
Sulphamoylated Estradiol Analogue Induces Reactive Oxygen Species Generation to Exert Its Antiproliferative Activity in Breast Cancer Cell Lines


Abstract
1. Introduction
2. Results
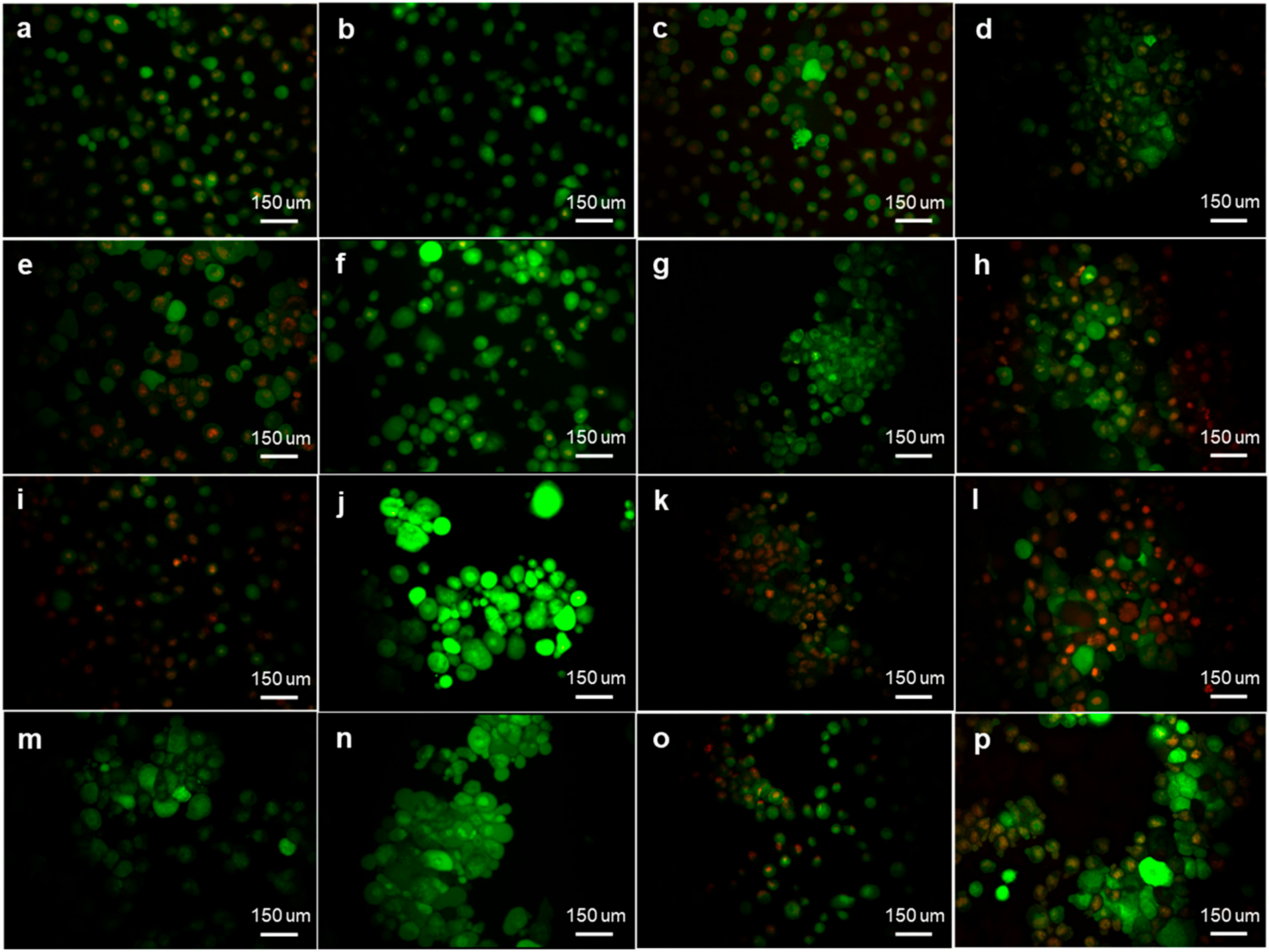
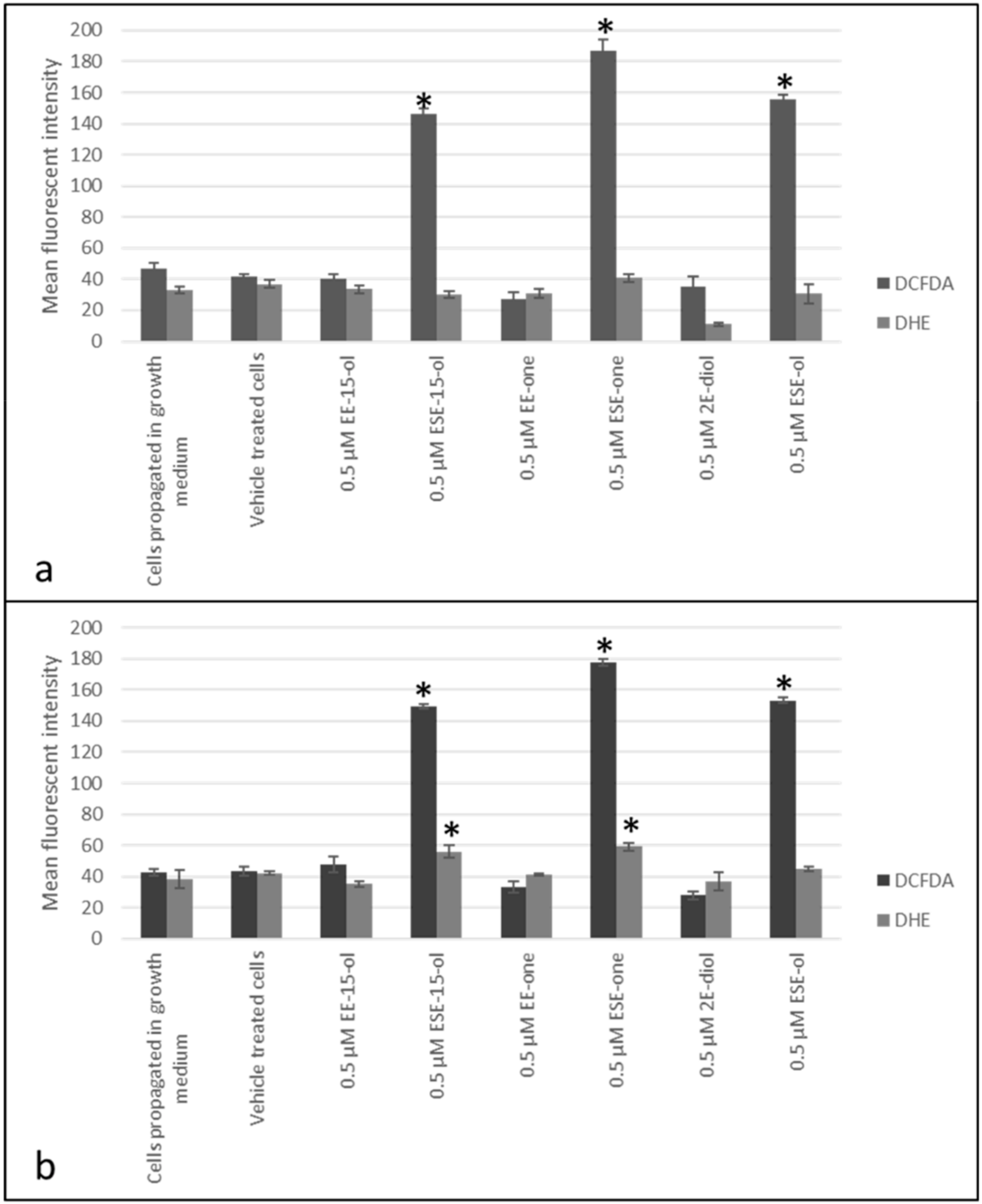
2.1. Sulphamoylated Compounds Induce Hydrogen Peroxide and Superoxide Anion
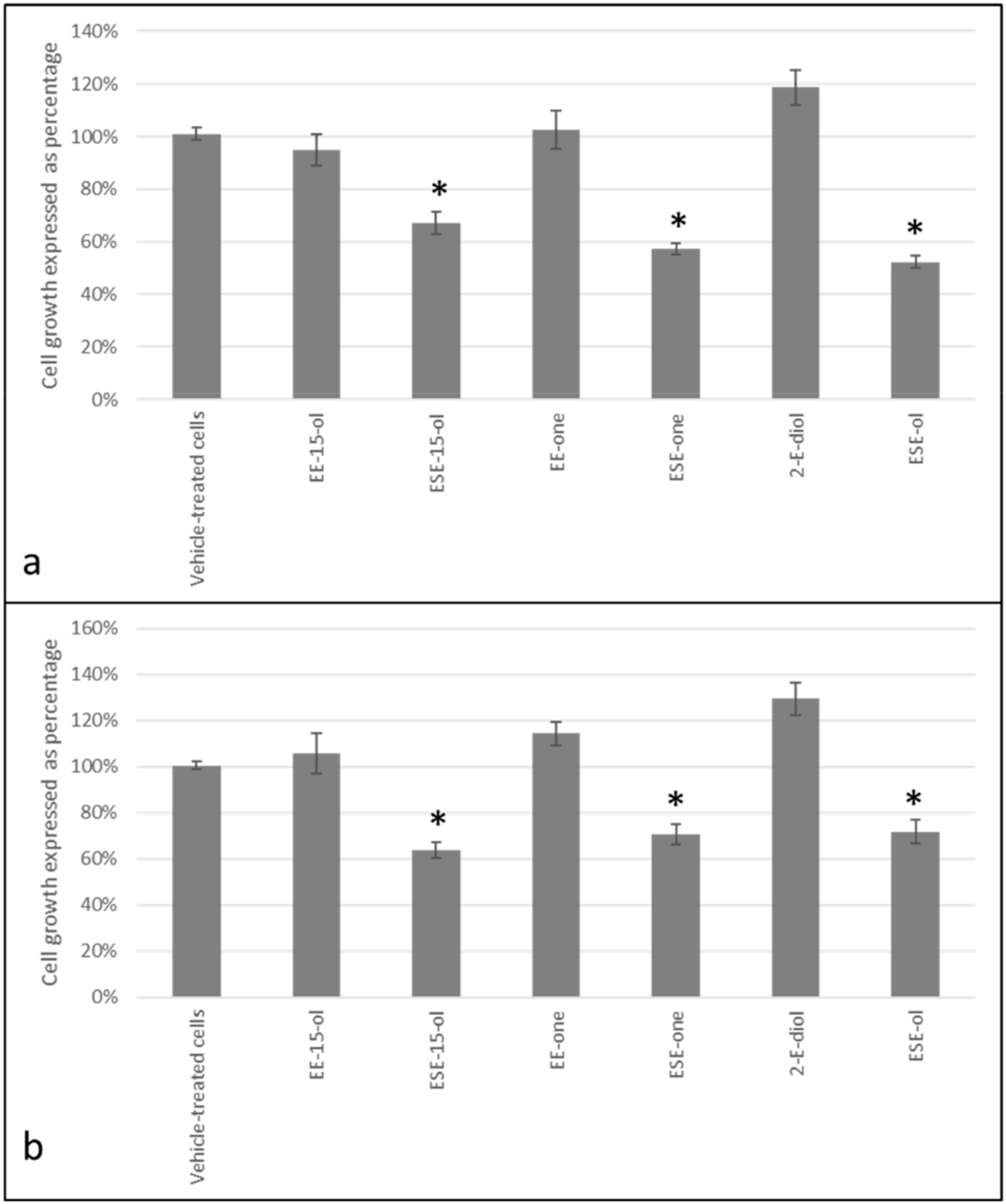
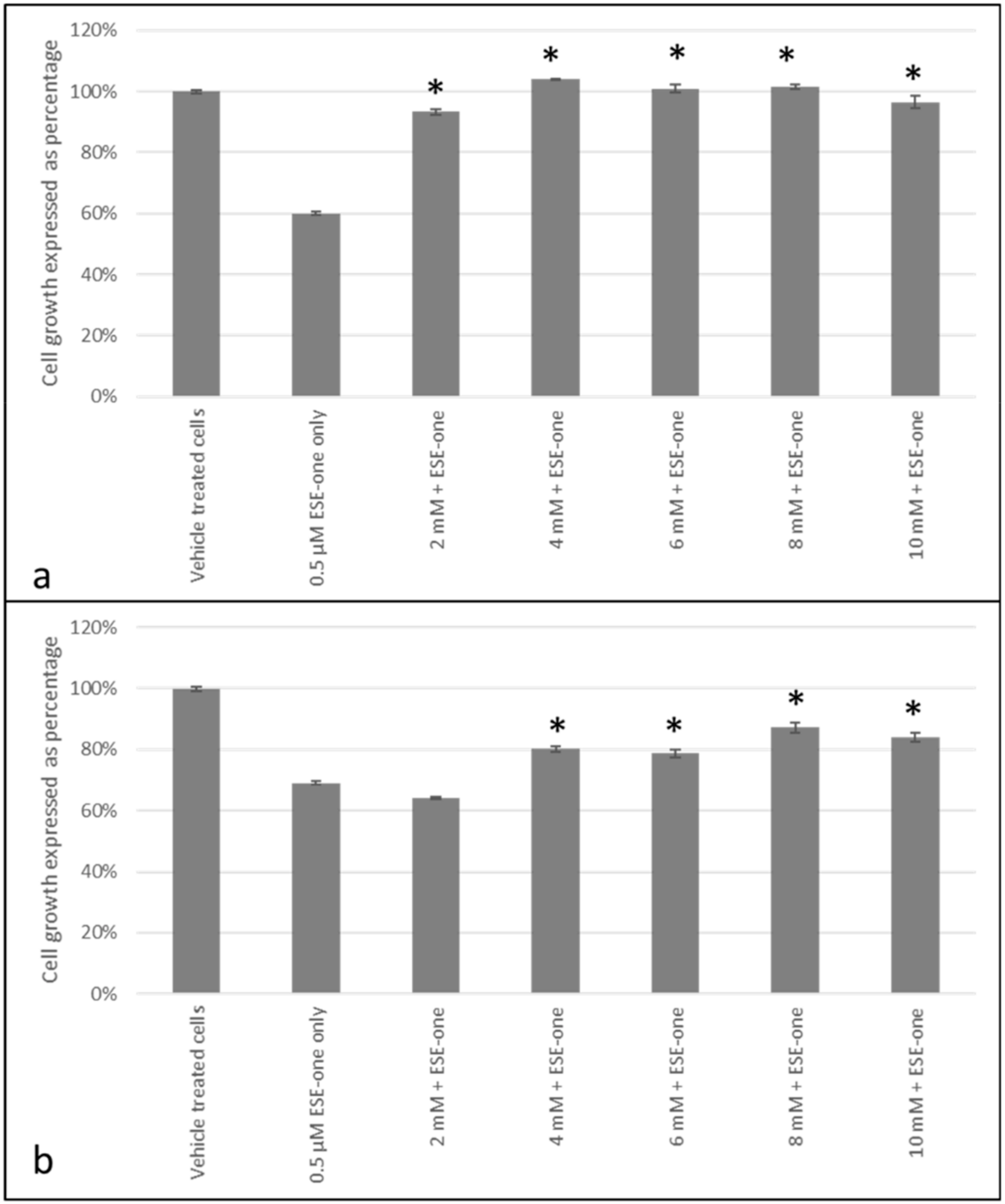
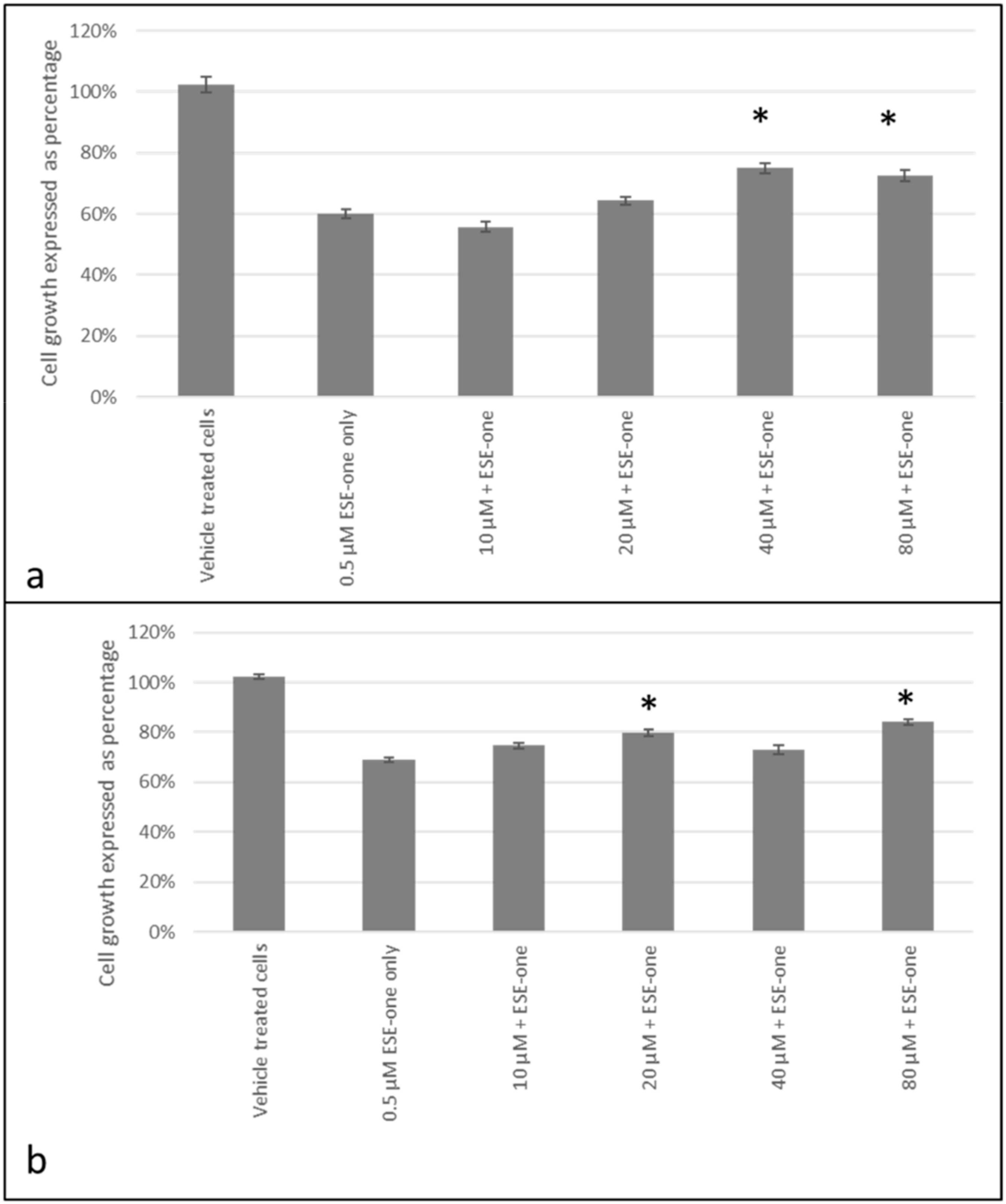
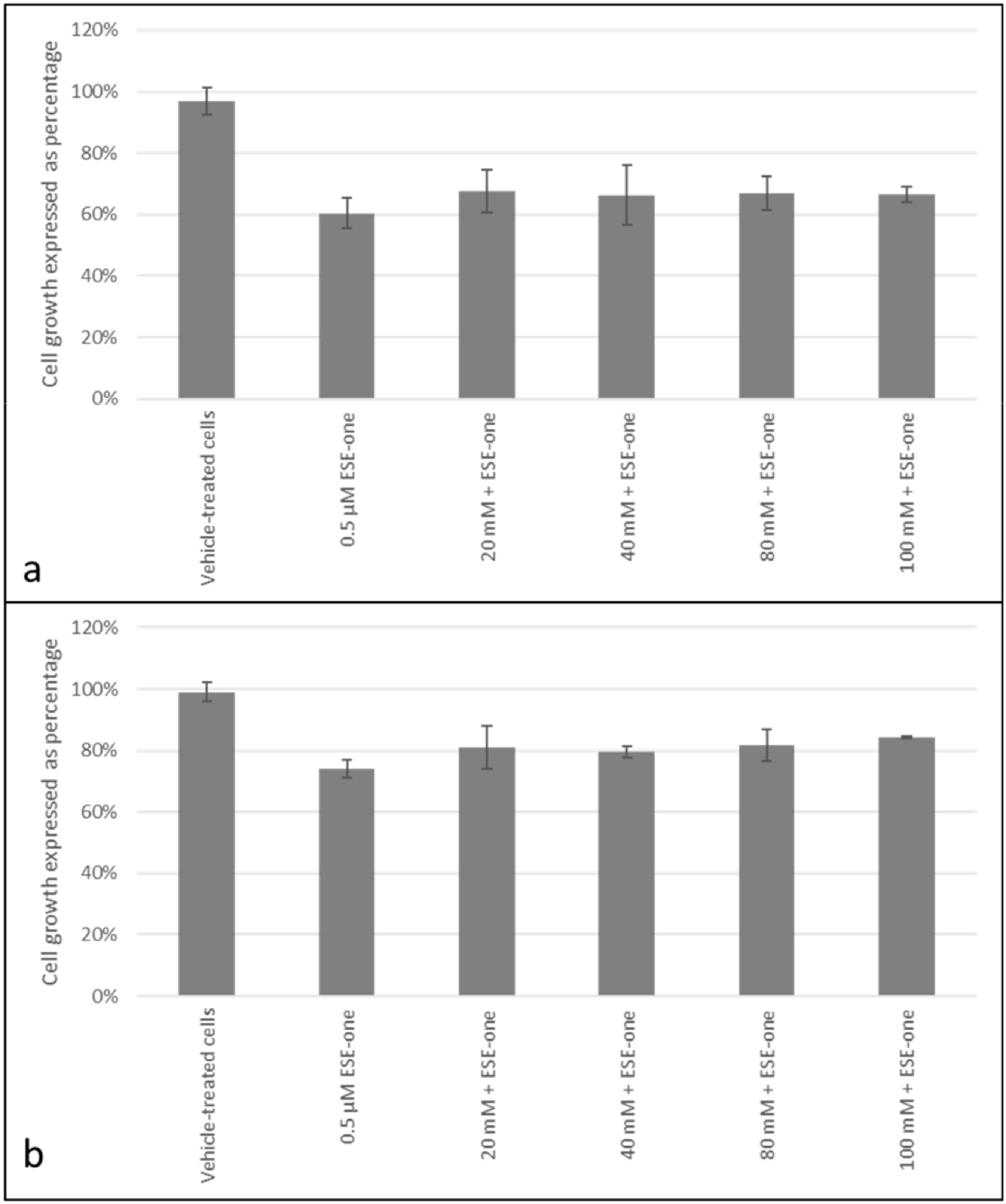
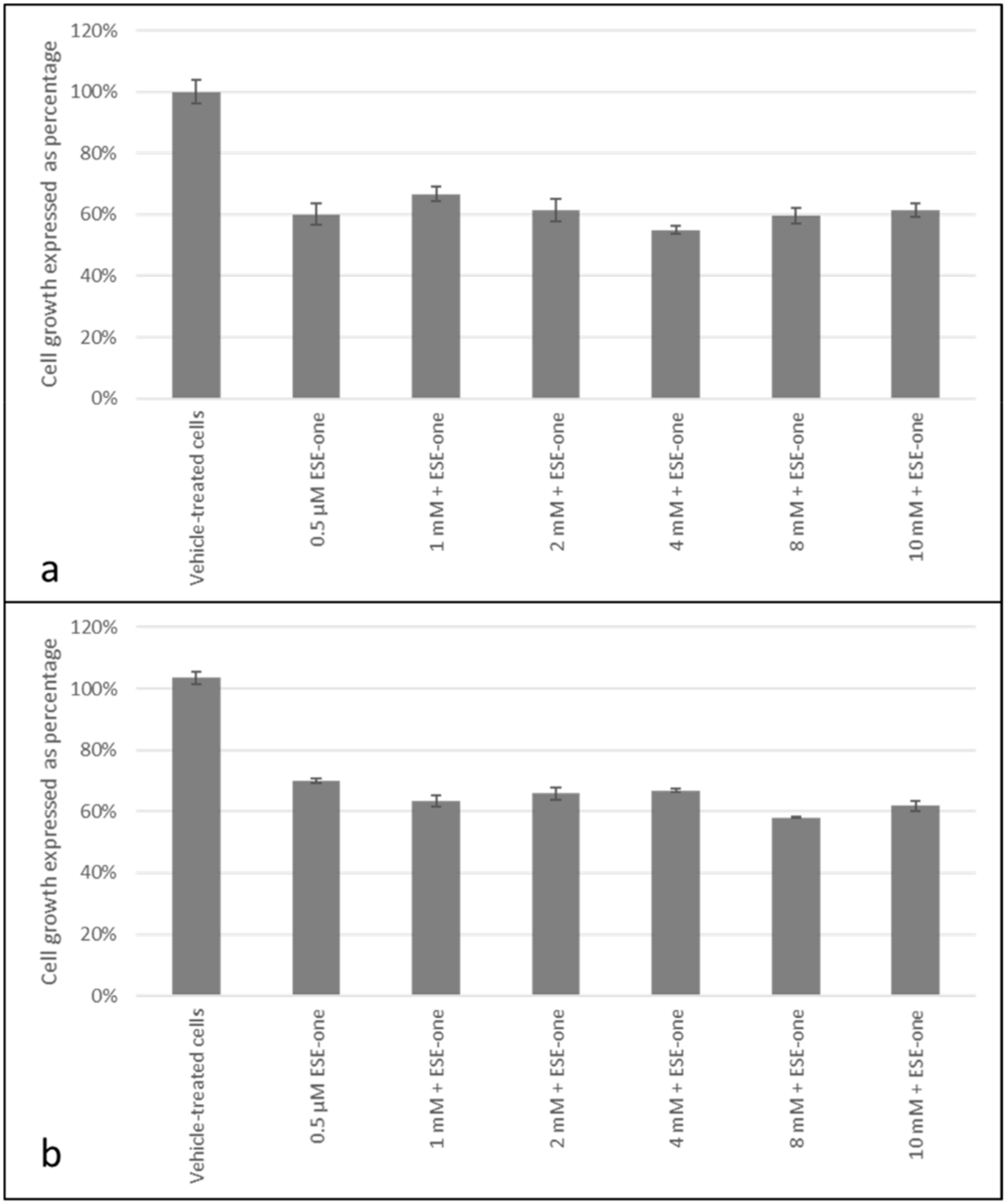
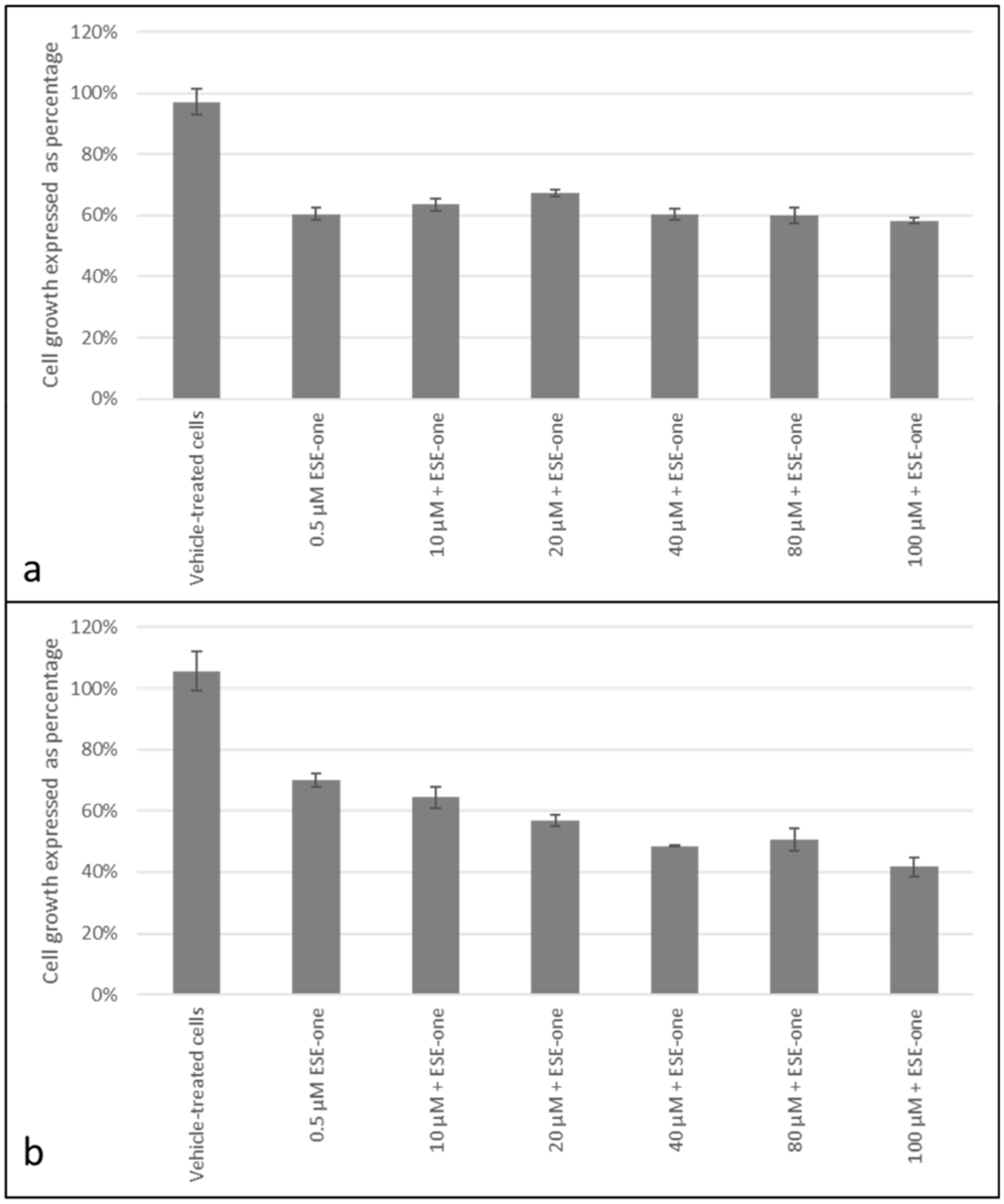
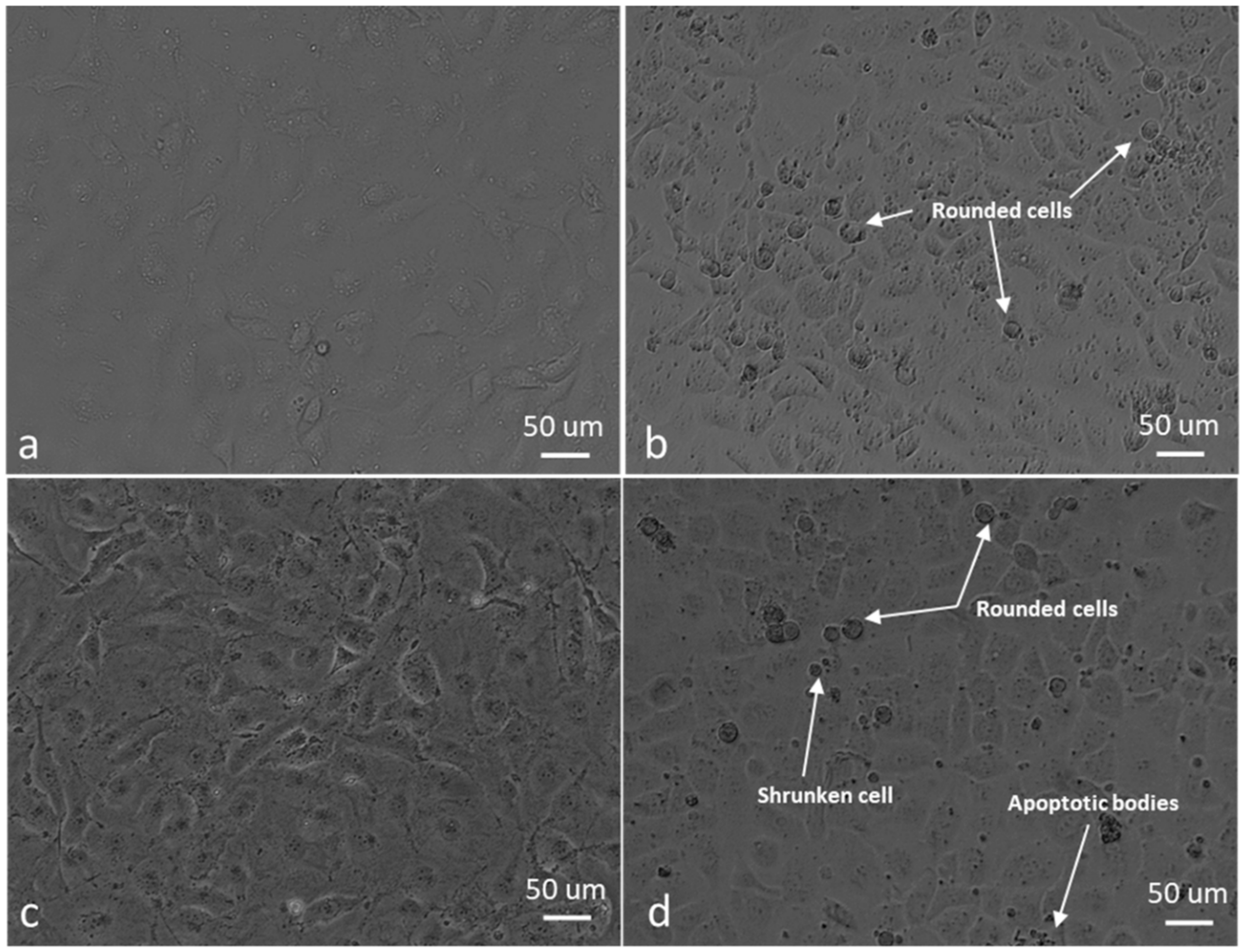
2.2. Decline in Cell Growth Owed to Sulphamoylated Compounds
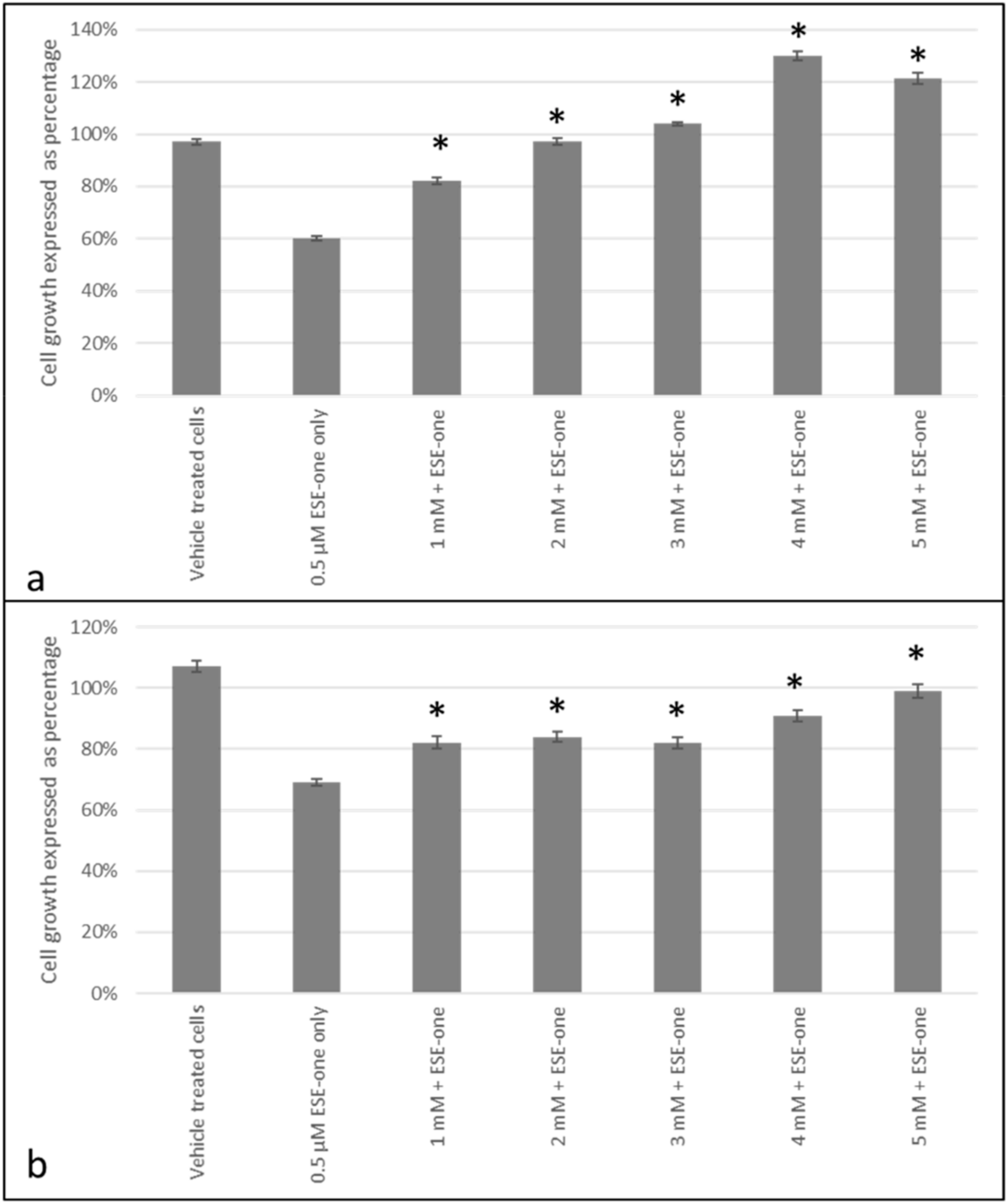
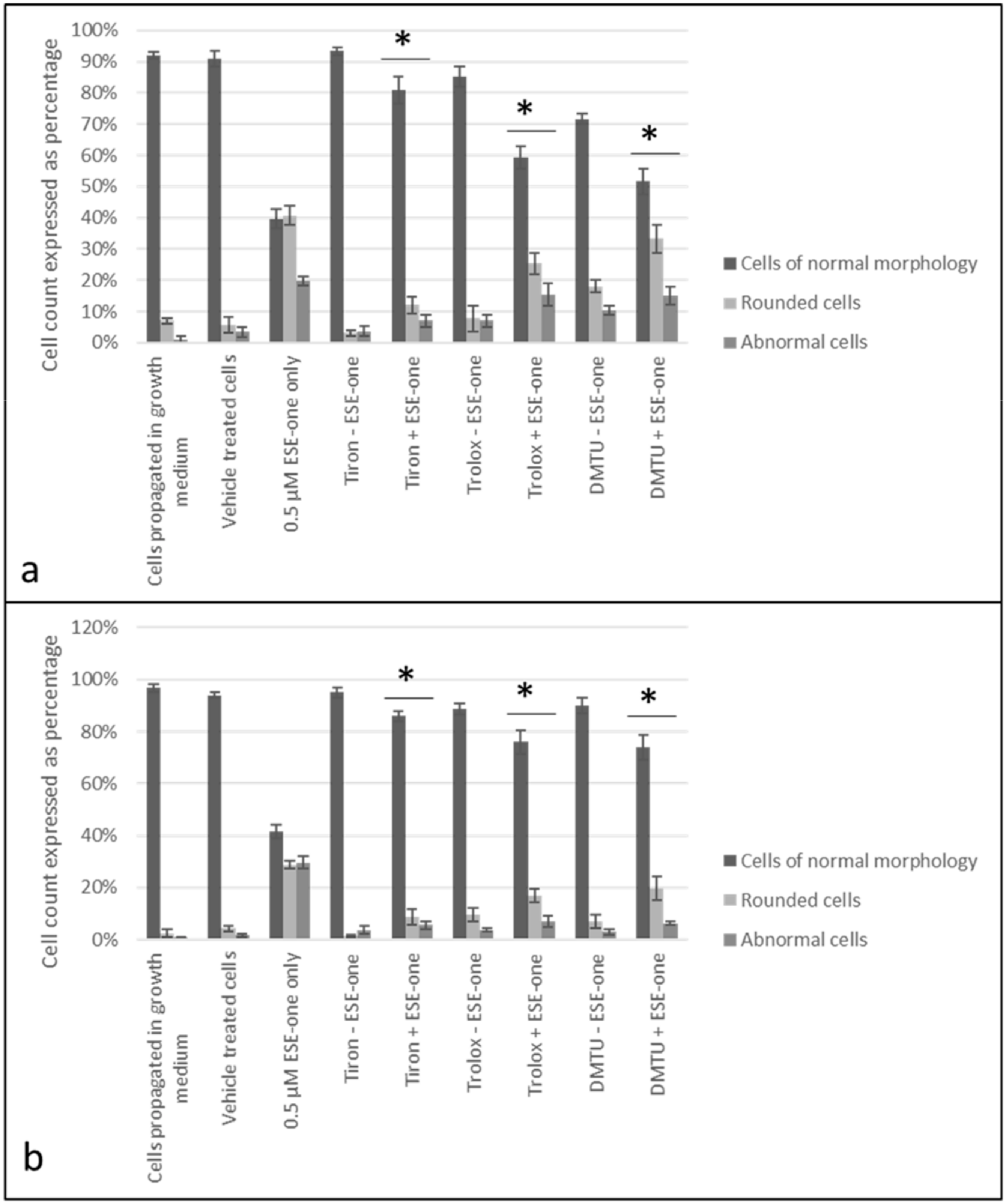
2.3. ROS Scavengers Oppose the Antiproliferative Effects of Sulphamoylated Compounds (ESE-One)
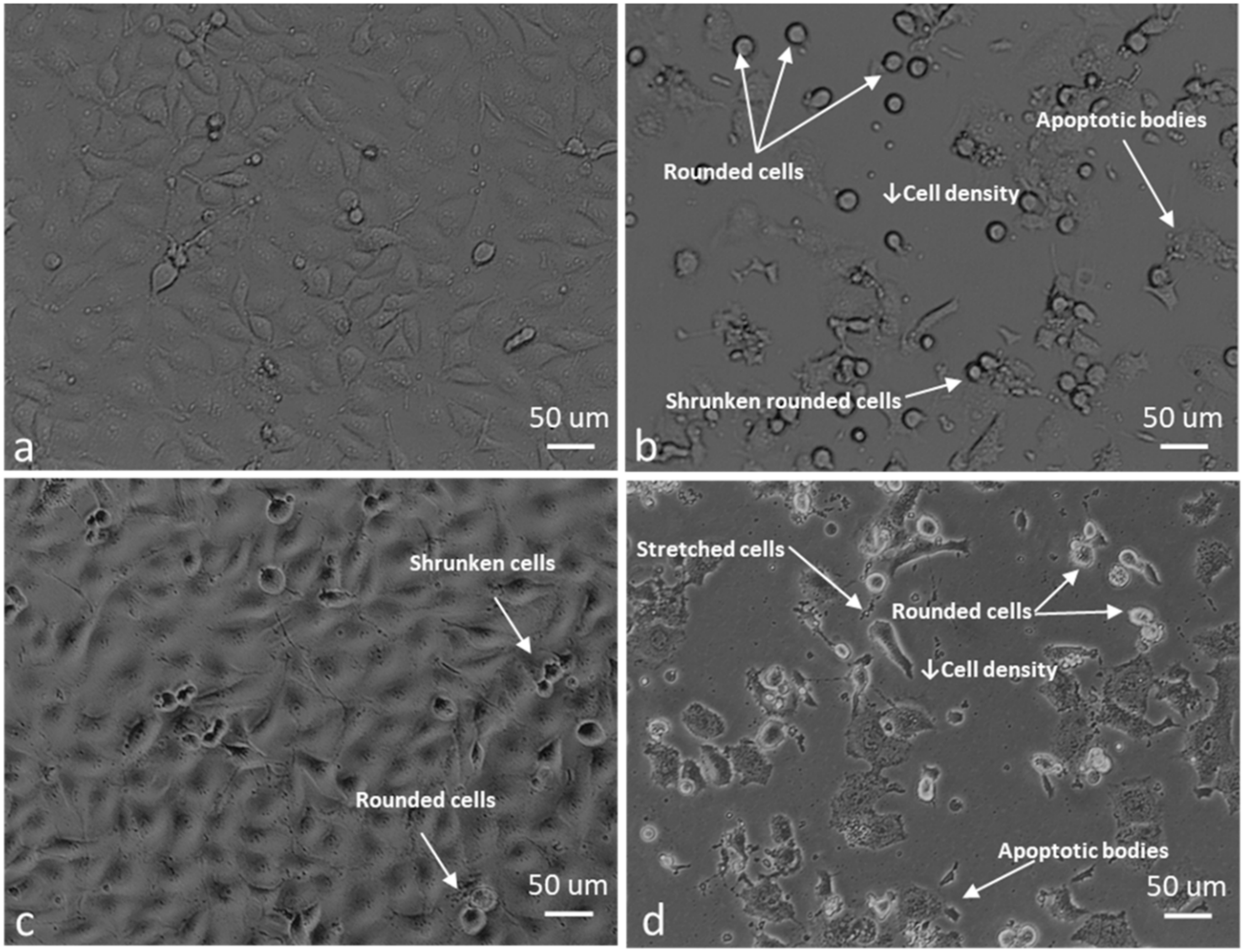
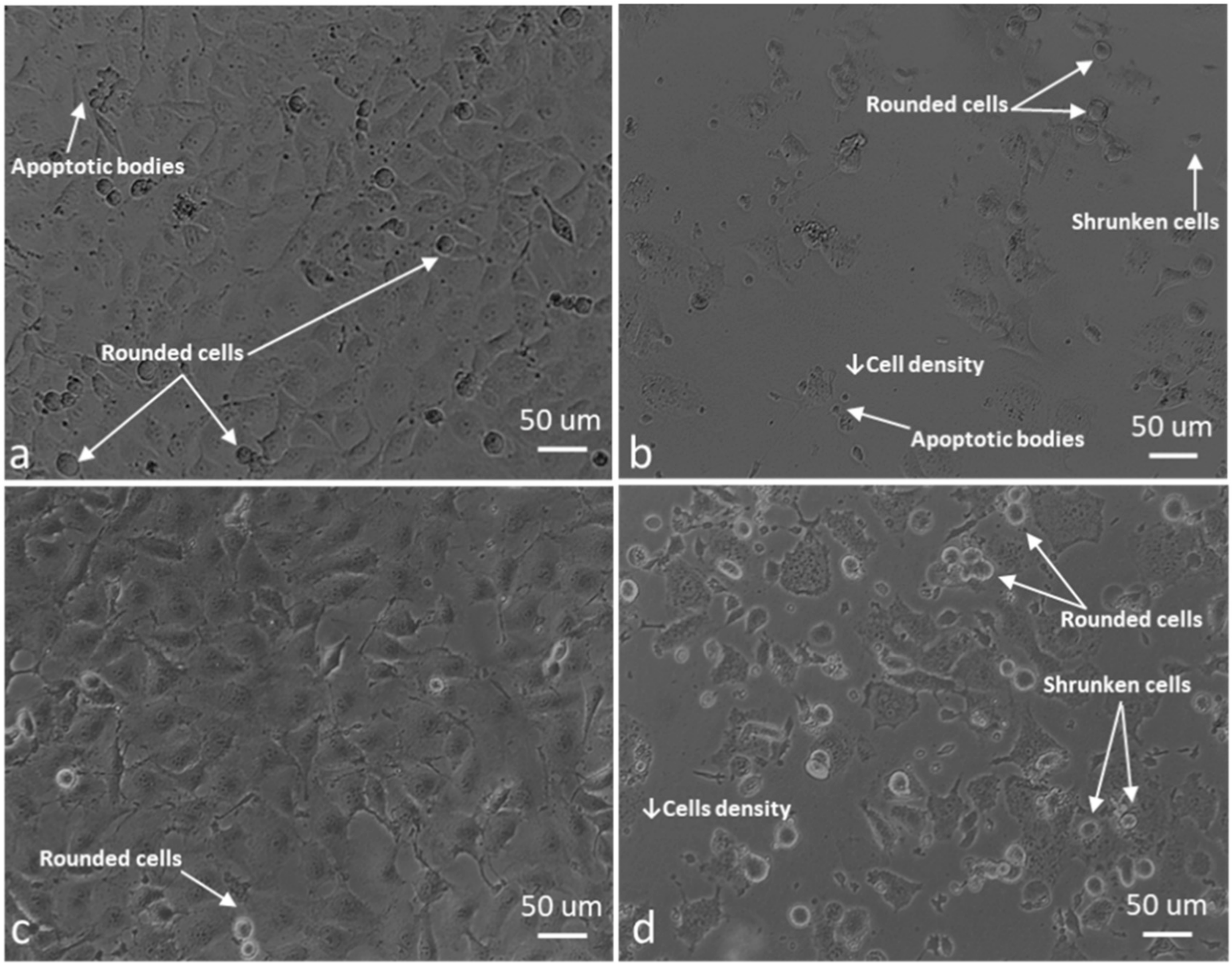
2.4. ROS Scavengers Oppose the Cell Rounding Effect of ESE-One
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Reagents
4.3. Methods
4.3.1. Hydrogen Peroxide and Superoxide Production (Fluorescent Microscopy)
4.3.2. Cell Proliferation (Spectrophotometry)
4.3.3. Cell Morphology
5. Statistics
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Visagie, M.H.; Joubert, A.M. In vitro effects of 2-methoxyestradiol-bis-sulphamate on reactive oxygen species and possible apoptosis induction in a breast adenocarcinoma cell line. Cancer Cell Int. 2011, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Visagie, M.H.; Mqoco, T.; Joubert, A. Sulphamoylated estradiol analogue induces antiproliferative activity and apoptosis in breast cell lines. Cell. Mol. Biol. Lett. 2012, 17, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Visagie, M.H.; Theron, A.; Mqoco, T.; Vieira, W.A.; Prudent, R.; Martinez, A.; Lafanechère, L.; Joubert, A. Sulphamoylated 2-Methoxyestradiol Analogues Induce Apoptosis in Adenocarcinoma Cell Lines. PLoS ONE 2013, 8, e71935. [Google Scholar] [CrossRef] [PubMed]
- Stander, A.; Joubert, F.; Joubert, A. Docking, Synthesis, and in vitro Evaluation of Antimitotic Estrone Analogs. Chem. Biol. Drug Des. 2011, 77, 173–181. [Google Scholar] [CrossRef]
- Visagie, M.H.; Birkholtz, L.M.; Joubert, A. 17-beta-estradiol analog inhibits cell proliferation by induction of apoptosis in breast cell lines. Microsc. Res. Tech. 2014, 77, 236–242. [Google Scholar] [CrossRef]
- Wolmarans, E.; Mqoco, T.; Stander, A.; Nkandeu, S.D.; Sippel, K.; McKenna, R.; Joubert, A. Novel estradiol analogue induces apoptosis and autophagy in esophageal carcinoma cells. Cell. Mol. Biol. Lett. 2014, 19, 98–115. [Google Scholar] [CrossRef]
- Visagie, M.H.; Birkholtz, L.M.; Joubert, A. A 2-methoxyestradiol bis-sulphamoylated derivative induces apoptosis in breast cell lines. Cell Biosci. 2015, 5, 19. [Google Scholar] [CrossRef]
- Visagie, M.H.; van der Bout, I.; Joubert, A. A bis-sulphamoylated estradiol derivative induces ROS-dependent cell cycle abnormalities and subsequent apoptosis. PLoS ONE 2017, 12, e176006. [Google Scholar] [CrossRef]
- Liou, G.Y.; Storz, P. Detecting reactive oxygen species by immunohistochemistry. Methods Mol. Biol. 2015, 1292, 97–104. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Glasauer, A.; Chandel, N.S. Targeting antioxidants for cancer therapy. Biochem. Pharmacol. 2014, 92, 90–101. [Google Scholar] [CrossRef]
- Diebold, L.; Chandel, N.S. Mitochondrial ROS regulation of proliferating cells. Free. Radic. Biol. Med. 2016, 100, 86–93. [Google Scholar] [CrossRef]
- Prasad, S.; Gupta, S.C.; Tyagi, A.K. Reactive oxygen species (ROS) and cancer: Role of antioxidative nutraceuticals. Cancer Lett. 2017, 387, 95–105. [Google Scholar] [CrossRef]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. 2011, 13, 215. [Google Scholar] [CrossRef] [PubMed]
- Stander, B.; Marais, S.; Vorster, C.; Joubert, A. In vitro effects of 2-methoxyestradiol on morphology, cell cycle progression, cell death and gene expression changes in the tumorigenic MCF-7 breast epithelial cell line. J. Steroid Biochem. Mol. Biol. 2010, 119, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Myhre, O.; Andersen, J.M.; Aarnes, H.; Fonnum, F. Evaluation of the probes 2’,7’-dichlorofluorescin diacetate, luminol, and lucigenin as indicators of reactive species formation. Biochem. Pharmacol. 2003, 65, 1575–1582. [Google Scholar] [CrossRef]
- Zhao, H.; Kalivendi, S.; Zhang, H.; Joseph, J.; Nithipatikom, K.; Vásquez-Vivar, J.; Kalyanaraman, B. Superoxide reacts with hydroethidine but forms a fluorescent product that is distinctly different from ethidium: Potential implications in intracellular fluorescence detection of superoxide. Free Radic. Biol. Med. 2003, 34, 1359–1368. [Google Scholar] [CrossRef]
- Sullivan, L.B.; Chandel, N.S. Mitochondrial reactive oxygen species and cancer. Cancer Metab. 2014, 2, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Marchi, S.; Giorgi, C.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Missiroli, S.; Patergnani, S.; Poletti, F.; et al. Mitochondria-Ros Crosstalk in the Control of Cell Death and Aging. J. Signal Transduct. 2012, 2012, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (Δψ m) in apoptosis; An update. Apoptosis 2003, 8, 115–128. [Google Scholar] [CrossRef]
- Li, P.; Dietz, R.; von Harsdorf, R. p53 regulates mitochondrial membrane potential through reactive oxygen species and induces cytochrome c-independent apoptosis blocked by Bcl-2. EMBO J. 1999, 18, 6027–6036. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. BBA Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Marais, S.; Mqoco, T.V.; Stander, A.; Van Papendorp, D.H.; Joubert, A.M. The in vitro effects of a sulphamoylated derivative of 2-methoxyestradiol on cell number, morphology and alpha-Tubulin disruption in cervical adenocarcinoma (HeLa) cells. Biomed. Res. 2012, 23, 357–362. [Google Scholar]
- Franco, R.; Panayiotidis, M.I.; Cidlowski, J.A. Glutathione Depletion Is Necessary for Apoptosis in Lymphoid Cells Independent of Reactive Oxygen Species Formation. J. Biol. Chem. 2007, 282, 30452–30465. [Google Scholar] [CrossRef] [PubMed]
- Bhowmick, R.; Girotti, A.W. Cytoprotective induction of nitric oxide synthase in a cellular model of 5-aminolevulinic acid-based photodynamic therapy. Free Radic. Biol. Med. 2010, 48, 1296–1301. [Google Scholar] [CrossRef] [PubMed]
- Drummen, G.P.; Makkinje, M.; Verkleij, A.J.; den Kamp, J.A.O.; Post, J.A. Attenuation of lipid peroxidation by antioxidants in rat-1 fibroblasts: Comparison of the lipid peroxidation reporter molecules cis-parinaric acid and C11-BODIPY 581/591 in a biological setting. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2004, 1636, 136–150. [Google Scholar] [CrossRef]
- Nicolescu, A.C.; Li, Q.; Brown, L.; Thatcher, G.R. Nitroxidation, nitration, and oxidation of a BODIPY fluorophore by RNOS and ROS. Nitric Oxide 2006, 15, 163–176. [Google Scholar] [CrossRef]
- Moisenovich, M.M.; Ol’Shevskaya, V.A.; Rokitskaya, T.I.; Ramonova, A.G.; Nikitina, R.G.; Savchenko, A.N.; Tatarskiy, V.V.; Kaplan, M.A.; Kalinin, V.N.; Kotova, E.A.; et al. Novel Photosensitizers Trigger Rapid Death of Malignant Human Cells and Rodent Tumor Transplants via Lipid Photodamage and Membrane Permeabilization. PLoS ONE 2010, 5, e12717. [Google Scholar] [CrossRef]
- Bleeke, T.; Zhang, H.; Madamanchi, N.R.; Patterson, C.; Faber, J.E. Catecholamine-Induced Vascular Wall Growth Is Dependent on Generation of Reactive Oxygen Species. Circ. Res. 2004, 94, 37–45. [Google Scholar] [CrossRef]
- Gauuan, P.J.F.; Trova, M.P.; Gregor-Boros, L.; Bocckino, S.B.; Crapo, J.D.; Day, B.J. Superoxide dismutase mimetics: Synthesis and structure–activity relationship study of MnTBAP analogues. Bioorg. Med. Chem. 2002, 10, 3013–3021. [Google Scholar] [CrossRef]
- Waris, G.; Ahsan, H. Reactive oxygen species: Role in the development of cancer and various chronic conditions. J. Carcinog. 2006, 5, 14. [Google Scholar] [CrossRef]
- Zhang, M.; Harashima, N.; Moritani, T.; Huang, W.; Harada, M. The Roles of ROS and Caspases in TRAIL-Induced Apoptosis and Necroptosis in Human Pancreatic Cancer Cells. PLoS ONE 2015, 10, e127386. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.T.; Korivi, M.; Huang, H.C.; Thiyagarajan, V.; Lin, K.Y.; Huang, P.J.; Liu, J.Y.; Hseu, Y.C.; Yang, H.L. Inhibition of ROS production, autophagy or apoptosis signaling reversed the anticancer properties of Antrodia salmonea in triple-negative breast cancer (MDA-MB-231) cells. Food Chem. Toxicol. 2017, 103, 1–17. [Google Scholar] [CrossRef]
- Gille, J.; Joenje, H. Cell culture models for oxidative stress: Superoxide and hydrogen peroxide versus normobaric hyperoxia. Mutat. Res. 1992, 275, 405–414. [Google Scholar] [CrossRef]
- Kello, M.; Drutovic, D.; Chripkova, M.; Pilátová, M.; Budovská, M.; Kulikova, L.; Urdzik, P.; Mojzis, J. ROS-Dependent Antiproliferative Effect of Brassinin Derivative Homobrassinin in Human Colorectal Cancer Caco2 Cells. Molecules 2014, 19, 10877–10897. [Google Scholar] [CrossRef]
- Cao, X.H.; Wang, A.H.; Wang, C.; Mao, D.Z.; Lu, M.F.; Cui, Y.; Jiao, R.Z. Surfactin induces apoptosis in human breast cancer MCF-7 cells through a ROS/JNK-mediated mitochondrial/caspase pathway. Chem. Biol. Interact. 2010, 183, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Mqoco, T.; Marais, S.; Joubert, A. Influence of estradiol analogue on cell growth, morphology and death in esophageal carcinoma cells. Biocell 2010, 34, 113–120. [Google Scholar]
Sample Availability: Samples of selected compounds (2-ethyl-3-hydroxyestra-1(10),2,4-trien-17-one, 2-vinylestra-1(10),2,4,16-tetraene-3,17-diol, 2-ethylestra-1(10),2,4-triene-3,17-diol) are available from the authors. |















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Sulphamoylated Compound | Sulphamoylated Compound |
|---|---|
 2-Ethyl-3-hydroxyestra-1(10),2,4-trien-17-one (EE-one) also known as C5). |  2-Ethyl-17-oxoestra-1,3,5(10)-trien-3-yl sulphamate (ESE-one) also known as C15. |
 2-Vinylestra-1(10),2,4,16-tetraene-3,17-diol (EE-15-ol) also known as C11. |  2-Ethyl-17-hydroxyestra-1(10),2,4,16-tetreane-3-yl sulfamate (ESE-15-ol) also known as C10. |
 2-Ethylestra-1(10),2,4-triene-3,17-diol (2-E-diol) also known as C13. |  2-Ethyl-17-hydroxyestra-1(10),2,4-trien-3-yl sulfamate (ESE-ol) also known as C16. |
| Reactive Oxygen Species | Scavenger | Concentration |
|---|---|---|
| Hydrogen peroxide | DMTU | 1–10 mM [32] |
| Hydroxyl radical | Mannitol | 20–100 mM [32] |
| Nitric oxide | Carboxy-PTIO | 10–100 µM [32,33] |
| Peroxyl radical | Trolox | 10–100 µM [34,35] |
| Singlet oxygen | Sodium azide | 1–10 mM [32,36]) |
| Superoxide anion | Tiron | 1–10 mM [37,38] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebelo, M.T.; Joubert, A.M.; Visagie, M.H. Sulphamoylated Estradiol Analogue Induces Reactive Oxygen Species Generation to Exert Its Antiproliferative Activity in Breast Cancer Cell Lines. Molecules 2020, 25, 4337. https://doi.org/10.3390/molecules25184337
Lebelo MT, Joubert AM, Visagie MH. Sulphamoylated Estradiol Analogue Induces Reactive Oxygen Species Generation to Exert Its Antiproliferative Activity in Breast Cancer Cell Lines. Molecules. 2020; 25(18):4337. https://doi.org/10.3390/molecules25184337
Chicago/Turabian StyleLebelo, Maphuti T., Anna M. Joubert, and Michelle H. Visagie. 2020. "Sulphamoylated Estradiol Analogue Induces Reactive Oxygen Species Generation to Exert Its Antiproliferative Activity in Breast Cancer Cell Lines" Molecules 25, no. 18: 4337. https://doi.org/10.3390/molecules25184337
APA StyleLebelo, M. T., Joubert, A. M., & Visagie, M. H. (2020). Sulphamoylated Estradiol Analogue Induces Reactive Oxygen Species Generation to Exert Its Antiproliferative Activity in Breast Cancer Cell Lines. Molecules, 25(18), 4337. https://doi.org/10.3390/molecules25184337