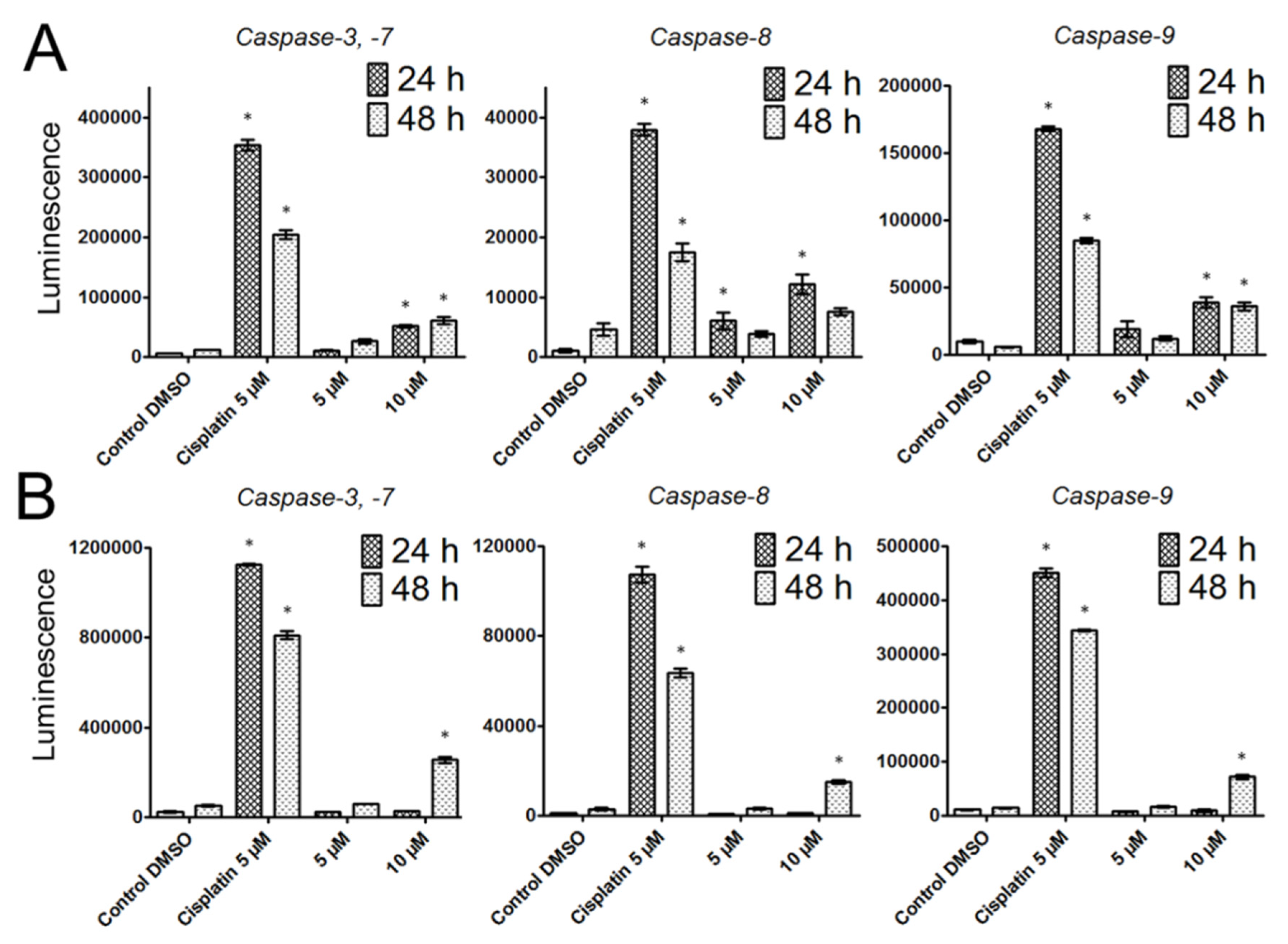
Since previous studies have provided clear evidence that berbamine treatment increased apoptotic rates, we decided to ascertain whether apoptosis also occurs after bersavine treatment. Exposure to bersavine resulted in increased levels of caspase-3/-7, caspase-8 and caspase-9 in Jurkat cells, which was generally more pronounced with a higher concentration of bersavine and in the early period of 24 h of treatment. The apoptotic effect of bersavine in Jurkat cells was considerably higher after 24 h of bersavine treatment by determining caspase activity than in MOLT-4 cells. The same bersavine treatment period caused a higher increase in the percentage of early apoptotic Annexin-V single-positive Jurkat cells assayed by flow cytometry compared to those seen in the MOLT-4 cells. The lack of evidence for increased caspase activity after 24 h of bersavine treatment in MOLT-4 cells can be attributed largely to the pronounced cell cycle arrest of MOLT-4 cells in the G1/S phase at this short incubation time period. Furthermore, considering the increased level of Chk1 phosphorylated at Ser345 and the decreased level of Rb phosphorylated at Ser807/811 after 24 h of bersavine (10 µM) treatment and the significantly increased levels of caspase-3/-7, -8 and -9 after 48 h of treatment, we hypothesise that the early treatment interval action (24 h) of bersavine on MOLT-4 cells primarily lies in arresting the cell-cycle progression rather than apoptosis induction. Our observations also indicate that the proapoptotic effect of bersavine is at least partially mediated via the upregulation of the protein levels of phosphorylated p53 at Ser392 in the case of p53-wild-type (p53+) MOLT-4 cells, similar to berbamine, as was described in work of Zhang et al. [
14].
4. Materials and Methods
4.1. Cell Culture and Culture Conditions
The selected human tumour cell lines Jurkat (acute T cell leukemia), MOLT-4 (acute lymphoblastic leukaemia), A549 (lung carcinoma), HT-29 (colorectal adenocarcinoma), PANC-1 (pancreas epithelioid carcinoma), A2780 (ovarian carcinoma), HeLa (cervix adenocarcinoma), MCF-7 (breast adenocarcinoma), SAOS-2 (osteosarcoma) and HepG2 (hepatocellular carcinoma) were purchased from either ATCC (Manassas, VA, USA) or Sigma-Aldrich (St. Louis, MO, USA) and were cultured in accordance with the provider´s culture method guidelines. The cell cultures were maintained under standard cell culture conditions at 37 °C in a humidified incubator in an atmosphere of 5% CO2, 95% air. Cells were passaged every 2–3 days to obtain exponential growth. Cells in the maximum range of 20 passages and in an exponential growth phase were used for this study.
4.2. Cell Treatment
Bersavine and berbamine—fresh stock solutions of bersavine and berbamine in concentrations of 50 mM were dissolved in dimethyl sulfoxide (DMSO) (Sigma-Aldrich, St. Louis, MO, USA). Stock solutions were freshly prepared before use in the experiments. Bersavine and berbamine were isolated from an alkaloidal extract of the root bark of
Berberis vulgaris (Berberidaceae). The detailed isolation and identification of the structures (1D-, 2D-NMR, HRMS and optical rotation) of the alkaloid has been published by Hostalkova and colleagues [
5]. For the experiments, the stock solutions were diluted using the complete culture medium to create final concentrations of 1–50 μM, making sure the concentration of DMSO was <0.1% to avoid any toxic effects on the cells. Cisplatin and doxorubicin were purchased from Sigma-Aldrich (Sigma-Aldrich, St. Louis, MO, USA). Control cells were sham-treated with a DMSO vehicle only (0.1%; control). Cells treated with 5% DMSO, cisplatin at 5 µM or doxorubicin at 1 µM were used as a positive control.
4.3. WST-1 Assay and Growth Percent Calculation
To determine cell proliferation, cell viability, and the cytotoxicity of cells treated with bersavine and berbamine at a single dose of 10 µM or in a broad concentration range of 0.1–100 µM (determination of IC50 values), we used a standard colorimetric method measuring a WST-1 tetrazolium salt reduction via mitochondrial dehydrogenase activity. At the onset of the experiments, each cell line was seeded at a previously established optimal density (1 × 103 to 50 × 103 cells per well) in a 96-well plate (TPP, Trasadingen, Switzerland) and the cells were allowed to settle overnight. The cells were treated for 48 h with bersavine. Doxorubicin (Sigma-Aldrich, St. Louis, MO, USA) at a concentration of 1 µM was used as a positive control. At the end of the cultivation period, a WST-1 proliferation and viability assay (Roche, Basel, Switzerland) was performed in accordance with the manufacturer’s instructions. Absorbance was measured using a Tecan Infinite M200 (Tecan, Männedorf, Switzerland) at 440 nm. Each value is the mean of three independent experiments and represents the percentage of proliferation/viability of control, nontreated cells (100%). The growth percent (GP) value was calculated for alkaloid tested. GP represents the mean of the proliferation/viability decrease as a percentage in regard to all of the 9 cell lines treated with bersavine or berbamine.
4.4. Screening for Antiproliferative Activity Using the xCELLigence System
The xCELLigence system (Roche, Basel, Switzerland and ACEA Biosciences, San Diego, CA, USA) was used to monitor cell adhesion, proliferation and cytotoxicity. It was connected and tested by a Resistor Plate before the RTCA Single Plate station was placed inside the incubator at 37 °C and 5% CO2. First, the optimal seeding concentration for the experiments was optimized for each cell line. After seeding, the respective number of cells in 190 µL of medium per well of the E-plate 96, the proliferation, attachment and spreading of the cells were monitored every 30 min by the xCELLigence system. Approximately 24 h after seeding, when the cells were in the log growth phase, the cells were exposed in triplicate to 10 µL of sterile deionized water containing bersavine, to obtain final concentrations of 1–50 μM. Controls received sterile deionized water + DMSO with a final concentration of 0.1%. Cells treated with 5% DMSO were used as a positive control. Growth curves were normalized to the time point of treatment. Evaluations were performed using xCELLigence 1.2.1 software (Roche, Basel, Switzerland and ACEA Biosciences, San Diego, CA, USA).
4.5. Trypan Blue Exclusion Test for Cell Proliferation and Viability
Cell proliferation and the viability of Jurkat and MOLT-4 cells were determined 24 and 48 h after treatment with 5, 10 and 20 μM of bersavine. Cells treated with 5 µM of cisplatin were used as a positive control. Cell membrane integrity was determined using the Trypan blue exclusion technique—mixing 10 μL of 0.4% Trypan blue (Sigma-Aldrich, St. Louis, MO, USA) and 10 μL of cell suspension. Cell counts were carried out using a Bürker chamber and a Nikon Eclipse E200 light microscope (Nikon, Tokyo, Japan).
4.6. Cell Cycle Distribution and Internucleosomal DNA Fragmentation Analysis
Where cell cycle distribution analysis is concerned, the cells were washed with ice-cold Phosphate Buffered Saline PBS and fixed with 70% ethanol. In order to detect low-molecular-weight fragments of DNA, the cells were incubated for 5 min at room temperature in a buffer (192 mL 0.2 M Na2HPO4 + 8 mL of 0.1 M citric acid, pH 7.8) and then labelled with propidium iodide in Vindelov’s solution for 1 h at 37 °C. The DNA content was determined using a CyAn flow cytometer (Beckman Coulter, Miami, FL, USA) with an excitation wavelength of 488 nm. The data were analysed using Multicycle AV software (Phoenix Flow Systems, San Diego, CA, USA).
4.7. Activity of Caspases
The induction of programmed cell death was determined by monitoring the activities of caspases-3/7, caspase-8 and caspase-9 by Caspase-Glo Assays (Promega, Madison, WI, USA) 24 and 48 h after treatment with 5 and 10 μM of bersavine. Cells treated with 5 µM of cisplatin were used as a positive control. The assay provides a proluminogenic substrate in an optimized buffer system. The addition of a Caspase-Glo Reagent results in cell lysis, followed by caspase cleavage of the substrate and the generation of a luminescent signal. A total of 1 × 104 cells were seeded per well using a 96-well plate format (Sigma-Aldrich, St. Louis, MO, USA). After treatment, the Caspase-Glo Assay Reagent was added to each well (50 μL/well) and incubated for 30 min before luminescence was measured using a Tecan Infinite M200 microplate reader (Tecan Group, Männedorf, Switzerland).
4.8. Analysis of Apoptosis
Apoptosis was determined by flow cytometry using an Alexa Fluor® 488 Annexin V/Dead Cell Apoptosis kit (Life Technologies, Grand Island, NY, USA) in accordance with the manufacturer’s instructions. The Alexa Fluor® 488 Annexin V/Dead Cell Apoptosis kit employs the property of Alexa Fluor® 488 conjugated to Annexin V to bind to phosphatidylserine in the presence of Ca2+, and the property of propidium iodide (PI) to enter cells with damaged cell membranes and to bind to DNA. Measurement was performed immediately using a CyAn flow cytometer (Beckman Coulter, Miami, FL, USA). Listmode data were analysed using Kaluza Analysis 1.3 software (Beckman Coulter, Miami, FL, USA).
4.9. Western Blot Analysis
Whole-cell lysates (Cell Lysis Buffer, Cell Signaling Technology, Danvers, MA, USA) were prepared 24 h following treatment of Jurkat and MOLT-4 cells with 5 and 10 μM of bersavine. Cells treated with 0.1% DMSO were used as a negative control. Cells treated with 5 μM of cisplatin were used as a positive control. Quantification of the protein content was performed using the bicinchoninic acid (BCA) assay (Sigma-Aldrich, St. Louis, MO, USA). The lysates (20 µg of purified protein) were loaded into lanes of polyacrylamide gel. After electrophoresis separation, the proteins were transferred to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad, Hercules, CA, USA). Any nonspecific binding of the membranes were blocked for 1 h in a Tris-buffered saline (TBS) containing 0.05% Tween 20 and 10% w/v nonfat dry milk. The membranes were washed in TBS. Incubation with a primary antibody against specific antigens (Chk1, Chk1_serine 345-Cell Signalling, Danvers, MA, USA; Rb, Rb_serine 807 and serine 811–Cell Signalling, Danvers, MA, USA; SAPK/JNK, SAPK/JNK_threonine 183 and tyrosine 185–Cell Signalling, Danvers, MA, USA; β-actin–Sigma-Aldrich, St. Louis, MO, USA; ERK1/2_threonine 202 and tyrosine 204–Cell Signalling, Danvers, MA, USA; p53, p53_serine 392–Exbio, Prague, Czech Republic) was performed at 4° C overnight. The following day, the membranes were washed 5 times with TBS, each time for 5 min, and once with TBS for 10 min, and then incubated with an appropriate secondary antibody (DakoCytomation, Glostrup, Denmark) for 1 h at room temperature. Band detection was performed using a chemiluminiscence detection kit (Roche, Basel, Switzerland). To ensure equal protein loading, each membrane was reprobed and β-actin was detected.
4.10. Calculation of IC50 Values
IC50 values were calculated on the basis of data obtained from proliferation/viability determined by the use of a WST-1 assay and were processed using GraphPad Prism 7 biostatistics (GraphPad Software, San Diego, CA, USA) software. Drug concentrations were plotted against the percentage of cell proliferation/viability and the IC50 values were determined using nonlinear regression.
4.11. Statistical Analysis
The descriptive statistics of the results were calculated and the charts were made using either Microsoft Office Excel 2010 (Microsoft, Redmond, WA, USA) or GraphPad Prism 7 biostatistics (GraphPad Software, San Diego, CA, USA) software. In this study, all the values were expressed as arithmetic means with the SD of triplicates, unless otherwise noted. For quantitative data, normality testing was performed to assess whether parametric or nonparametric tests should be used. For experiments with parametric variables, the significant differences between the groups were analysed using the Student’s t-test and a p-value < 0.05 was considered significant.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}