Increasing Susceptibility of Drug-Resistant Candida albicans to Fluconazole and Terbinafine by 2(5H)-Furanone Derivative

 ,
,  , and
, and
Abstract
1. Introduction
2. Results
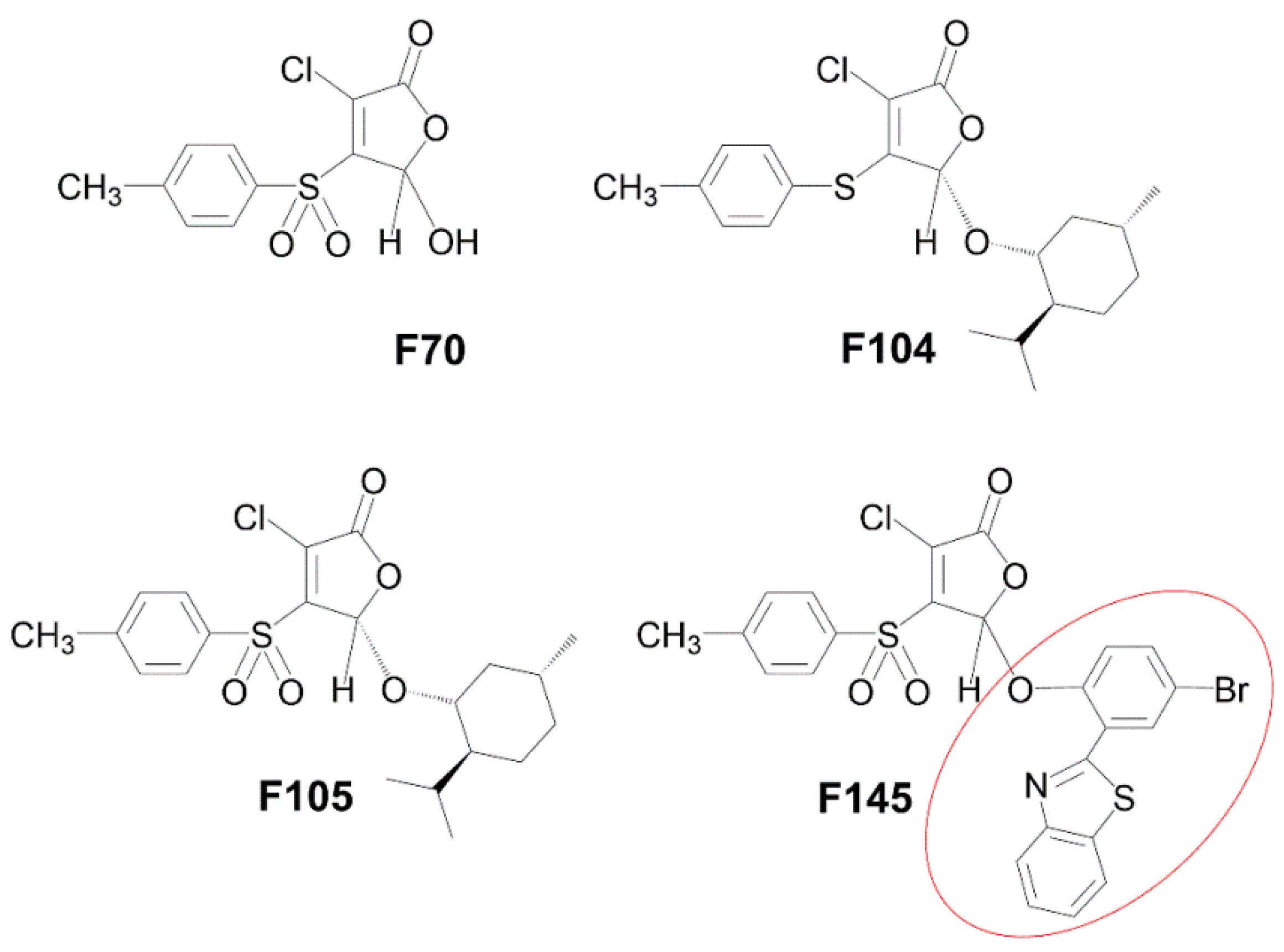
2.1. Antimycotic Activity of 2(5H)-Furanone Derivatives
2.1.1. Susceptibility of C. albicans Isolates to F105 and Conventional Antimycotics
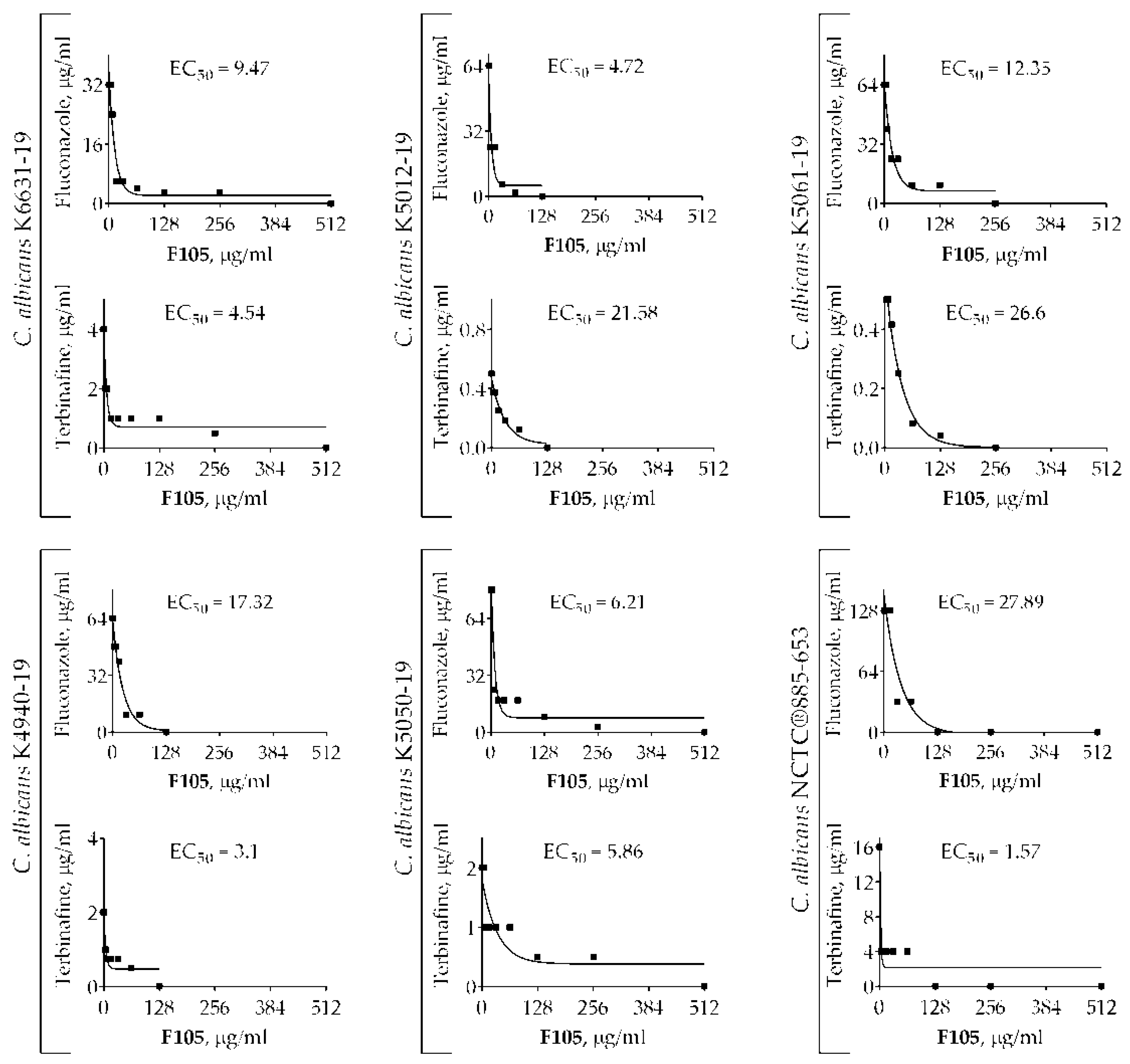
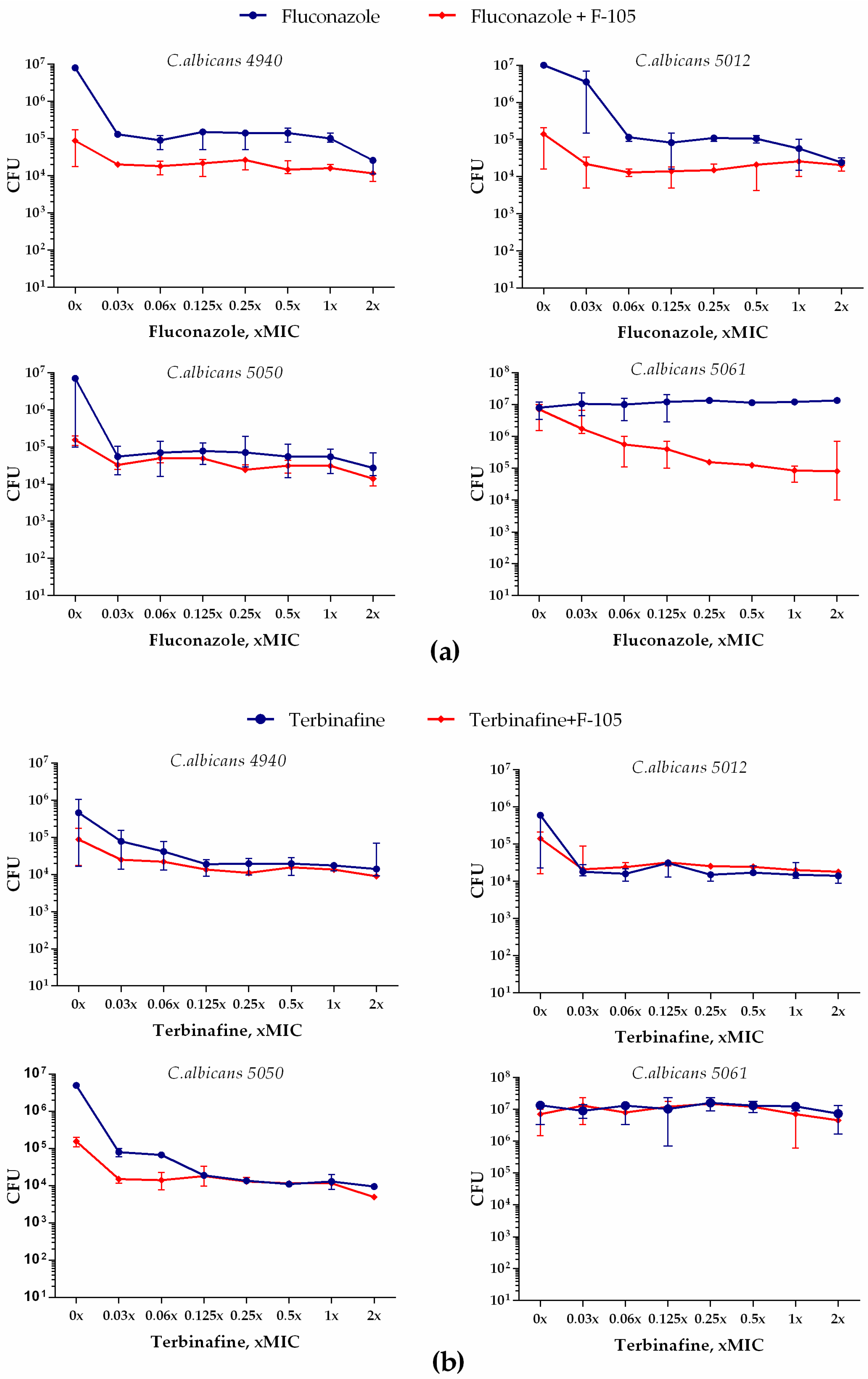
2.1.2. F105 Potentiated the Antifungal Activity of Fluconazole and Terbinafine against C. albicans Cells
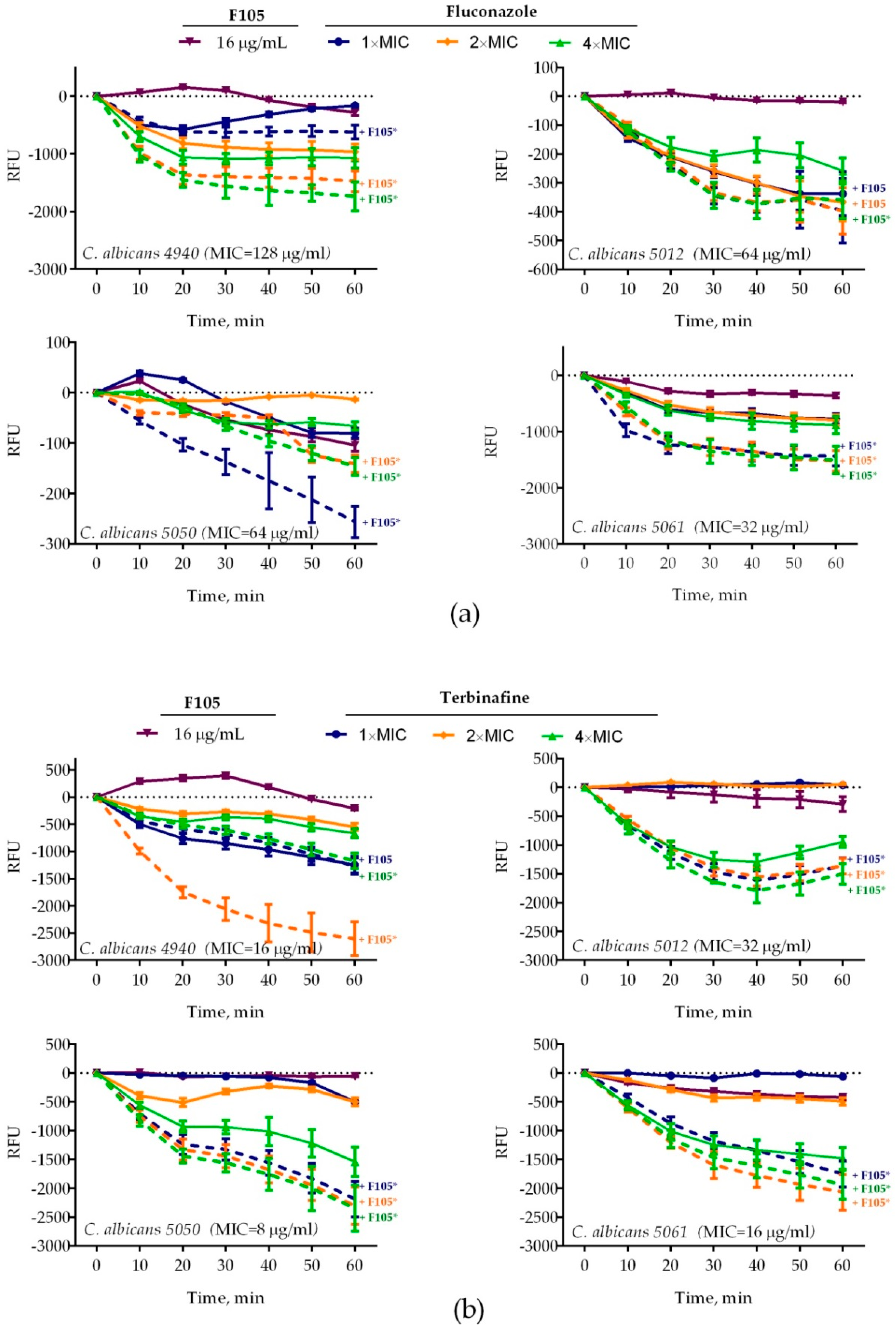
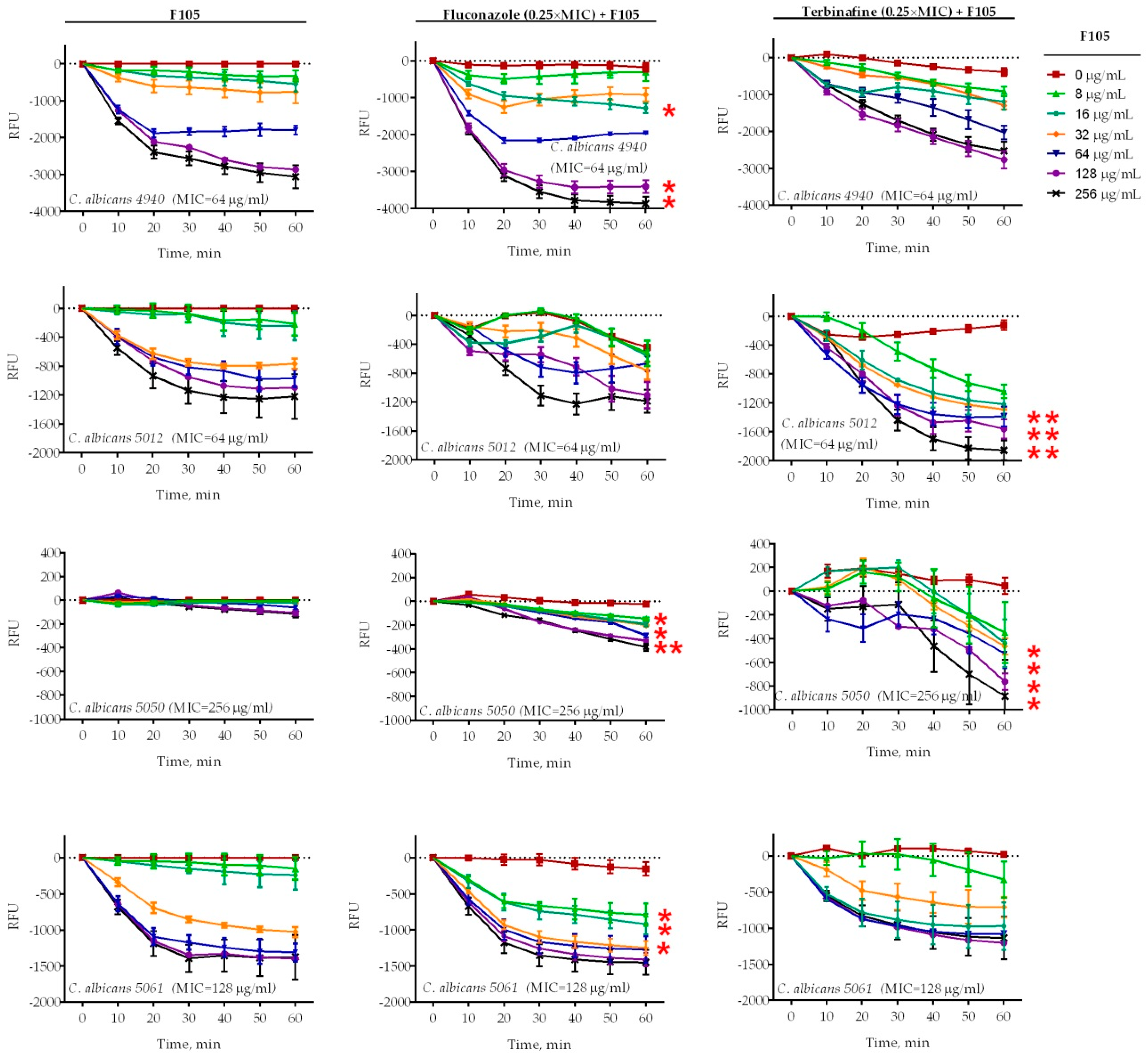
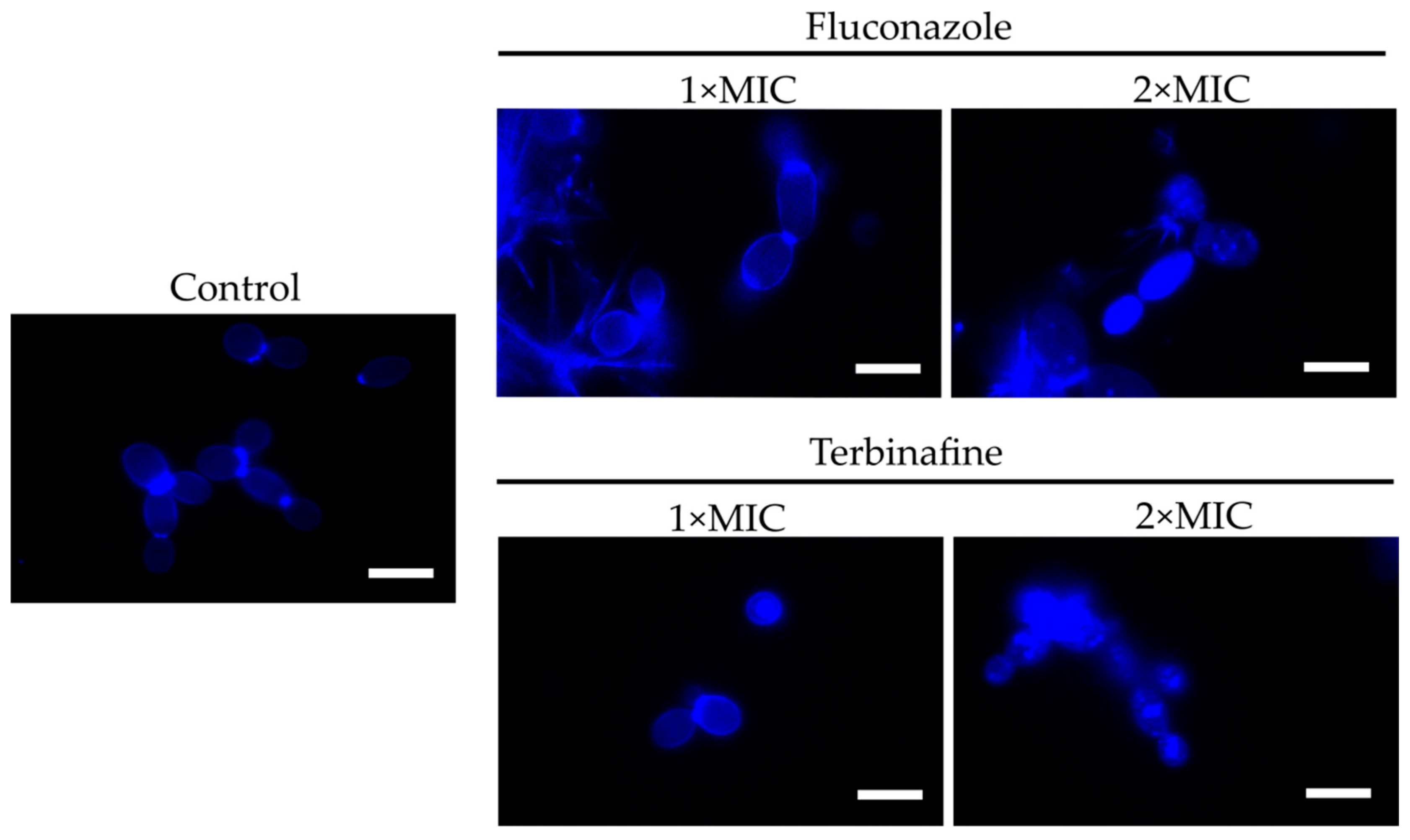
2.2. F105 Enhanced the Capacity of Fluconazole and Terbinafine to Disturb C. albicans Plasma Membrane Integrity
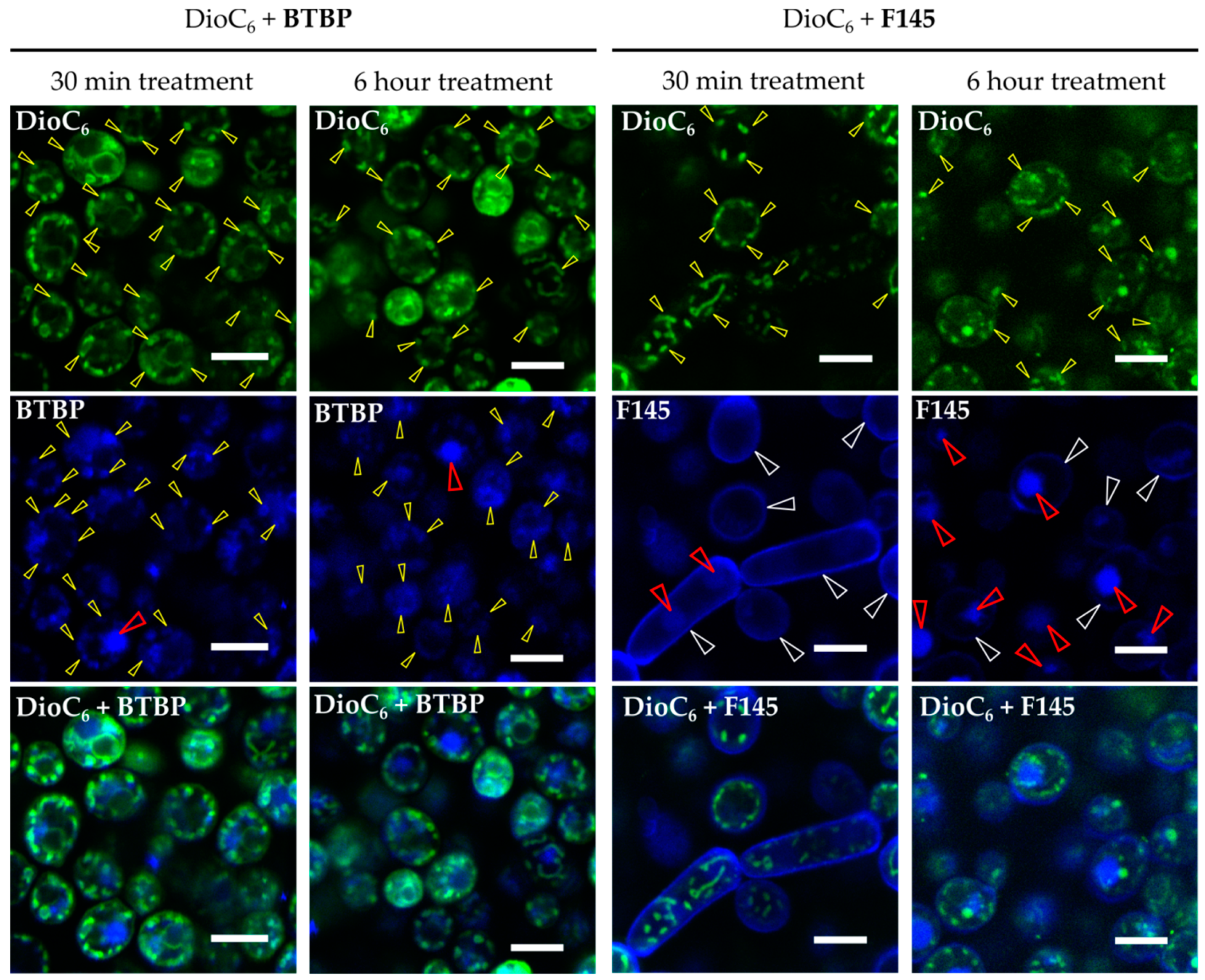
2.3. Penetration of Fluorescent 2(5H)-Furanone Derivative into C. albicans Cells
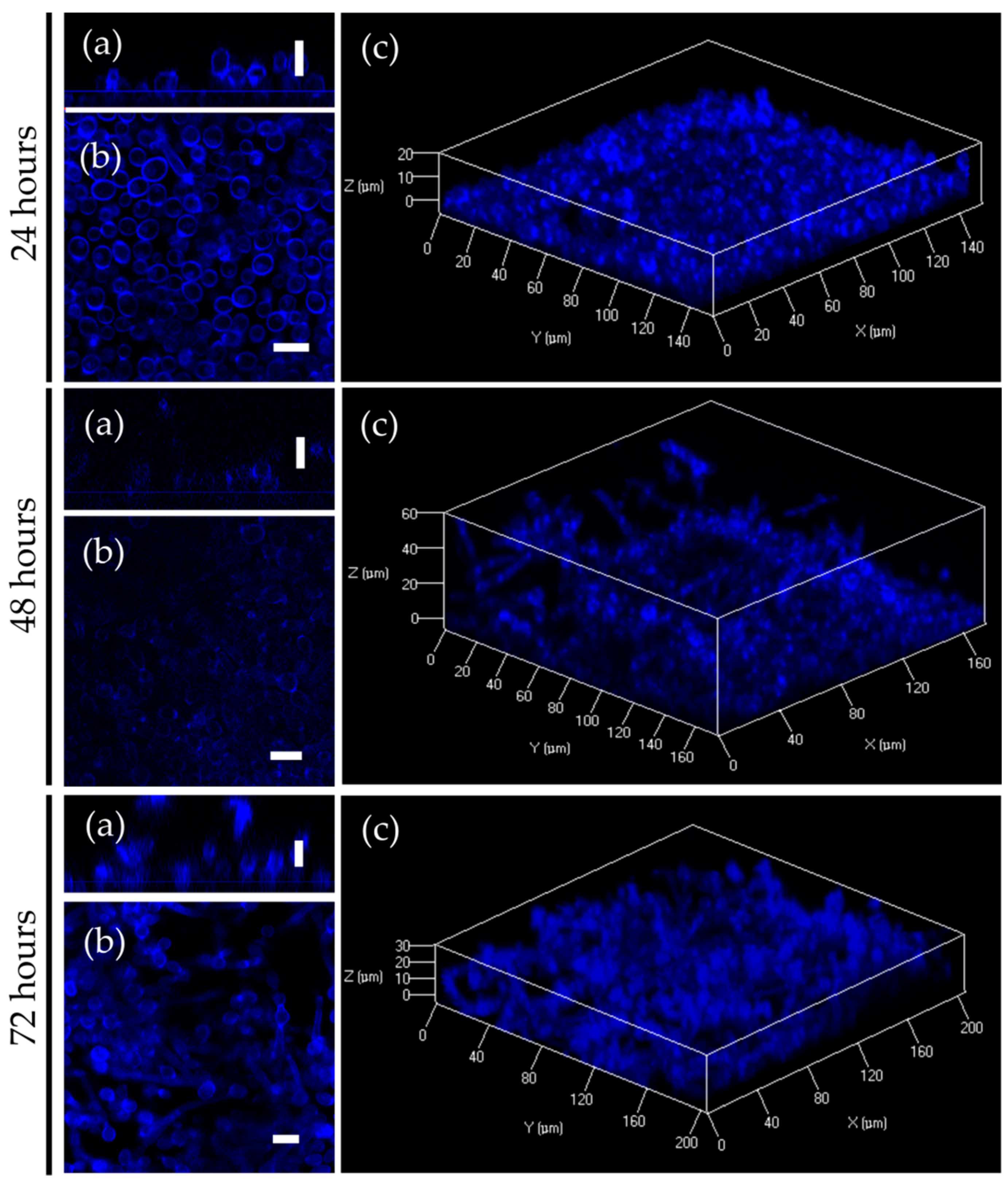
2.4. Penetration of Fluorescent 2(5H)-Furanone Derivative into C. albicans Biofilm
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and Growth Conditions
5.2. Tested Compounds
5.3. Determination of Minimum Inhibitory Concentration (MIC)
5.4. Assessment of Synergy Between F105 and Conventional Antifungal Agents
5.5. Membrane Potential Evaluation
5.6. Confocal Laser Scanning Microscopy
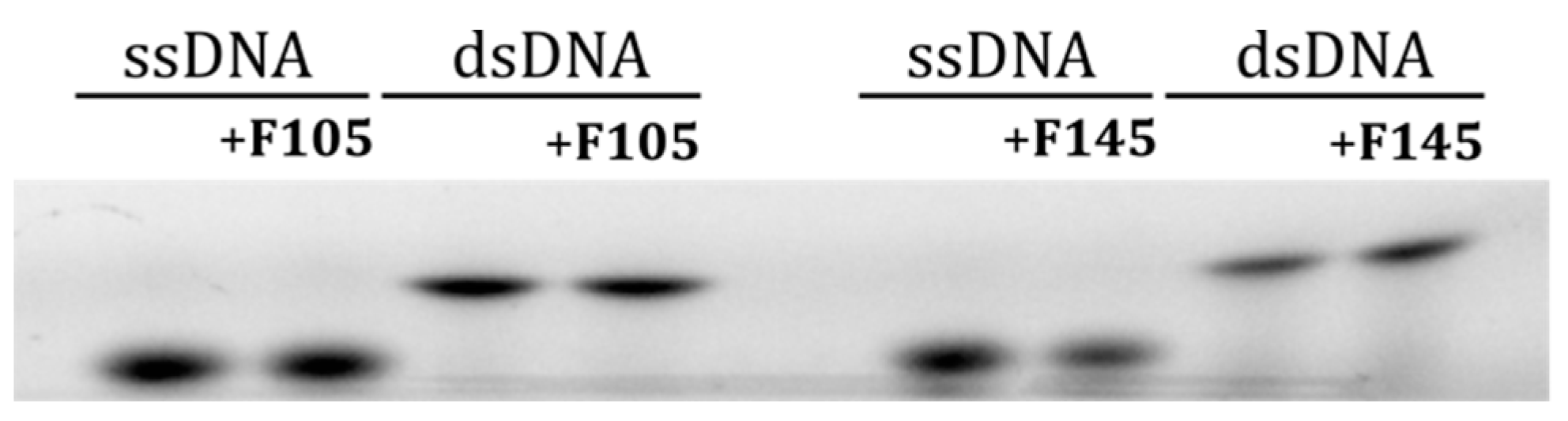
5.7. Investigation of DNA-Binding Ability of F105
Author Contributions
Funding
Conflicts of Interest
References
- Hofs, S.; Mogavero, S.; Hube, B. Interaction of Candida albicans with host cells: Virulence factors, host defense, escape strategies, and the microbiota. J. Microbiol. 2016, 54, 149–169. [Google Scholar] [CrossRef] [PubMed]
- Metwalli, K.H.; Khan, S.A.; Krom, B.P.; Jabra-Rizk, M.A. Streptococcus mutans, Candida albicans, and the Human Mouth: A Sticky Situation. Plos Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Kashem, S.W.; Kaplan, D.H. Skin Immunity to Candida albicans. Trends Immunol. 2016, 37, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.H.; Xie, J.; Xu, J.P.; Li, J.; Li, D.M.; Zong, L.L.; Zheng, Y.C.; Bai, F.Y. Prevalence of specific and phylogenetically closely related genotypes in the population of Candida albicans associated with genital candidiasis in China. Fungal Genet. Biol. 2012, 49, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Scheibler, E.; Garcia, M.C.R.; da Silva, R.M.; Figueiredo, M.A.; Salum, F.G.; Cherubini, K. Use of nystatin and chlorhexidine in oral medicine: Properties, indications and pitfalls with focus on geriatric patients. Gerodontology 2017, 34, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Eschenauer, G.A.; Nguyen, M.H.; Clancy, C.J. Is Fluconazole or an Echinocandin the Agent of Choice for Candidemia. Ann. Pharmacother. 2015, 49, 1068–1074. [Google Scholar] [CrossRef]
- Baixench, M.T.; Aoun, N.; Desnos-Ollivier, M.; Garcia-Hermoso, D.; Bretagne, S.; Ramires, S.; Piketty, C.; Dannaoui, E. Acquired resistance to echinocandins in Candida albicans: Case report and review. J. Antimicrob. Chemother. 2007, 59, 1076–1083. [Google Scholar] [CrossRef]
- Wiederhold, N.P. Antifungal resistance: Current trends and future strategies to combat. Infect. Drug Resist. 2017, 10, 249–259. [Google Scholar] [CrossRef]
- Ryder, N.S.; Wagner, S.; Leitner, I. In vitro activities of terbinafine against cutaneous isolates of Candida albicans and other pathogenic yeasts. Antimicrob. Agents Chemother. 1998, 42, 1057–1061. [Google Scholar] [CrossRef]
- Hu, Y.X.; Hu, Y.Q.; Lu, Y.; Huang, S.Y.; Liu, K.X.; Han, X.; Mao, Z.H.; Wu, Z.; Zhou, X.Y. A Case Report of Penile Infection Caused by Fluconazole- and Terbinafine-Resistant Candida albicans. Mycopathologia 2017, 182, 397–402. [Google Scholar] [CrossRef]
- Berkow, E.L.; Lockhart, S.R. Fluconazole resistance in Candida species: A current perspective. Infect. Drug Resist. 2017, 10, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Corte, L.; Roscini, L.; Colabella, C.; Tascini, C.; Leonildi, A.; Sozio, E.; Menichetti, F.; Merelli, M.; Scarparo, C.; Meyer, W.; et al. Exploring ecological modelling to investigate factors governing the colonization success in nosocomial environment of Candida albicans and other pathogenic yeasts. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Song, Z.; Hentzer, M.; Andersen, J.B.; Molin, S.; Givskov, M.; Hoiby, N. Synthetic furanones inhibit quorum-sensing and enhance bacterial clearance in Pseudomonas aeruginosa lung infection in mice. J. Antimicrob. Chemother. 2004, 53, 1054–1061. [Google Scholar] [CrossRef]
- Janssens, J.C.A.; Steenackers, H.; Robijns, S.; Gellens, E.; Levin, J.; Zhao, H.; Hermans, K.; De Coster, D.; Verhoeven, T.L.; Marchal, K.; et al. Brominated Furanones Inhibit Biofilm Formation by Salmonella enterica Serovar Typhimurium. Appl. Environ. Microbiol. 2008, 74, 6639–6648. [Google Scholar] [CrossRef]
- Hentzer, M.; Riedel, K.; Rasmussen, T.B.; Heydorn, A.; Andersen, J.B.; Parsek, M.R.; Rice, S.A.; Eberl, L.; Molin, S.; Hoiby, N.; et al. Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiol. -Sgm 2002, 148, 87–102. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Trizna, E.Y.; Baidamshina, D.R.; Ryzhikova, M.N.; Sibgatullina, R.R.; Khabibrakhmanova, A.M.; Latypova, L.Z.; Kurbangalieva, A.R.; Rozhina, E.V.; Klinger-Strobel, M.; et al. Antimicrobial Effects of Sulfonyl Derivative of 2(5H)-Furanone against Planktonic and Biofilm Associated Methicillin-Resistant and -Susceptible Staphylococcus aureus. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Lonn-Stensrud, J.; Landin, M.A.; Benneche, T.; Petersen, F.C.; Scheie, A.A. Furanones, potential agents for preventing Staphylococcus epidermidis biofilm infections? J. Antimicrob. Chemother. 2009, 63, 309–316. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Pavlova, A.S.; Khabibrakhmanova, A.M.; Faizova, G.M.; Kurbangalieva, A.R.; Katsunori, T.; Trizna, E.Y.; Baidamshina, D.R.; Bogachev, M.I.; Kayumov, A.R. Targeting Bacillus cereus cells: Increasing efficiency of antimicrobials by the bornyl-possessing 2(5H)-furanone derivative. New Microbiol. 2019. [Google Scholar]
- Buchta, V.; Pour, M.; Kubanova, P.; Silva, L.; Votruba, I.; Voprsalova, M.; Schiller, R.; Fakova, H.; Spulak, M. In vitro activities of 3-(halogenated phenyl)-5-acyloxymethyl-2,5-dihydrofuran-2-ones against common and emerging yeasts and molds. Antimicrob. Agents Chemother. 2004, 48, 873–878. [Google Scholar] [CrossRef]
- Husain, A.; Alam, M.M.; Shaharyar, M.; Lal, S. Antimicrobial activities of some synthetic butenolides and their pyrrolone derivatives. J. Enzym. Inhib. Med. Chem. 2010, 25, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Abdel-Razek, O.A.; Cheng, F.; Bandyopadhyay, D.; Shetye, G.S.; Wang, G.R.; Luk, Y.Y. Bicyclic brominated furanones: A new class of quorum sensing modulators that inhibit bacterial biofilm formation. Bioorganic Med. Chem. 2014, 22, 1313–1317. [Google Scholar] [CrossRef] [PubMed]
- Kuehl, R.; Al-Bataineh, S.; Gordon, O.; Luginbuehl, R.; Otto, M.; Textor, M.; Landmann, R. Furanone at Subinhibitory Concentrations Enhances Staphylococcal Biofilm Formation by luxS Repression. Antimicrob. Agents Chemother. 2009, 53, 4159–4166. [Google Scholar] [CrossRef] [PubMed]
- Trizna, E.; Latypova, L.; Kurbangalieva, A.; Bogachev, M.I.; Kayumov, A. 2(5H)-Furanone Derivatives as Inhibitors of Staphylococcal Biofilms. BioNanoScience 2016. [Google Scholar] [CrossRef]
- Latypova, L.Z.; Saigitbatalova, E.S.; Chulakova, D.R.; Lodochnikova, O.A.; Kurbangalieva, A.R.; Berdnikov, E.A.; Chmutova, G.A. Sulfides, sulfones, and sulfoxides of the furan-2(5H)-one series. synthesis and structure. Russ. J. Org. Chem. 2014, 50, 521–534. [Google Scholar] [CrossRef]
- Sharafutdinov, I.S.; Pavlova, A.S.; Akhatova, F.S.; Khabibrakhmanova, A.M.; Rozhina, E.V.; Romanova, Y.J.; Fakhrullin, R.; Lodochnikova, O.A.; Kurbangalieva, A.R.; Bogachev, M.I.; et al. Unraveling the Molecular Mechanism of Selective Antimicrobial Activity of 2(5H)-Furanone Derivative against Staphylococcus aureus. Int. J. Mol. Sci. 2019, 20, 694. [Google Scholar] [CrossRef] [PubMed]
- Sergeyev, Y.V.; Shpigel, B.I.; Sergeev, A.Y. Pharmacotherapy of Mycosis; Medicine for Everyone: Moscow, Russia, 2003; p. 200. [Google Scholar]
- Trizna, E.Y.; Khakimullina, E.N.; Latypova, L.Z.; Kurbangalieva, A.R.; Sharafutdinov, I.S.; Evtyugin, V.G.; Babynin, E.V.; Bogachev, M.I.; Kayumov, A.R. Thio Derivatives of 2(5H)-Furanone As Inhibitors against Bacillus subtilis Biofilms. Acta Nat. 2015, 7, 102–107. [Google Scholar] [CrossRef]
- Fjaervik, E.; Zotchev, S.B. Biosynthesis of the polyene macrolide antibiotic nystatin in Streptomyces noursei. Appl. Microbiol. Biotechnol. 2005, 67, 436–443. [Google Scholar] [CrossRef]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida albicans and Emerging Non-albicans Candida Species. Front. Microbiol. 2017, 7. [Google Scholar] [CrossRef]
- Klis, F.M.; Sosinska, G.J.; de Groot, P.W.J.; Brul, S. Covalently linked cell wall proteins of Candida albicans and their role in fitness and virulence. Fems Yeast Res. 2009, 9, 1013–1028. [Google Scholar] [CrossRef]
- Chaffin, W.L. Candida albicans cell wall proteins. Microbiol. Mol. Biol. Rev. 2008, 72, 495. [Google Scholar] [CrossRef]
- Ene, I.V.; Heilmann, C.J.; Sorgo, A.G.; Walker, L.A.; de Koster, C.G.; Munro, C.A.; Klis, F.M.; Brown, A.J.P. Carbon source-induced reprogramming of the cell wall proteome and secretome modulates the adherence and drug resistance of the fungal pathogen Candida albicans. Proteomics 2012, 12, 3164–3179. [Google Scholar] [CrossRef]
- Angiolella, L.; Micocci, M.M.; D’Alessio, S.; Girolamo, A.; Maras, B.; Cassone, A. Identification of major glucan-associated cell wall proteins of Candida albicans and their role in fluconazole resistance. Antimicrob. Agents Chemother. 2002, 46, 1688–1694. [Google Scholar] [CrossRef]
- Fedorova, K.; Kayumov, A.; Woyda, K.; Ilinskaja, O.; Forchhammer, K. Transcription factor TnrA inhibits the biosynthetic activity of glutamine synthetase in Bacillus subtilis. Febs Lett. 2013, 587, 1293–1298. [Google Scholar] [CrossRef]
- Kean, R.; Rajendran, R.; Haggarty, J.; Townsend, E.M.; Short, B.; Burgess, K.E.; Lang, S.; Millington, O.; Mackay, W.G.; Williams, C.; et al. Candida albicans Mycofilms Support Staphylococcus aureus Colonization and Enhances Miconazole Resistance in Dual-Species Interactions. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kayumov, A.R.; Khakimullina, E.N.; Sharafutdinov, I.S.; Trizna, E.Y.; Latypova, L.Z.; Hoang Thi, L.; Margulis, A.B.; Bogachev, M.I.; Kurbangalieva, A.R. Inhibition of biofilm formation in Bacillus subtilis by new halogenated furanones. J. Antibiot. 2015, 68, 297–301. [Google Scholar] [CrossRef]
- Otsubo, T.; Minami, A.; Fujii, H.; Taguchi, R.; Takahashi, T.; Suzuki, T.; Teraoka, F.; Ikeda, K. 2-(Benzothiazol-2-yl)-phenyl-beta-D-galactopyranoside derivatives as fluorescent pigment dyeing substrates and their application for the assay of beta-D-galactosidase activities. Bioorganic Med. Chem. Lett. 2013, 23, 2245–2249. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef] [PubMed]
- Bogachev, M.I.; Volkov, V.Y.; Markelov, O.A.; Trizna, E.Y.; Baydamshina, D.R.; Melnikov, V.; Murtazina, R.R.; Zelenikhin, P.V.; Sharafutdinov, I.S.; Kayumov, A.R. Fast and simple tool for the quantification of biofilm-embedded cells sub-populations from fluorescent microscopic images. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate/Strain | Source | MIC (μg/mL) | |||
|---|---|---|---|---|---|
| F105 | Fluconazole | Terbinafine | Nystatin | ||
| NCTC®885-653 | NCTC Collection | 64 | 64 | 8 | 8 |
| K3836-19 | Phlegm | 64 | 32 | 4 | 4 |
| K4026-19 | Buccal swab | 32 | 8 | 2 | 8 |
| K4085-19 | Buccal swab | 256 | 16 | 4 | 4 |
| K4146-19 | Buccal swab | 256 | 16 | 4 | 8 |
| K46-19 | Buccal swab | >256 | 128 | 8 | 8 |
| K47-19 | Buccal swab | 256 | 64 | 32 | 16 |
| K4940-19 | Buccal swab | 64 | 128 | 16 | 8 |
| K4941-19 | Buccal swab | >256 | 64 | 8 | 8 |
| K4956-19 | Buccal swab | >256 | >128 | 16 | 4 |
| K4978-19 | Buccal swab | 64 | 32 | 8 | 2 |
| K5002-19 | Cervical canal | >256 | 128 | 16 | 8 |
| K5007-19 | Buccal swab | 64 | >128 | 16 | 4 |
| K5012-19 | Buccal swab | 64 | 64 | 32 | 8 |
| K5014-19 | Buccal swab | 64 | 16 | 16 | 8 |
| K5030-19 | Buccal swab | 32 | 128 | 16 | 8 |
| K5038-19 | Urethral mucosa | >256 | 32 | 8 | 16 |
| K5050-19 | Buccal swab | 256 | 64 | 8 | 2 |
| K5061-19 | Buccal swab | 128 | 32 | 16 | 4 |
| K5077-19 | Buccal swab | 128 | 64 | 16 | 4 |
| K5081-19 | Buccal swab | 128 | 64 | 32 | 4 |
| K5092-19 | Buccal swab | 256 | 128 | 16 | 8 |
| K5094-19 | Buccal swab | >256 | 64 | 16 | 8 |
| K5096-19 | Buccal swab | 32 | 64 | 16 | 2 |
| K5097-19 | Buccal swab | 256 | 64 | 16 | 4 |
| K625-19 | Vaginal swab | 256 | 32 | 4 | 4 |
| K6631-19 | Throat swab | >256 | 32 | 4 | 2 |
| MIC | MIC (F105) | FIC | FIC (F105) | FICImin | EC50 | |
|---|---|---|---|---|---|---|
| Fluconazole | 32 | 512 | 8 | 16 | 0.28 | 9.47 |
| Nystatin | 4 | 512 | 2 | 64 | 0.63 | 106.2 |
| Terbinafine | 4 | 512 | 1 | 16 | 0.28 | 4.54 |
| C. albicans strain | MIC (F105) | MIC (FLC) | FIC (F105) | FIC (FLC) | FICImin | EC50 (F105) | |
| F105 + Fluconazole (FLC) | K6631-19 | 512 | 32 | 16 | 8 | 0.28 | 9.47 |
| K5012-19 | 64 | 64 | 16 | 4 | 0.31 | 4.72 | |
| K5061-19 | 128 | 32 | 4 | 8 | 0.31 | 12.35 | |
| K4940-19 | 64 | 128 | 16 | 16 | 0.38 | 17.32 | |
| K5050-19 | 256 | 64 | 4 | 16 | 0.27 | 6.21 | |
| NCTC®885-653 | 64 | 64 | 16 | 16 | 0.5 | 27.89 | |
| C. albicans strain | MIC (F105) | MIC (TRB) | FIC (F105) | FIC (TRB) | FICImin | EC50 (F105) | |
| F105 + Terbinafine (TRB) | K6631-19 | 512 | 4 | 16 | 1 | 0.28 | 4.54 |
| K5012-19 | 64 | 32 | 16 | 8 | 0.5 | 21.58 | |
| K5061-19 | 128 | 16 | 8 | 2 | 0.56 | 26.6 | |
| K4940-19 | 64 | 16 | 4 | 4 | 0.31 | 3.1 | |
| K5050-19 | 256 | 8 | 64 | 2 | 0.5 | 5.86 | |
| NCTC®885-653 | 64 | 8 | 2 | 2 | 0.28 | 1.57 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharafutdinov, I.S.; Ozhegov, G.D.; Sabirova, A.E.; Novikova, V.V.; Lisovskaya, S.A.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; Kayumov, A.R. Increasing Susceptibility of Drug-Resistant Candida albicans to Fluconazole and Terbinafine by 2(5H)-Furanone Derivative. Molecules 2020, 25, 642. https://doi.org/10.3390/molecules25030642
Sharafutdinov IS, Ozhegov GD, Sabirova AE, Novikova VV, Lisovskaya SA, Khabibrakhmanova AM, Kurbangalieva AR, Bogachev MI, Kayumov AR. Increasing Susceptibility of Drug-Resistant Candida albicans to Fluconazole and Terbinafine by 2(5H)-Furanone Derivative. Molecules. 2020; 25(3):642. https://doi.org/10.3390/molecules25030642
Chicago/Turabian StyleSharafutdinov, Irshad S., Georgii D. Ozhegov, Alina E. Sabirova, Valentina V. Novikova, Svetlana A. Lisovskaya, Alsu M. Khabibrakhmanova, Almira R. Kurbangalieva, Mikhail I. Bogachev, and Airat R. Kayumov. 2020. "Increasing Susceptibility of Drug-Resistant Candida albicans to Fluconazole and Terbinafine by 2(5H)-Furanone Derivative" Molecules 25, no. 3: 642. https://doi.org/10.3390/molecules25030642
APA StyleSharafutdinov, I. S., Ozhegov, G. D., Sabirova, A. E., Novikova, V. V., Lisovskaya, S. A., Khabibrakhmanova, A. M., Kurbangalieva, A. R., Bogachev, M. I., & Kayumov, A. R. (2020). Increasing Susceptibility of Drug-Resistant Candida albicans to Fluconazole and Terbinafine by 2(5H)-Furanone Derivative. Molecules, 25(3), 642. https://doi.org/10.3390/molecules25030642