
Differences in Production, Composition, and Antioxidant Activities of Exopolymeric Substances (EPS) Obtained from Cultures of Endophytic Fusarium culmorum Strains with Different Effects on Cereals

 , , , , , ,
, , , , , ,  and
and
Abstract
1. Introduction
2. Results and Discussion
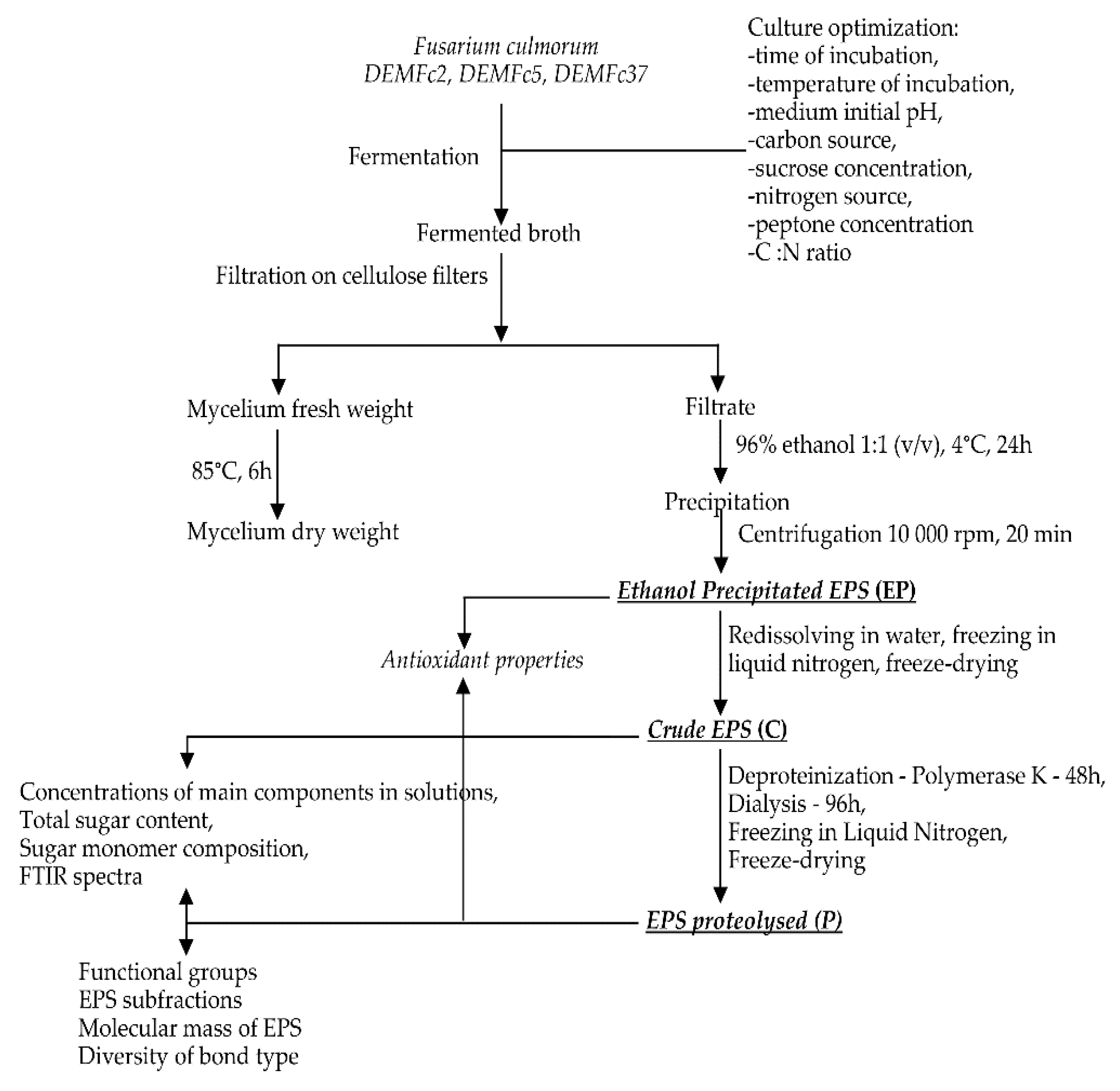
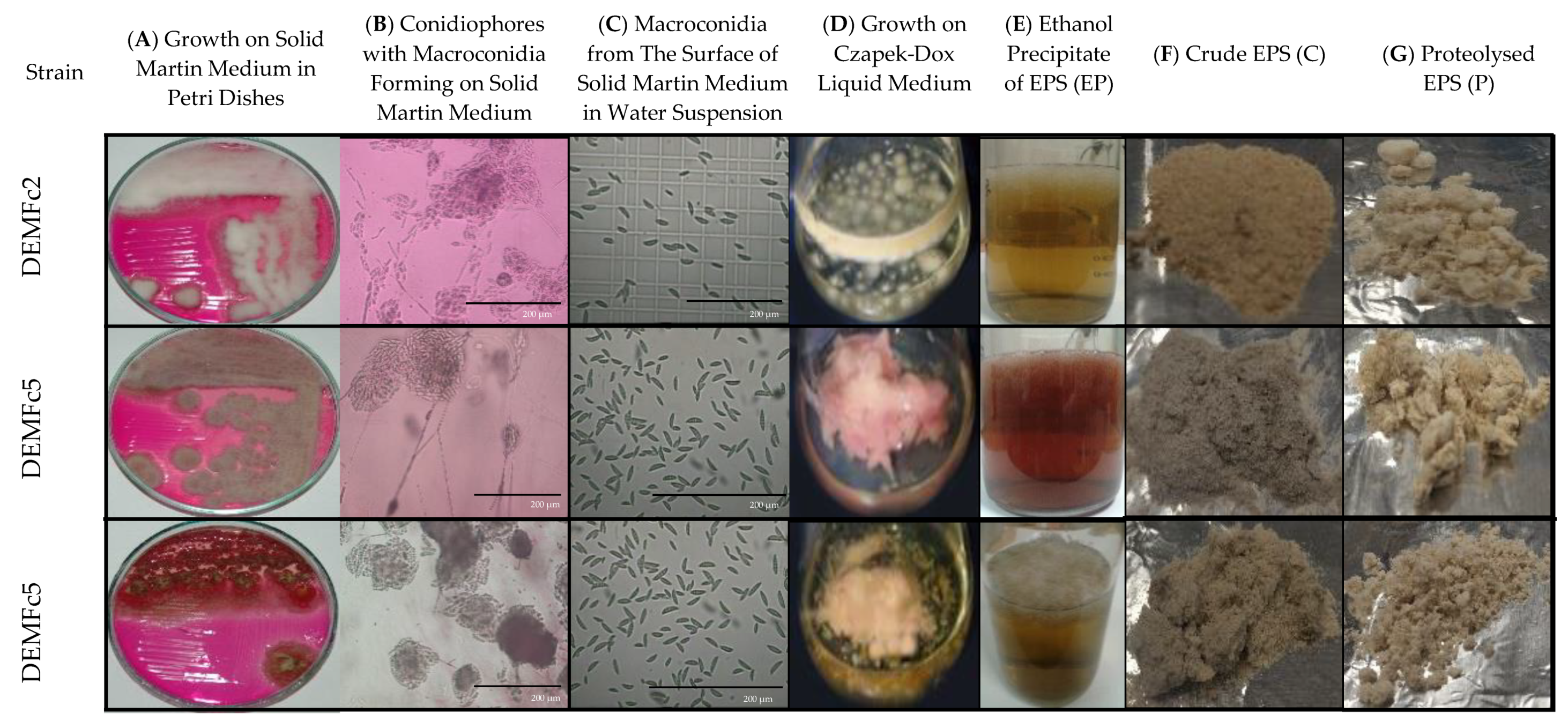
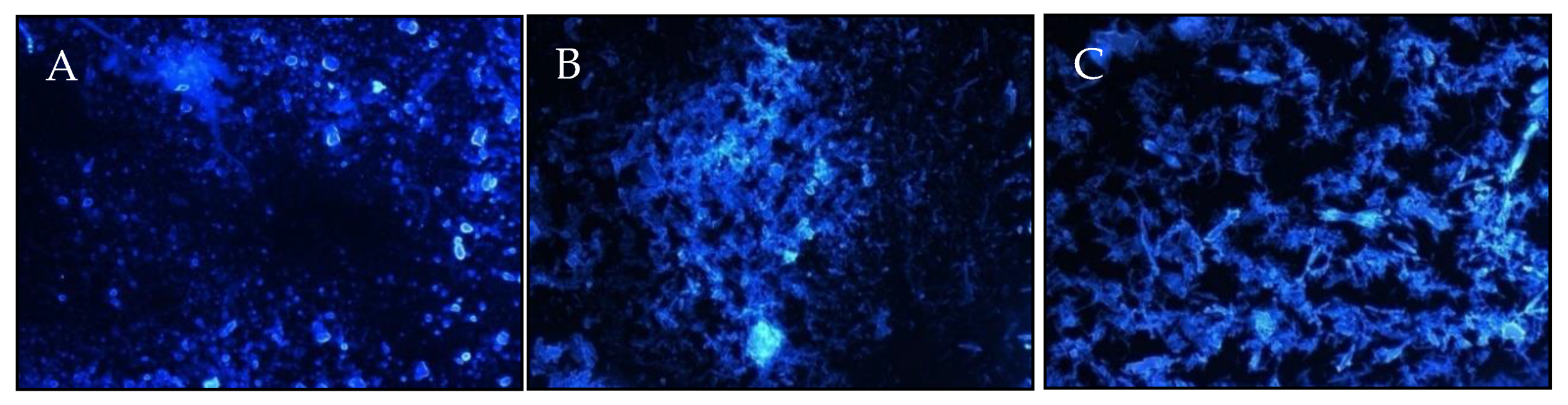
2.1. EPS Isolation and Fractionation
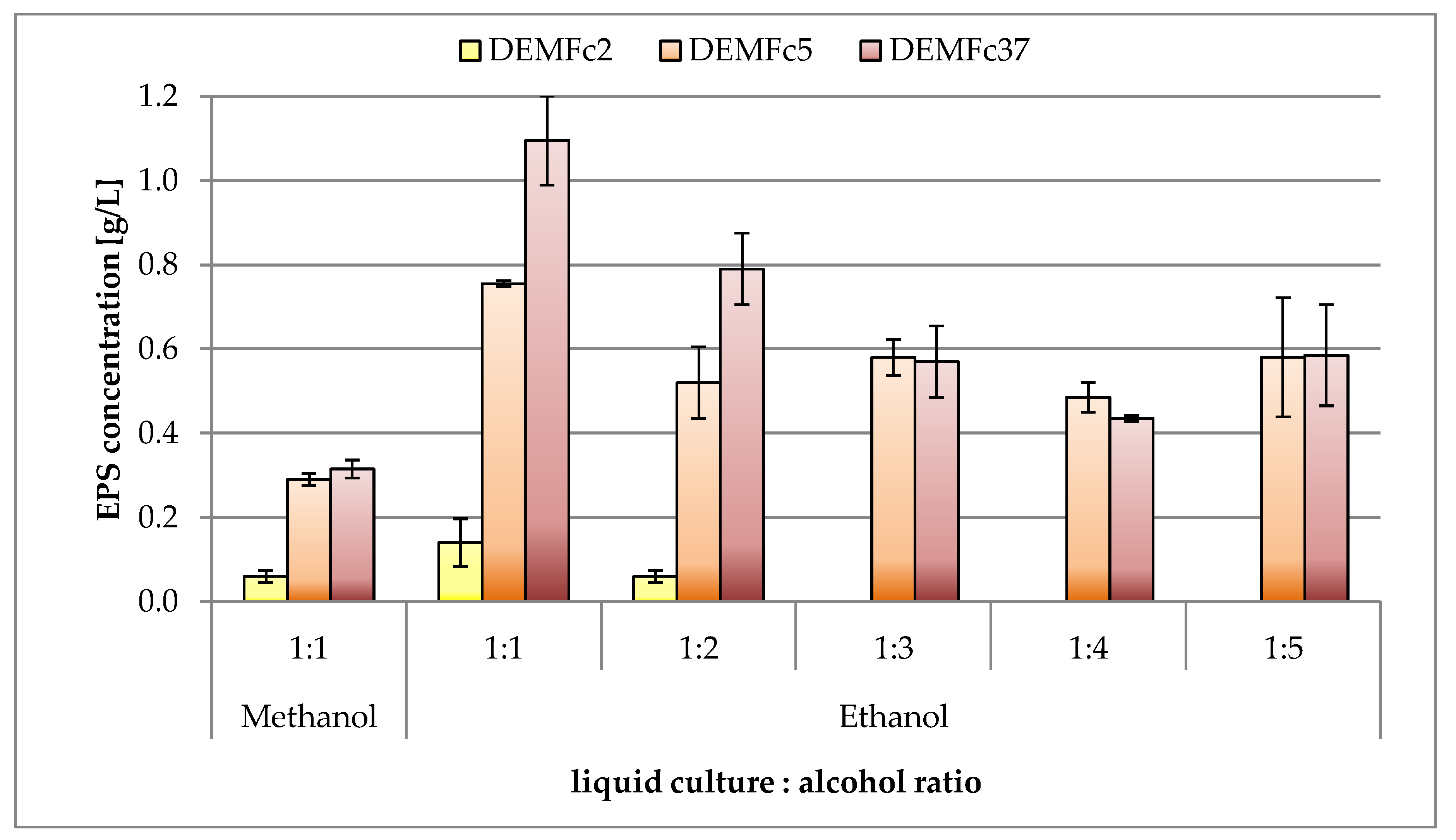
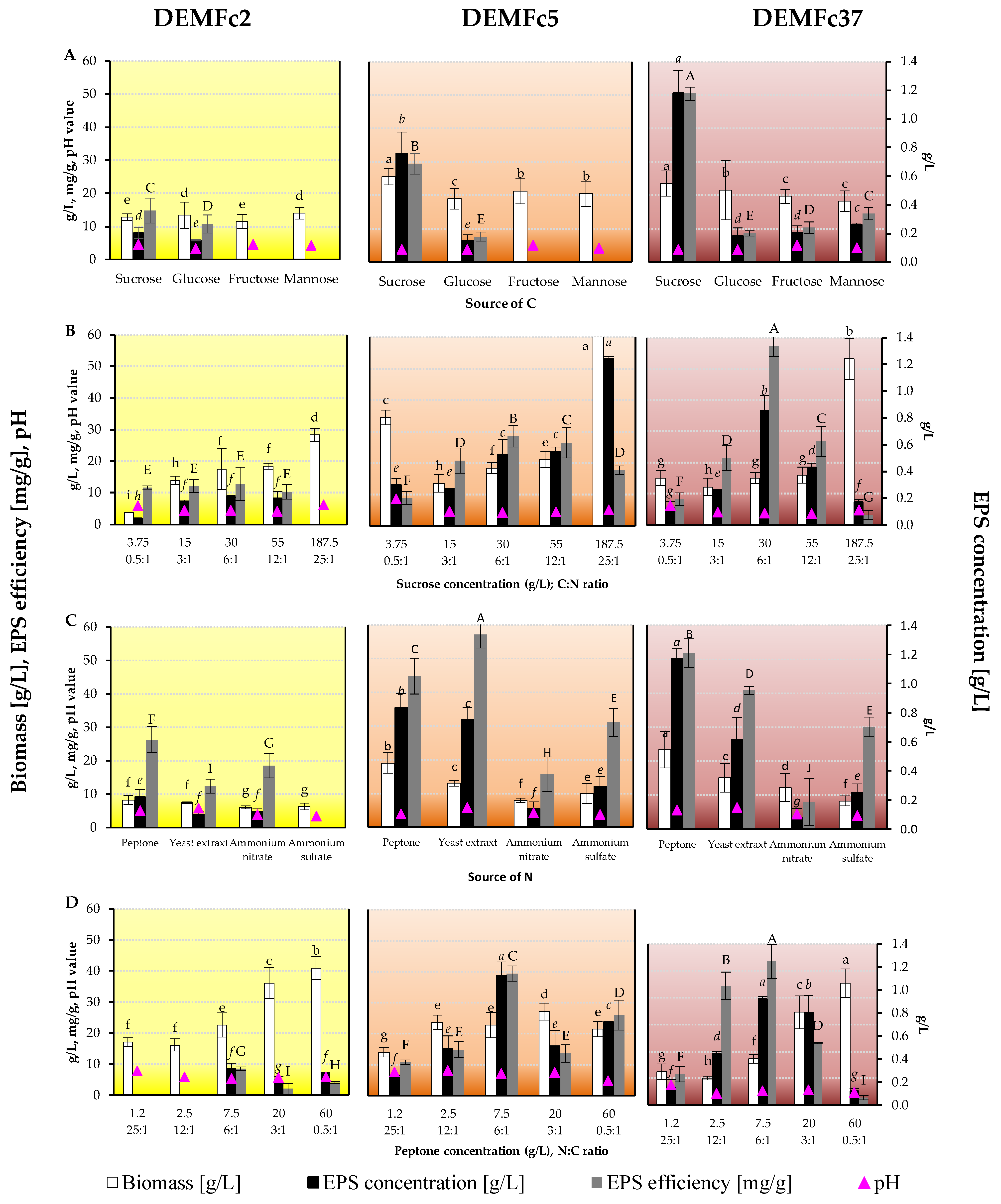
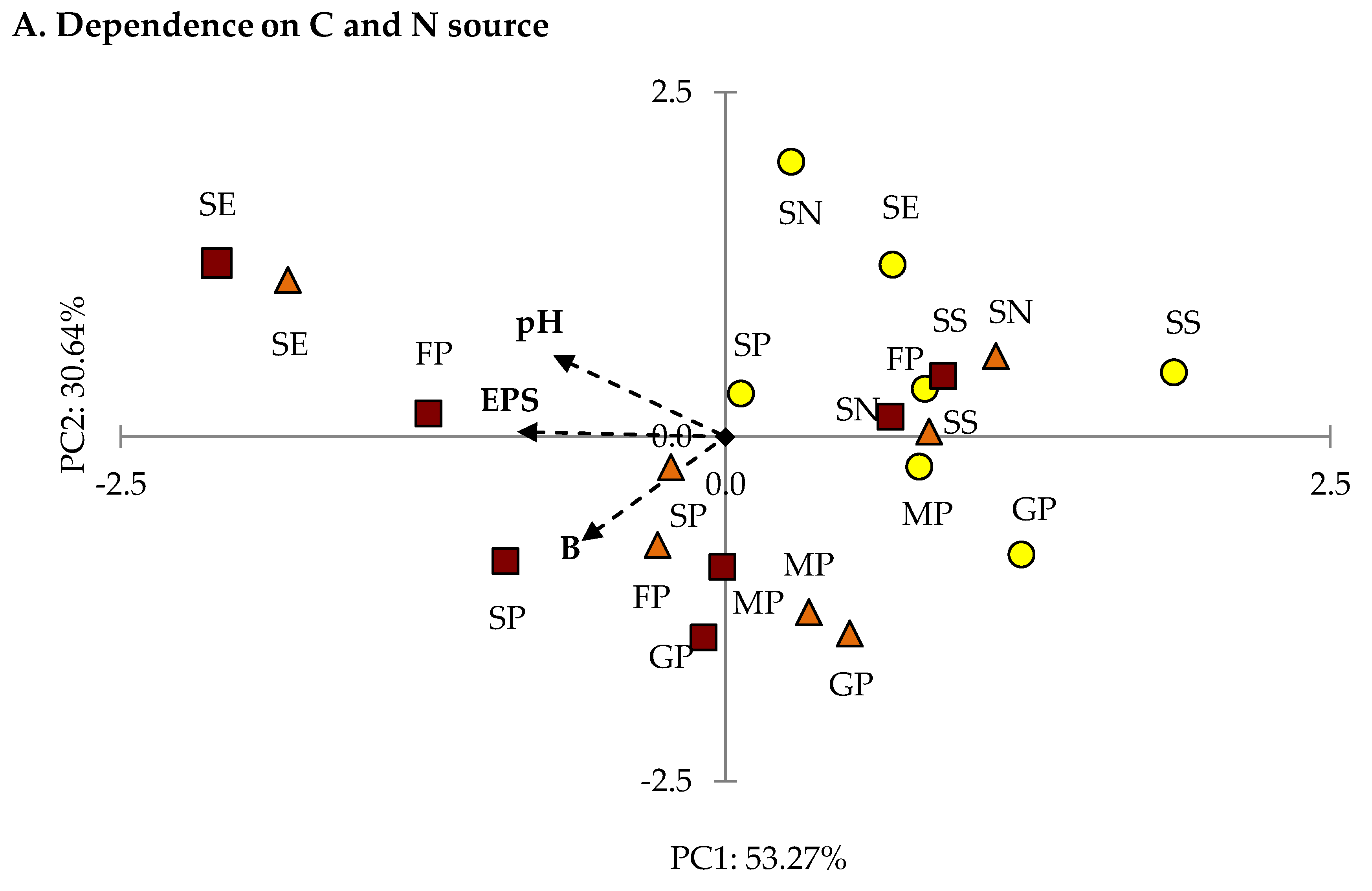
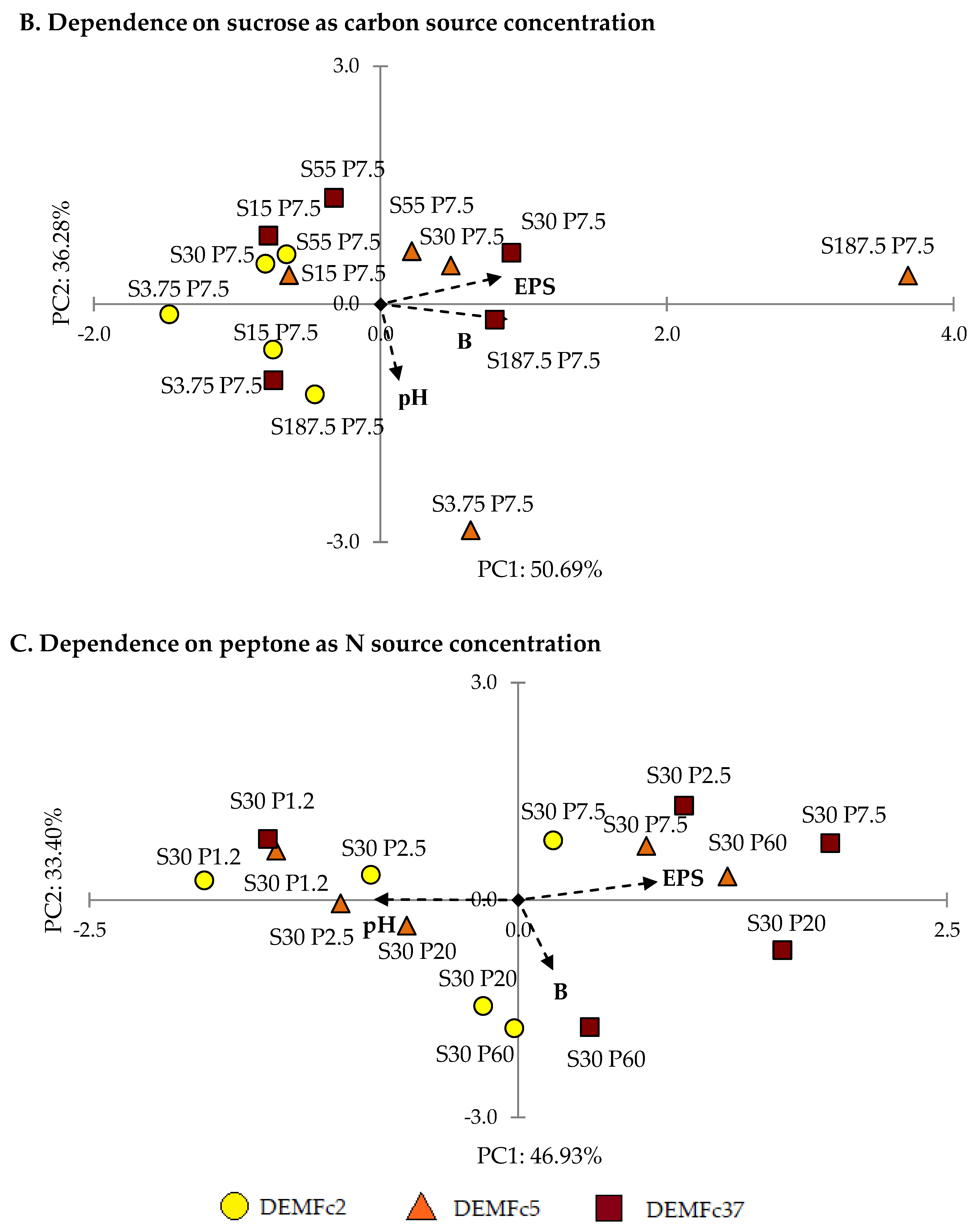
2.2. Optimization of the EPS Production
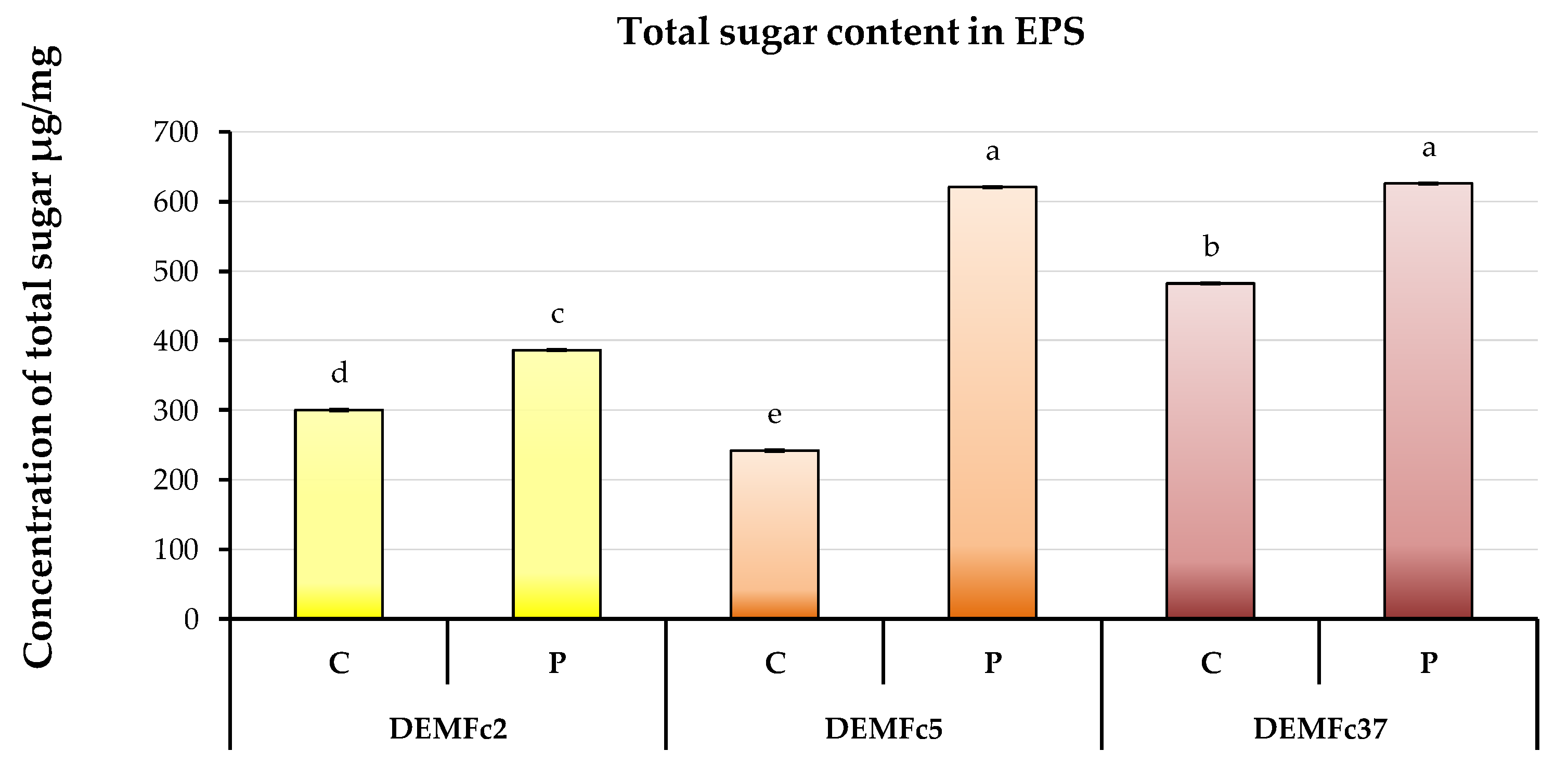
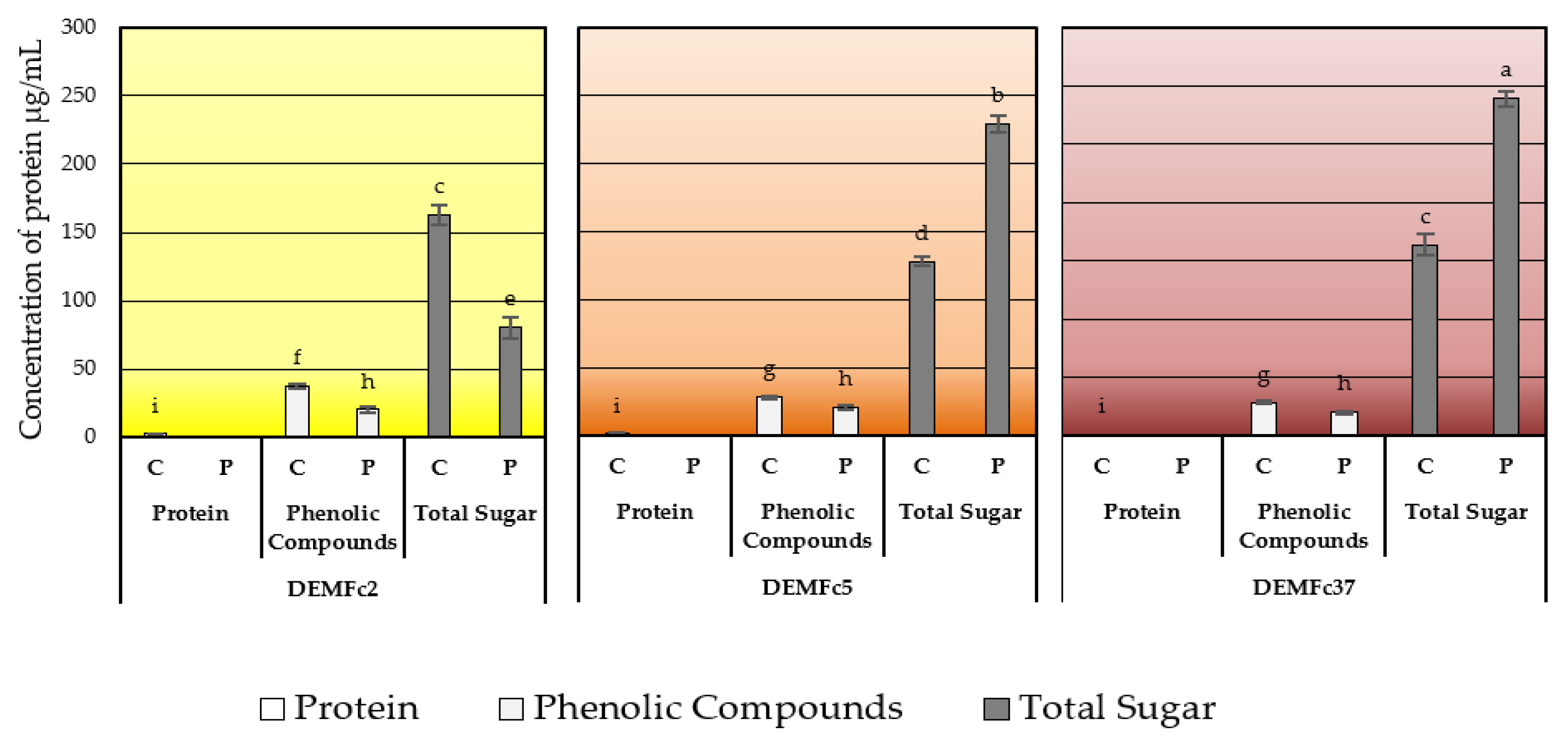
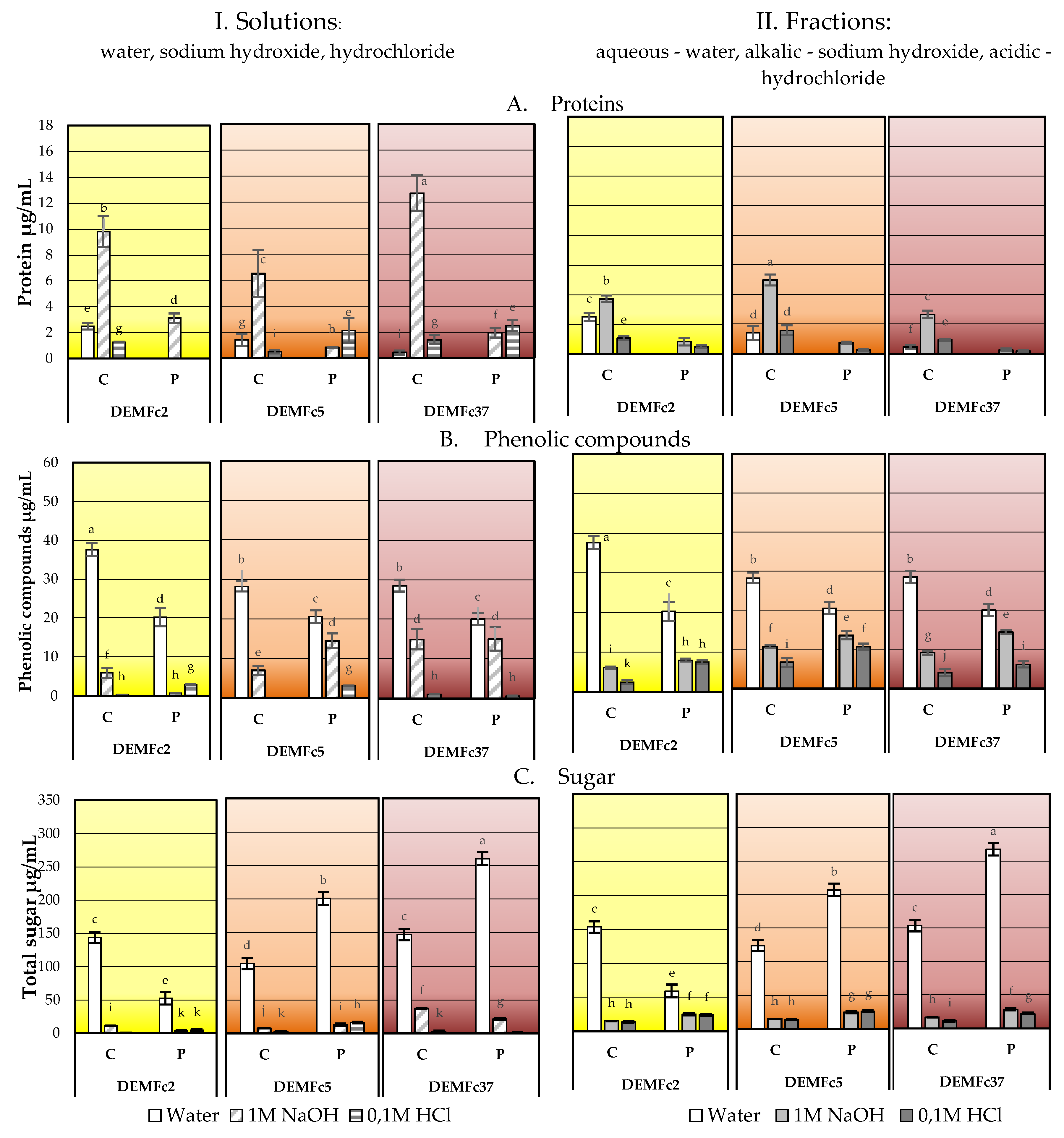
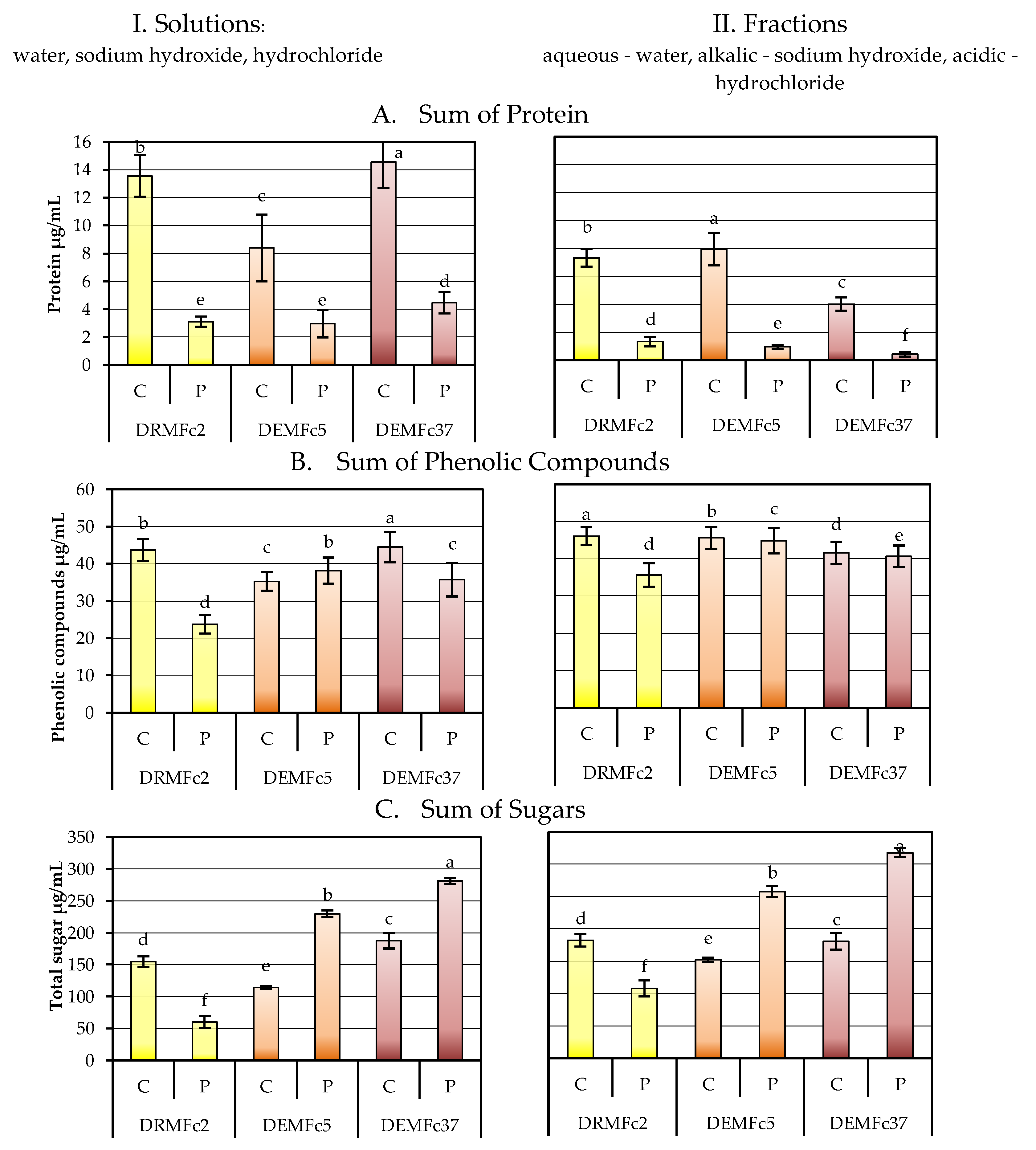
2.3. Composition of the EPS Solution in Three Solvents (Water, Sodium Base, Acid) and Total Sugar Content of EPS
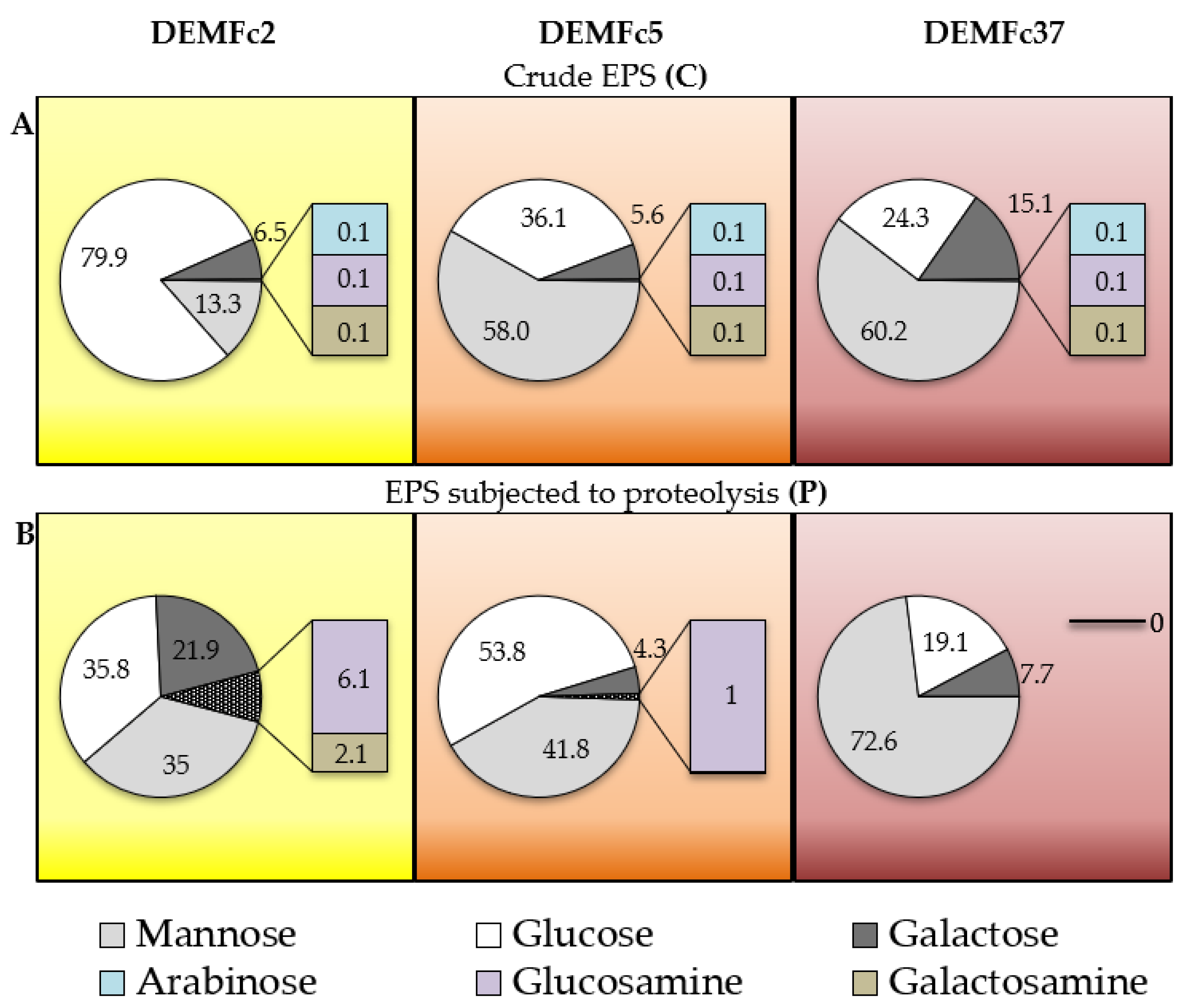
Sugar Composition of the EPS
2.4. Characterization of EPSs
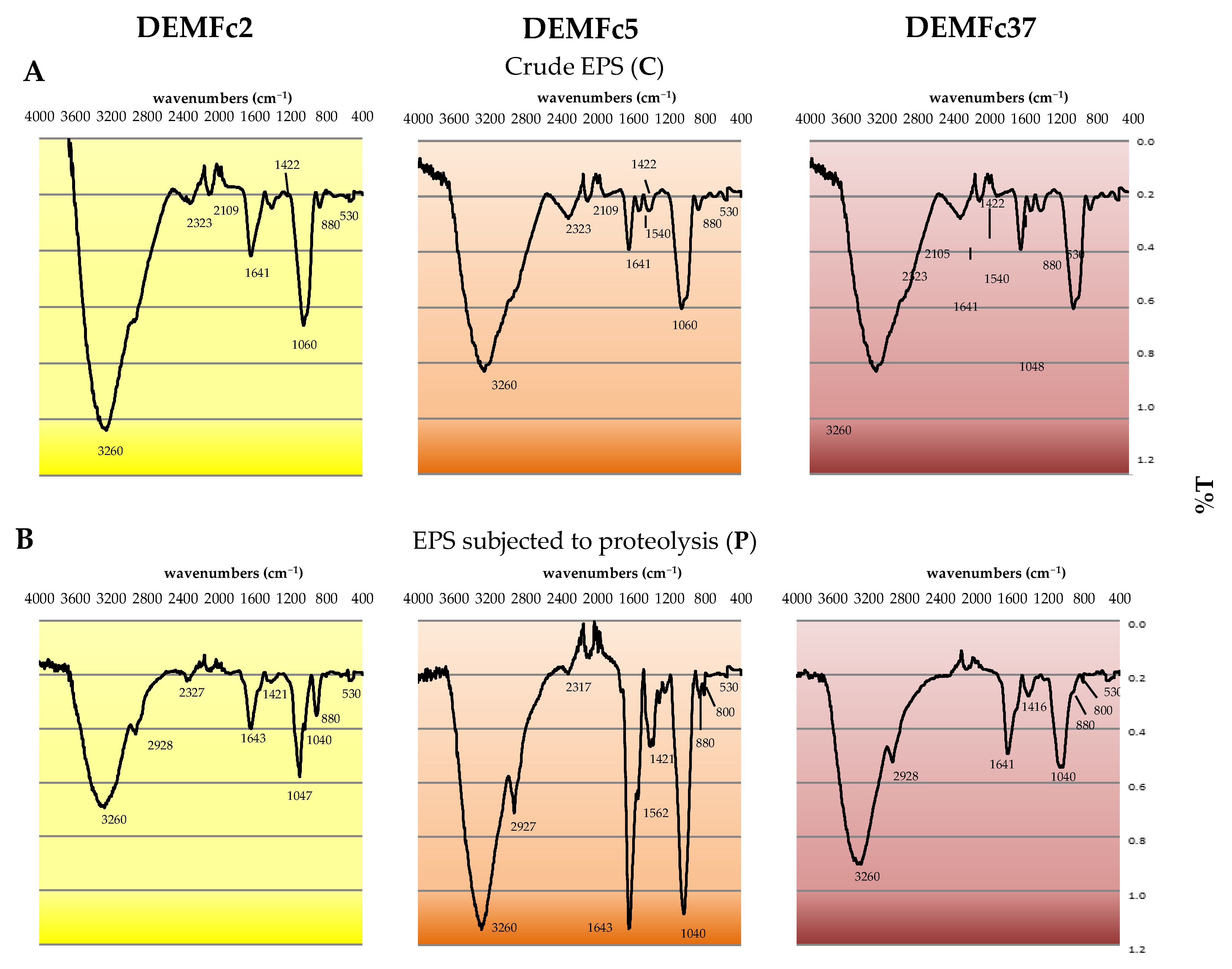
2.4.1. FTIR Spectra
2.4.2. Molecular Weights of EPS Polysaccharidic Fractions, Glycosidic Linkages
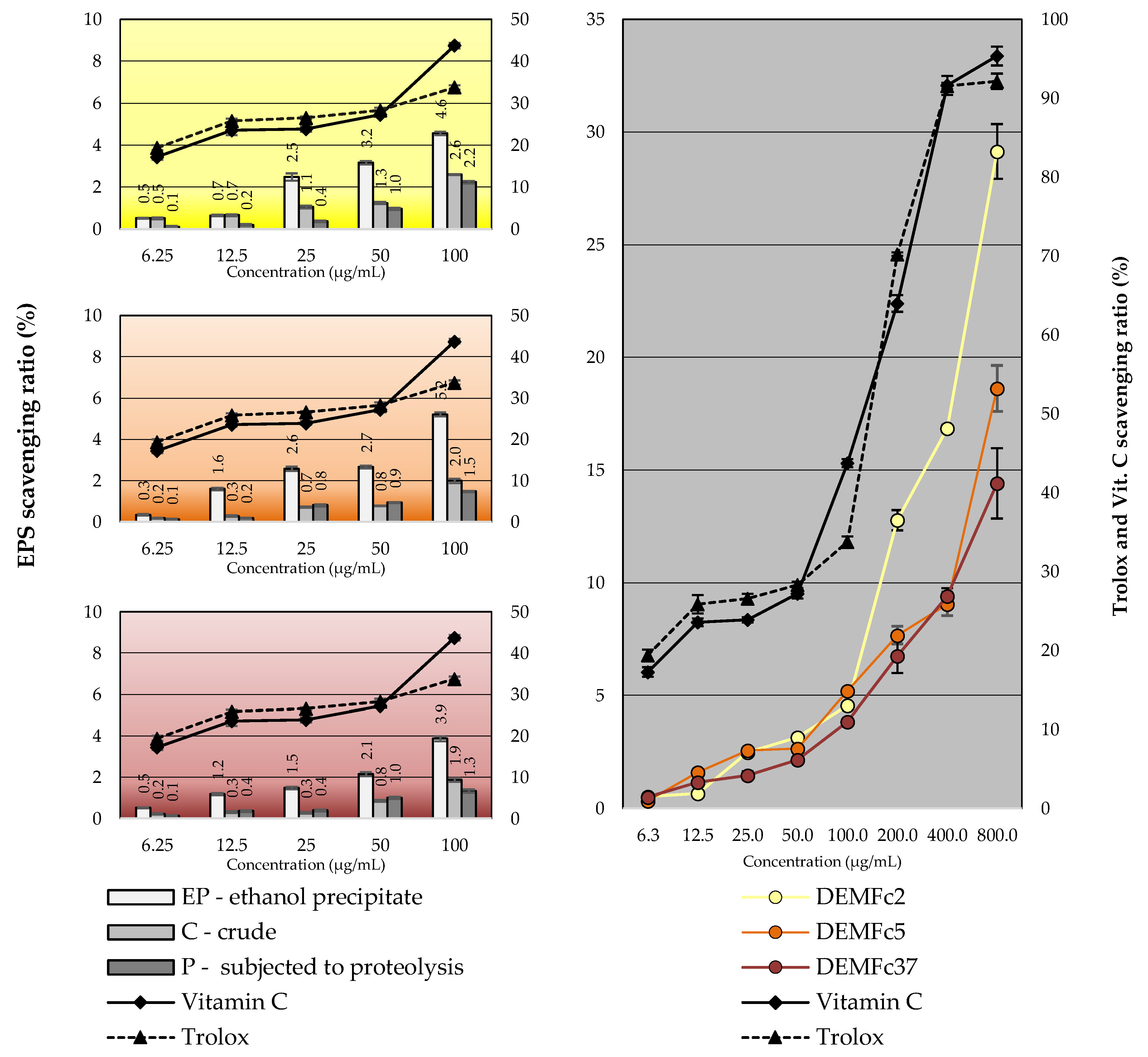
2.5. EPS Antioxidant Properties
3. Materials and Methods
3.1. Fungal Strains
3.2. Optimization of Fungal Culture
3.3. Isolation of Exopolymeric Substances (EPS)
3.4. Deproteinization of EPS
3.5. Determination of the Biochemical Composition in Water, Alkali, and Acid Solutions
3.5.1. Obtaining Aqueous, Alkaline, And Acidic EPS Fractions by Step Fractionation
3.5.2. Preparation of Aqueous, Alkaline, and Acidic EPS Solutions by Direct Dissolution of EPS in Solvents
3.5.3. Total Sugar
3.5.4. Protein
3.5.5. Phenolic Compounds
3.6. Visualization of Polysaccharides Using Confocal Microscopy
3.7. Gas Chromatography
3.8. Determination of the Size of Polymers in Preparations after Proteinase K Treatment/Molecular Weights of Polysaccharidic Subfractions of EPSs
3.9. FTIR Spectroscopy Analysis
3.10. Determination of the Presence of Glycosidic Bonds
3.11. Antioxidant Properties-ABTS Method
3.12. Statistical Analysis
4. Conclusions and Future Perspective
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| EPS | exopolymeric substances |
| DEM | department of environmental microbiology |
| PGPF | plant growth-promoting fungi |
| DRMO | deleterious rhizosphere microorganisms |
| kDa | kilodaltons |
| FTIR | infrared spectroscopy |
| PS | polymeric substances |
| UV | ultraviolet |
| DMSO | dimethyl sulfoxide |
| CWDE | cell wall degrading enzymes |
| PCA | principal component analysis |
| FTGM | fungal-types galactomannansfungal-type |
| Hex | hexose |
| Man | mannose |
| TPC | total phenolic content |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| ABTS | 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| Vit. C | vitamin C |
References
- Wingender, J.; Neu, T.R.; Flemming, H.-C. What are bacterial extracellular polymeric substances? In Microbial Extracellular Polymeric Substances; Springer: Berlin/Heidelberg, Germany, 1999; pp. 1–19. [Google Scholar]
- Selbmann, L.; Onofri, S.; Fenice, M.; Federici, F.; Petruccioli, M. Production and structural characterization of the exopolysaccharide of the Antarctic fungus Phoma herbarum CCFEE 5080. Res. Microbiol. 2002, 153, 585–592. [Google Scholar] [CrossRef]
- Ates, O. Systems biology of microbial exopolysaccharides production. Front. Bioeng. Biotech. 2015, 3, 200. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.A.; Kanekar, P.P. Production of exopolysaccharide by Vagococcus carniphilus MCM B-1018 isolated from alkaline Lonar Lake, India. Ann. Microbiol. 2011, 61, 733–740. [Google Scholar] [CrossRef]
- Freitas, F.; Torres, C.A.; Reis, M.A. Engineering aspects of microbial exopolysaccharide production. Bioresour. Technol. 2017, 245, 1674–1683. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.-J.; Yang, H.-R.; Ou, X.-Y.; Su, H.-H.; Zong, M.-H.; Yang, J.-G.; Lou, W.-Y. Fungal polysaccharide similar with host Dendrobium officinale polysaccharide: Preparation, structure characteristics and biological activities. Int. J. Biol. Macromol. 2019, 141, 460–475. [Google Scholar] [CrossRef] [PubMed]
- Nouha, K.; Kumar, R.S.; Balasubramanian, S.; Tyagi, R.D. Critical review of EPS production, synthesis and composition for sludge flocculation. J. Env. Sci. 2018, 66, 225–245. [Google Scholar] [CrossRef]
- Costa, O.Y.; Raaijmakers, J.M.; Kuramae, E.E. Microbial extracellular polymeric substances: Ecological function and impact on soil aggregation. Front. Microbiol. 2018, 9, 1636. [Google Scholar] [CrossRef]
- Czaczyk, K.; Myszka, K. Biosynteza egzopolisacharydów i ich rola w adhezji Bacillus megaterium do powierzchni stali nierdzewnej. Żywn. Nauka Technol. JakośćSupl. 2004, 11, 27–35. [Google Scholar]
- Górska, S.; Grycko, P.; Rybka, J.; Gamian, A. Egzopolisacharydy bakterii kwasu mlekowego–biosynteza i struktura Exopolysaccharides of lactic acid bacteria: Structure and biosynthesis. Postepy Hig. Med. Dosw. (Online) 2007, 61, 805–818. [Google Scholar]
- Donot, F.; Fontana, A.; Baccou, J.; Schorr-Galindo, S. Microbial exopolysaccharides: Main examples of synthesis, excretion, genetics and extraction. Carbohydr. Polym. 2012, 87, 951–962. [Google Scholar] [CrossRef]
- Pereira, G.A.; Arruda, H.S.; de Morais, D.R.; Eberlin, M.N.; Pastore, G.M. Carbohydrates, volatile and phenolic compounds composition, and antioxidant activity of calabura (Muntingia calabura L.) fruit. Food Res. Int. 2018, 108, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Xia, L.; Zhou, X.; Zhang, D.; Hu, C. Effects of light intensity on components and topographical structures of extracellular polysaccharides from the cyanobacteria Nostoc sp. J. Microbiol. 2014, 52, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Bonomi, H.R.; Posadas, D.M.; Paris, G.; del Carmen Carrica, M.D.C.; Frederickson, M.; Pietrasanta, L.I.; Bogomolni, R.A.; Zorreguieta, A.; Goldbaum, F.A. Light regulates attachment, exopolysaccharide production, and nodulation in Rhizobium leguminosarum through a LOV-histidine kinase photoreceptor. Proc. Natl. Acad. Sci. 2012, 109, 12135–12140. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, D.A.; Shcherba, V.V.; Bisko, N.A.; Poyedinok, N.L. Some biologically active substances from a mycelial biomass of medicinal caterpillar fungus Cordyceps sinensis (Berk.) Sacc.(Ascomycetes). Int. J. Med. Mushrooms. 2009, 11, 69–76. [Google Scholar] [CrossRef]
- Madla, S.; Methacanon, P.; Prasitsil, M.; Kirtikara, K. Characterization of biocompatible fungi-derived polymers that induce IL-8 production. Carbohydr. Polym. 2005, 59, 275–280. [Google Scholar] [CrossRef]
- Sardari, R.R.; Kulcinskaja, E.; Ron, E.Y.; Björnsdóttir, S.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó.; Karlsson, E.N. Evaluation of the production of exopolysaccharides by two strains of the thermophilic bacterium Rhodothermus marinus. Carbohydr. Polym. 2017, 156, 1–8. [Google Scholar] [CrossRef]
- Osińska-Jaroszuk, M.; Jarosz-Wilkołazka, A.; Jaroszuk-Ściseł, J.; Szałapata, K.; Nowak, A.; Jaszek, M.; Ozimek, E.; Majewska, M. Extracellular polysaccharides from Ascomycota and Basidiomycota: Production conditions, biochemical characteristics, and biological properties. WorldJ. Microb. Biot. 2015, 31, 1823–1844. [Google Scholar] [CrossRef]
- Rey, T.; Schornack, S. Interactions of beneficial and detrimental root-colonizing filamentous microbes with plant hosts. Genome Biol. 2013, 14, 121. [Google Scholar] [CrossRef]
- Jenkins, R.; Hall, J. Production and Applications of Microbial Exopolysaccharides; Butterworth Heinemann Limited: Oxford, UK, 1997. [Google Scholar]
- Mahapatra, S.; Banerjee, D. Fungal exopolysaccharide: Production, composition and applications. Microbiol. Insights 2013, 6, 1–16. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.P.; Heil, M. The microbe-free plant: Fact or artifact? Front. Plant. Sci. 2011, 2, 100. [Google Scholar] [CrossRef]
- Gientka, I.; Madejska, A. Ocena przydatności szczepów drożdży wyizolowanych z kefirów do syntezy polimerów zewnątrzkomórkowych. Zesz. Probl. Postepow. Nauk Roln. 2013, 574, 19–27. [Google Scholar]
- Güler, G.; Vorob’ev, M.M.; Vogel, V.; Mäntele, W. Proteolytically-induced changes of secondary structural protein conformation of bovine serum albumin monitored by Fourier transform infrared (FT-IR) and UV-circular dichroism spectroscopy. Spectrochim. Acta Part. A: Mol. Biomol. Spectrosc. 2016, 161, 8–18. [Google Scholar] [CrossRef]
- Pal, A.; Paul, A. Microbial extracellular polymeric substances: Central elements in heavy metal bioremediation. IndianJ. Microbiol. 2008, 48, 49. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Nimrichter, L.; Cordero, R.; Casadevall, A. Fungal polysaccharides: Biological activity beyond the usual structural properties. Front. Microbiol. 2011, 2, 171. [Google Scholar] [CrossRef]
- Wasser, S. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [CrossRef]
- Malinowska, E.; Krzyczkowski, W.; Herold, F. Pozyskiwanie, budowa i działanie biologiczne polisacharydów grzybowych na przykładzie soplówki jeżowatej (Hericium erinaceum). Biotechnologia 2008, 1, 109–121. [Google Scholar]
- Ustyuzhanina, N.E.; Kulakovskaya, E.V.; Kulakovskaya, T.V.; Menshov, V.M.; Dmitrenok, A.S.; Shashkov, A.S.; Nifantiev, N.E. Mannan and phosphomannan from Kuraishia capsulata yeast. Carbohydr. Polym. 2018, 181, 624–632. [Google Scholar] [CrossRef]
- Turkiewicz, M.; Czub, W. Kurdlan-struktura, wlaściwości, wykorzystanie. Biotechnologia 1997, 3, 16–26. [Google Scholar]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. DairyJ. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Rütering, M.; Schmid, J.; Gansbiller, M.; Braun, A.; Kleinen, J.; Schilling, M.; Sieber, V. Rheological characterization of the exopolysaccharide Paenan in surfactant systems. Carbohydr. Polym. 2018, 181, 719–726. [Google Scholar] [CrossRef]
- Rodrigues, M.; Nimrichter, L. In good company: Association between fungal glycans generates molecular complexes with unique functions. Front Microbiol. 2012, 3, 249. [Google Scholar] [CrossRef] [PubMed]
- Rühmann, B.; Schmid, J.; Sieber, V. Methods to identify the unexplored diversity of microbial exopolysaccharides. Front. Microbiol. 2015, 6, 565. [Google Scholar] [CrossRef] [PubMed]
- Wiater, A.; Pleszczyńska, M.; Szczodrak, J.; Próchniak, K. α-(1→ 3)-Glukany ściany komórkowej żółciaka siarkowego–Laetiporus sulphureus (Bull.: Fr.) Murrill–izolacja, charakterystyka i zastosowanie do indukcji syntezy. Biotechnologia 2008, 2, 174–189. [Google Scholar]
- Choma, A.; Nowak, K.; Komaniecka, I.; Waśko, A.; Pleszczyńska, M.; Siwulski, M.; Wiater, A. Chemical characterization of alkali-soluble polysaccharides isolated from a Boletus edulis (Bull.) fruiting body and their potential for heavy metal biosorption. Food Chem. 2018, 266, 329–334. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White Jr, J.F.W.; Arnold, A.E.; Redman, a.R.a. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Lilledahl, M.B.; Stokke, B.T. Novel imaging technologies for characterization of microbial extracellular polysaccharides. Front. Microbiol. 2015, 6, 525. [Google Scholar] [CrossRef][Green Version]
- Orlandelli, R.C.; Vasconcelos, A.F.D.; Azevedo, J.L.; da Silva, M.d.L.C.; Pamphile, J.A. Screening of endophytic sources of exopolysaccharides: Preliminary characterization of crude exopolysaccharide produced by submerged culture of Diaporthe sp. JF766998 under different cultivation time. Biochim.Open 2016, 2, 33–40. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Pu, H.; Liu, S.; Kan, J.; Jin, C. Recent advances in endophytic exopolysaccharides: Production, structural characterization, physiological role and biological activity. Carbohydr. Polym. 2017, 157, 1113–1124. [Google Scholar] [CrossRef]
- Toghueo, R.M.K. Bioprospecting endophytic fungi from Fusarium genus as sources of bioactive metabolites. Mycology 2019, 1–21. [Google Scholar] [CrossRef]
- Li, D.; Xi, H. Layered Extraction and Adsorption Performance of Extracellular Polymeric Substances from Activated Sludge in the Enhanced Biological Phosphorus Removal Process. Molecules 2019, 24, 3358. [Google Scholar] [CrossRef]
- Caicedo, N.H.; Davalos, A.F.; Puente, P.A.; Rodríguez, A.Y.; Caicedo, P.A. Antioxidant activity of exo-metabolites produced by Fusarium oxysporum: An endophytic fungus isolated from leaves of Otoba gracilipes. Microbiol. Open 2019, 8, n903. [Google Scholar] [CrossRef] [PubMed]
- Prathyusha, A.M.V.N.; Mohana Sheela, G.; Bramhachari, P.V. Chemical characterization and antioxidant properties of exopolysaccharides from mangrove filamentous fungi Fusarium equiseti ANP2. Biotechnol. Rep. 2018, 19, e00277. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Kloepper, J.W.; Ryu, C.-M. Rhizobacterial exopolysaccharides elicit induced resistance on cucumber. J. Microbiol. Biotechnol. 2008, 18, 1095–1100. [Google Scholar] [PubMed]
- Desjardins, A.; Proctor, R. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Lin, J.; Cao, J.; Zhang, P.; Bai, Y.; Chen, G.; Chen, K. An exopolysaccharide from Trichoderma pseudokoningii and its apoptotic activity on human leukemia K562 cells. Carbohydr. Polym. 2012, 89, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Kogan, G.; Matulová, M.; Michalková, E. Extracellular polysaccharides of Penicillium vermiculatum. Z. Nat. C 2002, 57, 452–458. [Google Scholar] [CrossRef]
- Camargo-de-Morais, M.; Ramos, S.; Pimentel, M.; de Morais Jr, M.; Lima Filho, J. Production of an extracellular polysaccharide with emulsifier properties by Penicillium citrinum. WorldJ. Microb. Biot. 2003, 19, 191–194. [Google Scholar] [CrossRef]
- Nehad, E.; El-Shamy, A. Physiological studies on the production of exopolysaccharide by fungi. Agric. Biol. J. N Am. 2010, 1, 1303–1308. [Google Scholar] [CrossRef]
- Siddiqui, I.; Adams, G. An extracellular polysaccharide from Gibberella fujikuroi (Fusarium moniliforme). Can. J. Chem. 1961, 39, 1683–1694. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Structural elucidation and bioactivity of a novel exopolysaccharide from endophytic Fusarium solani SD5. Carbohydr. Polym. 2012, 90, 683–689. [Google Scholar] [CrossRef]
- Mahapatra, S.; Banerjee, D. Optimization of a bioactive exopolysaccharide production from endophytic Fusarium solani SD5. Carbohydr. Polym. 2013, 97, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Mou, Y.; Shan, T.; Xu, J.; Li, Y.; Lu, S.; Zhou, L. Effects of polysaccharide elicitors from endophytic Fusarium oxysporium Dzf17 on growth and diosgenin production in cell suspension culture of Dioscorea zingiberensis. Molecules 2011, 16, 9003–9016. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Mao, W.; Li, Y.; Tian, J.; Xu, J. Structural elucidation of the exopolysaccharide produced by fungus Fusarium oxysporum Y24-2. Carbohydr. Res. 2013, 365, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Summerell, B.A.; Leslie, J.F. Fifty years of Fusarium: How could nine species have ever been enough? Fungal Divers. 2011, 50, 135. [Google Scholar] [CrossRef]
- Stępień, Ł.; Lalak-Kańczugowska, J.; Witaszak, N.; Urbaniak, M. Fusarium Secondary Metabolism Biosynthetic Pathways: So Close But So Far Away. Co-Evolution of Secondary Metabolites; Springer: Cham, Switzerland, 2018; pp. 1–37. [Google Scholar]
- Jaroszuk-Ściseł, J.; Kurek, E.; Winiarczyk, K.; Baturo, A.; Łukanowski, A. Colonization of root tissues and protection against Fusarium wilt of rye (Secale cereale) by nonpathogenic rhizosphere strains of Fusarium Culmorum. Biol. Control. 2008, 45, 297–307. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E.; Rodzik, B.; Winiarczyk, K. Interactions between rye (Secale cereale) root border cells (RBCs) and pathogenic and nonpathogenic rhizosphere strains of Fusarium culmorum. Mycol. Res. 2009, 113, 1053–1061. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Majewska, M.; Hanaka, A.; Tyśkiewicz, K.; Pawlik, A.; Janusz, G. Phytohormones (Auxin, Gibberellin) and ACC Deaminase In Vitro Synthesized by the Mycoparasitic Trichoderma DEMTkZ3A0 Strain and Changes in the Level of Auxin and Plant Resistance Markers in Wheat Seedlings Inoculated with this Strain Conidia. Int. J. Mol. Sci. 2019, 20, 4923. [Google Scholar] [CrossRef]
- Imazaki, I.; Kadota, I. Molecular phylogeny and diversity of Fusarium endophytes isolated from tomato stems. Fems Microbiol. Ecol. 2015, 91, fiv098. [Google Scholar] [CrossRef]
- Atanasoff, D.; Johnson, A.G. Treatment of cereal seeds by dry heat. J. Agric. Res. 1920, 18, 379–390. [Google Scholar]
- Jaroszuk-Ściseł, J.; Kurek, E.; Słomka, A.; Janczarek, M.; Rodzik, B. Activities of cell wall degrading enzymes in autolyzing cultures of three Fusarium culmorum isolates: Growth-promoting, deleterious and pathogenic to rye (Secale cereale). Mycologia 2011, 103, 929–945. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E. Hydrolysis of fungal and plant cell walls by enzymatic complexes from cultures of Fusarium isolates with different aggressiveness to rye (Secale cereale). Arch. Microbiol. 2012, 194, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Jaroszuk-Ściseł, J.; Kurek, E.; Trytek, M. Efficiency of indoleacetic acid, gibberellic acid and ethylene synthesized in vitro by Fusarium culmorum strains with different effects on cereal growth. Biologia 2014, 69, 281–292. [Google Scholar] [CrossRef]
- Lemanceau, P.; Alabouvette, C. Biological control of fusarium diseases by fluorescent Pseudomonas and non-pathogenic Fusarium. Crop. Prot. 1991, 10, 279–286. [Google Scholar] [CrossRef]
- Benhamou, N.; Garand, C. Cytological analysis of defense-related mechanisms induced in pea root tissues in response to colonization by nonpathogenic Fusarium oxysporum Fo47. Phytopathology 2001, 91, 730–740. [Google Scholar] [CrossRef]
- Zhong, L.; Niu, B.; Tang, L.; Chen, F.; Zhao, G.; Zhao, J. Effects of polysaccharide elicitors from endophytic Fusarium oxysporum Fat9 on the growth, flavonoid accumulation and antioxidant property of Fagopyrum tataricum sprout cultures. Molecules 2016, 21, 1590. [Google Scholar] [CrossRef]
- Yadav, R.; Tarafdar, J. Phytase and phosphatase producing fungi in arid and semi-arid soils and their efficiency in hydrolyzing different organic P compounds. Soil Biol. Biochem. 2003, 35, 745–751. [Google Scholar] [CrossRef]
- Guo, S.; Mao, W.; Yan, M.; Zhao, C.; Li, N.; Shan, J.; Lin, C.; Liu, X.; Guo, T.; Guo, T. Galactomannan with novel structure produced by the coral endophytic fungus Aspergillus ochraceus. Carbohydr. Polym. 2014, 105, 325–333. [Google Scholar] [CrossRef]
- Li, P.; Luo, H.; Meng, J.; Sun, W.; Wang, X.; Lu, S.; Peng, Y.; Zhou, L. Effects of oligosaccharides from endophytic Fusarium oxysporum Dzf17 on activities of defense-related enzymes in Dioscorea zingiberensis suspension cell and seedling cultures. Electron. J. Biotechn. 2014, 17, 156–161. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Mao, W.-J.; Tao, H.-W.; Zhu, W.-M.; Yan, M.-X.; Liu, X.; Guo, T.-T.; Guo, T. Preparation and characterization of a novel extracellular polysaccharide with antioxidant activity, from the mangrove-associated fungus Fusarium oxysporum. Mar. Biotechnol. 2015, 17, 219–228. [Google Scholar] [CrossRef]
- Barbosa, A.M.; Steluti, R.M.; Dekker, R.F.; Cardoso, M.S.; Da Silva, M.C. Structural characterization of Botryosphaeran: A (1→ 3; 1→ 6)-β-d-glucan produced by the ascomyceteous fungus, Botryosphaeria sp. Carbohydr. Res. 2003, 338, 1691–1698. [Google Scholar] [CrossRef]
- He, P.; Geng, L.; Mao, D.; Xu, C. Production, characterization and antioxidant activity of exopolysaccharides from submerged culture of Morchella crassipes. Bioproc. Biosyst. Eng. 2012, 35, 1325–1332. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, M.; Xin, Y.; Ma, D.; She, Z.; Wang, Z.; Sun, C.; Ren, Y. Effect of C/N ratio on extracellular polymeric substances of activated sludge from an anoxic–aerobic sequencing batch reactor treating saline wastewater. Env. Technol. 2014, 35, 2821–2828. [Google Scholar] [CrossRef]
- More, T.; Yadav, J.S.S.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Extracellular polymeric substances of bacteria and their potential environmental applications. J. Env. Manag. 2014, 144, 1–25. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.; Pang, X.; Yao, W.; Gao, X. Characterization and antioxidant activity of two low-molecular-weight polysaccharides purified from the fruiting bodies of Ganoderma lucidum. Int. J. Biol. Macromol. 2010, 46, 451–457. [Google Scholar] [CrossRef]
- Kim, S.S.; Lee, J.S.; Cho, J.Y.; Kim, Y.E.; Hong, E.K. Effects of C/N ratio and trace elements on mycelial growth and exo-polysaccharide production of Tricholoma matsutake. Biotechnol. Bioproc. E. 2010, 15, 293–298. [Google Scholar] [CrossRef]
- Yuan, B.; Chi, X.; Zhang, R. Optimization of exopolysaccharides production from a novel strain of Ganoderma lucidum CAU5501 in submerged culture. Braz. J. Microbiol. 2012, 43, 490–497. [Google Scholar] [CrossRef]
- Shi, T.; Aryantini, N.P.D.; Uchida, K.; Urashima, T.; Fukuda, K. Enhancement of Exopolysaccharide production of Lactobacillus fermentum TDS030603 by modifying culture conditions. Biosci. Microb. FoodH. 2014, 33, 85–90. [Google Scholar] [CrossRef]
- Ye, F.; Ye, Y.; Li, Y. Effect of C/N ratio on extracellular polymeric substances (EPS) and physicochemical properties of activated sludge flocs. J. Hazard. Mater. 2011, 188, 37–43. [Google Scholar] [CrossRef]
- Durmaz, B.; Sanin, F. Effect of carbon to nitrogen ratio on the composition of microbial extracellular polymers in activated sludge. Water Sci. Technol. 2001, 44, 221–229. [Google Scholar] [CrossRef]
- Yeh, C.-W.; Zang, C.-Z.; Lin, C.-C.; Kan, S.-C.; Chang, W.-F.; Shieh, C.-J.; Liu, Y.-C. Quantitative and morphologic analysis on exopolysaccharide and biomass production from a truffle endophytic fungus Hypocreales sp. NCHU01. J. Taiwan Insti. Chem. E. 2014, 45, 108–114. [Google Scholar] [CrossRef]
- Smirnou, D.; Hrubošová, D.; Kulhánek, J.; Švík, K.; Bobková, L.; Moravcová, V.; Krčmář, M.; Franke, L.; Velebný, V. Cryptococcus laurentii extracellular biopolymer production for application in wound management. App. Biochem. Biotech. 2014, 174, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mao, W.; Wang, J.; Zhu, W.; Zhao, C.; Li, N.; Wang, C.; Yan, M.; Guo, T.; Liu, X. Preparation and structural elucidation of a glucomannogalactan from marine fungus Penicillium commune. Carbohydr. Polym. 2013, 97, 293–299. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.d.L.C.; Fukuda, E.K.; Vasconcelos, A.F.D.; Dekker, R.F.; Matias, A.C.; Monteiro, N.K.; Cardoso, M.S.; Barbosa, A.M.; Silveira, J.L.; Sassaki, G.L. Structural characterization of the cell wall D-glucans isolated from the mycelium of Botryosphaeria rhodina MAMB-05. Carbohydr. Res. 2008, 343, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Oka, T. Biosynthesis of galactomannans found in filamentous fungi belonging to Pezizomycotina. Biosci. Biotech. Bioch. 2018, 82, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S.-i. A rapid permethylation of glycolipid, and polysaccharide catalyzed by methylsulfinyl carbanion in dimethyl sulfoxide. J. Biochem. 1964, 55, 205–208. [Google Scholar] [PubMed]
- Pan, F.; Su, T.-J.; Cai, S.-M.; Wu, W. Fungal endophyte-derived Fritillaria unibracteata var. wabuensis: Diversity, antioxidant capacities in vitro and relations to phenolic, flavonoid or saponin compounds. Sci. Rep. 2017, 7, 42008. [Google Scholar] [CrossRef] [PubMed]
- Kadiri, O. A review on the status of the phenolic compounds and antioxidant capacity of the flour: Effects of cereal processing. Int. J. Food Prop. 2017, 20, S798–S809. [Google Scholar] [CrossRef]
- Hanaka, A.; Nowak, A.; Plak, A.; Dresler, S.; Ozimek, E.; Jaroszuk-Ściseł, J.; Wójciak-Kosior, M.; Sowa, I. Bacterial isolate inhabiting spitsbergen soil modifies the physiological response of Phaseolus coccineus in control conditions and under exogenous application of methyl jasmonate and copper excess. J. Mol. Sci. 2019, 20, 1909. [Google Scholar] [CrossRef]
- Hameed, A.; Hussain, S.A.; Yang, J.; Ijaz, M.U.; Liu, Q.; Suleria, H.A.R.; Song, Y. Antioxidants potential of the filamentous fungi (Mucor circinelloides). Nutrients 2017, 9, 1101. [Google Scholar] [CrossRef]
- Guo, Y.-J.; Deng, G.-F.; Xu, X.-R.; Wu, S.; Li, S.; Xia, E.-Q.; Li, F.; Chen, F.; Ling, W.-H.; Li, H.-B. Antioxidant capacities, phenolic compounds and polysaccharide contents of 49 edible macro-fungi. Food Funct. 2012, 3, 1195–1205. [Google Scholar] [CrossRef]
- Saraswaty, V.; Srikandace, Y.; Simbiyani, N.; Setiyanto, H.; Udin, Z. Antioxidant activity and total phenolic content of endophytic fungus Fennellia nivea NRRL 5504. Pak. J. Biol. Sci. Pjbs 2013, 16, 1574–1578. [Google Scholar] [CrossRef] [PubMed]
- Druzian, S.P.; Pinheiro, L.N.; Susin, N.M.B.; Dal Prá, V.; Mazutti, M.A.; Kuhn, R.C.; de Marsillac Terra, L. Production of metabolites with antioxidant activity by Botryosphaeria dothidea in submerged fermentation. Bioprocess. Biosyst. Eng. 2020, 43, 13–20. [Google Scholar] [CrossRef]
- Mercado-Mercado, G.; Laura, A.; Alvarez-Parrilla, E. Effect of pectin on the interactions among phenolic compounds determined by antioxidant capacity. J. Mol. Struct. 2020, 1199, 126967. [Google Scholar] [CrossRef]
- Kurek, E.; Machowicz, Z.; Kulpa, D.; Slomka, A. The microorganisms of rye [Secale cereale L.] rhizosphere. Acta Microbiol. Pol. 1994, 2, 1053–1061. [Google Scholar]
- Czapek, F. Untersuchungen über die Stickstoffgewinnung. Hofmeisters Beiträge Zur Chem. Phys. U. Pathol. Bd. Ii 1902, 1, 559. [Google Scholar]
- Dox, A.W. The Intracellular Enzyms of Penicillium and Aspergillus: With Special Reference to Those of Penicillium camemberti; US Department of Agriculture, Bureau of Animal Industry: Washington, DC, USA, 1910; Volume 111.
- Komaniecka, I.; Choma, A.; Mazur, A.; Duda, K.A.; Lindner, B.; Schwudke, D.; Holst, O. Occurrence of an unusual hopanoid-containing lipid A among lipopolysaccharides from Bradyrhizobium species. J. Biol. Chem. 2014, 289, 35644–35655. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.t.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On tyrosine and tryptophane determinations in proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar]
- van den Berg, R.V.D.; Haenen, G.R.; van den Berg, H.V.D.; Bast, A. Applicability of an improved Trolox equivalent antioxidant capacity (TEAC) assay for evaluation of antioxidant capacity measurements of mixtures. Food Chem. 1999, 66, 511–517. [Google Scholar] [CrossRef]
- Sridhar, K.; Charles, A.L. In vitro antioxidant activity of Kyoho grape extracts in DPPH and ABTS assays: Estimation methods for EC50 using advanced statistical programs. Food Chem. 2019, 275, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Carlton, M.A.; Brandt, S. Data analysis: Statistical and computational methods for scientists and engineers. Am. Stat. 2000, 54, 155. [Google Scholar] [CrossRef]
Sample Availability: Not available. |


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavenumbers [cm−1] | Functional Groups Identified |
|---|---|
| 3260 | -OH; stretching vibrations of OH groups related to inter-residue hydrogen linkages |
| 2928 | C-H; stretching vibrations of aromatic and aliphatic C-H groups |
| 2320 | N-H groups; stretching vibrations of N-H groups from protein |
| 1643 | C=C ring carbons; stretching vibrations from the hexameric ring carbons (phenolic compounds or amino acids) |
| 1420 | -OH groups; stretching vibrations from alcohol -OH groups |
| 1040–1060 | -C-O- and -C-C- stretching vibrations and –C-OH- bending vibrations |
| 880 | the β-glycosidic linkages from sugars |
| 860 | C-H aryl; bending vibrations from aromatic rings |
| Peak No. | DEMFc2 | DEMFc5 | DEMFc37 |
|---|---|---|---|
| Mass (kDa) | Mass (kDa) | Mass (kDa) | |
| 1 | 1000–736 | 74 | 1000–736 |
| 2 | 16 | 34 | 19–12 |
| 3 | - | - | 5 |
| Ret. Time | Glycosidic Linkage | DEMFc2 | DEMFc5 | DEMFc37 |
|---|---|---|---|---|
| 11.14 | (terminal) 6-d-Hex(1→ | - | 3,4 | - |
| 12.51 | (terminal) Hex(1→(Man) | 21,5 | 17,5 | 50,4 |
| 14.40 | →2)Hex(1→ | Traces | 16,8 | 24,5 |
| 14.45 | →3)Hex I(1→ | 25,3 | 10,3 | - |
| 14.54 | →4)Hex(1→ | 53,2 | 29,0 | 5,5 |
| 14.88 | →3)Hex II(1→ | - | 3,1 | 7,2 |
| 15.10 | →6)Hex(1→ | - | 4,8 | - |
| 17.03 | →3,6)Hex(1→ | - | 15,2 | 12,3 |
| Scavenging Ratio (%) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration (µg/mL) | DEMFc2 | DEMFc5 | DEMFc37 | Vit. C | Trolox | ||||||
| EP | C | P | EP | C | P | EP | C | P | |||
| 6.25 | 0.52 ± 0.02 | 0.50 ± 0.04 | 0.10 ± 0.02 | 0.33 ± 0.03 | 0.20 ± 0.02 | 0.10 ± 0.03 | 0.52 ± 0.01 | 0.19 ± 0.02 | 0.10 ± 0.02 | 20.12 ± 0.68 | 19.13 ± 0.61 |
| 12.5 | 0.63 ± 0.03 | 0.66 ± 0.04 | 0.20 ± 0.03 | 1.59 ± 0.06 | 0.29 ± 0.03 | 0.20 ± 0.01 | 1.15 ± 0.04 | 0.29 ± 0.03 | 0.36 ± 0.04 | 24.30 ± 1.16 | 25.32 ± 0.49 |
| 25 | 2.50 ± 0.17 | 1.00 ± 0.06 | 0.36 ± 0.03 | 2.55 ± 0.11 | 0.69 ± 0.03 | 0.85 ± 0.05 | 1.40 ± 0.05 | 0.30 ± 0.03 | 0.40 ± 0.04 | 25.77 ± 0.64 | 26.40 ± 0.32 |
| 50 | 2.25 ± 0.08 | 1.20 ± 0.05 | 0.95 ± 0.04 | 2.58 ± 0.06 | 0.79 ± 0.04 | 0.96 ± 0.02 | 2.03 ± 0.09 | 0.85 ± 0.04 | 0.98 ± 0.05 | 27.40 ± 0.41 | 27.70 ± 0.65 |
| 100 | 2.94 ± 0.09 | 2.60 ± 0.02 | 2.20 ± 0.06 | 3.08 ± 0.02 | 1.90 ± 0.06 | 1.50 ± 0.02 | 2.99 ± 0.11 | 1.85 ± 0.09 | 1.36 ± 0.07 | 42.8 ± 0.67 | 33.90 ± 0.51 |
| 200 | 12.77 ± 0.75 | IS | IS | 7.68 ± 0.39 | IS | IS | 6.75 ± 0.44 | IS | IS | 63.98 ± 0.20 | 70.23 ± 1.05 |
| 400 | 16.85 ± 0.35 | IS | IS | 9.06 ± 0.52 | IS | IS | 9.41 ± 0.15 | IS | IS | 91.62 ± 0.39 | 91.55 ± 1.20 |
| 800 | 29.14 ± 1.55 | IS | IS | 18.62 ± 1.02 | IS | IS | 14.41 ± 1.21 | IS | IS | 95.35 ± 0.97 | 92.14 ± 1.19 |
| R2 | +0.629 | +0.791 | +0.654 | +0.664 | +0.75 | +0.886 | +0.721 | +0.676 | +0.851 | +0.874 | +0.840 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaroszuk-Ściseł, J.; Nowak, A.; Komaniecka, I.; Choma, A.; Jarosz-Wilkołazka, A.; Osińska-Jaroszuk, M.; Tyśkiewicz, R.; Wiater, A.; Rogalski, J. Differences in Production, Composition, and Antioxidant Activities of Exopolymeric Substances (EPS) Obtained from Cultures of Endophytic Fusarium culmorum Strains with Different Effects on Cereals. Molecules 2020, 25, 616. https://doi.org/10.3390/molecules25030616
Jaroszuk-Ściseł J, Nowak A, Komaniecka I, Choma A, Jarosz-Wilkołazka A, Osińska-Jaroszuk M, Tyśkiewicz R, Wiater A, Rogalski J. Differences in Production, Composition, and Antioxidant Activities of Exopolymeric Substances (EPS) Obtained from Cultures of Endophytic Fusarium culmorum Strains with Different Effects on Cereals. Molecules. 2020; 25(3):616. https://doi.org/10.3390/molecules25030616
Chicago/Turabian StyleJaroszuk-Ściseł, Jolanta, Artur Nowak, Iwona Komaniecka, Adam Choma, Anna Jarosz-Wilkołazka, Monika Osińska-Jaroszuk, Renata Tyśkiewicz, Adrian Wiater, and Jerzy Rogalski. 2020. "Differences in Production, Composition, and Antioxidant Activities of Exopolymeric Substances (EPS) Obtained from Cultures of Endophytic Fusarium culmorum Strains with Different Effects on Cereals" Molecules 25, no. 3: 616. https://doi.org/10.3390/molecules25030616
APA StyleJaroszuk-Ściseł, J., Nowak, A., Komaniecka, I., Choma, A., Jarosz-Wilkołazka, A., Osińska-Jaroszuk, M., Tyśkiewicz, R., Wiater, A., & Rogalski, J. (2020). Differences in Production, Composition, and Antioxidant Activities of Exopolymeric Substances (EPS) Obtained from Cultures of Endophytic Fusarium culmorum Strains with Different Effects on Cereals. Molecules, 25(3), 616. https://doi.org/10.3390/molecules25030616