
Production and Characterization of Antioxidative Hydrolysates and Peptides from Corn Gluten Meal Using Papain, Ficin, and Bromelain



Abstract
1. Introduction
2. Results and Discussion
2.1. Effect of Hydrolysis Time on Antioxidant Production with Papain, Ficin, and Bromelain
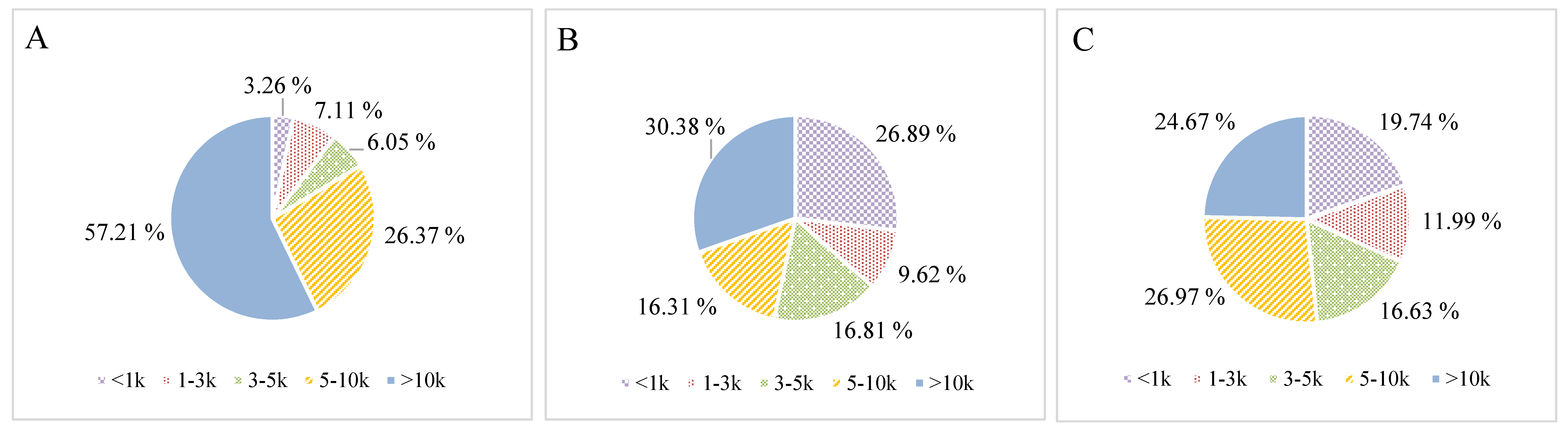
2.2. Antioxidant Properties of Ultrafiltrated Hydrolysate Fractions
2.3. Identification of Peptide Sequences
2.4. Inhibition of Lipid Oxidation in Ground Pork
3. Experimental Section
3.1. Materials
3.2. Preparation of Corn Gluten Meal Hydrolysates
3.3. Determination of Antioxidant Yield
3.4. Determination of Degree of Hydrolysis (DH)
3.5. Fractionation of CGM Hydrolysate by Ultrafiltration
3.6. Determination of Total Phenolic Content (TPC)
3.7. Determination of Antioxidant Activity
3.7.1. Determination of DPPH Radical Scavenging Activity
3.7.2. Determination of ABTS Radical Scavenging Activity
3.7.3. Determination of Ferrous ion (Fe2+) Chelating Activity
3.8. Identification of Peptide Sequences of Selected Antioxidant Peptide
3.9. Antioxidant Activity of Selected Hydrolysates in Ground Pork
3.10. Statistic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ortiz-Martinez, M.; Otero-Pappatheodorou, J.T.; Serna-Saldívar, S.O.; García-Lara, S. Antioxidant and characterization of protein fractions and hydrolysates from normal and quality protein maize kernels. J. Cereal Sci. 2017, 76, 85–91. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Liu, Q.; Kong, B.; Diao, X. Effects of zein hydrolysates coupled with sage (salvia officinalis) extract on the emulsifying and oxidative stability of myofibrillar protein prepared oil-in-water emulsions. Food Hydrocoll. 2019, 87, 149–157. [Google Scholar] [CrossRef]
- Wang, Y.; Yuan, Y.; Yang, X.; Wang, J.; Guo, J.; Lin, Y. Comparison of the colloidal stability, bioaccessibility and antioxidant activity of corn protein hydrolysate and sodium caseinate stabilized curcumin nanoparticles. Food Sci. Technol. 2016, 53, 2923–2932. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.M.; Flôres, S.H.; Heck, J.X.; Ayub, M.A.Z. Production of high-protein hydrolysate from poultry industry residue and their molecular profiles. Food Biotechnol. 2009, 23, 229–242. [Google Scholar]
- Bougatef, A.; Hajji, M.; Balti, R.; Lassoued, I.; Triki-Ellouz, Y.; Nasri, M. Antioxidant and free radical-scavenging activities of smooth hound (Mustelus mustelus) muscle protein hydrolysates obtained by gastrointestinal proteases. Food Chem. 2009, 114, 1198–1205. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T. Isolation and characterization of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Kim, J.M.; Whang, J.H.; Kim, K.M.; Koh, J.H.; Suh, H.J. Preparation of corn gluten hydrolysates with angiotensin I converting enzyme inhibitory activity and its solubility and moisture sorption. Process Biochem. 2004, 39, 989–994. [Google Scholar] [CrossRef]
- Samaraweera Mudiyanselage, C.H. Production and Characterization of Phosphopeptides from Egg Yolk Phosvitin. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2012. [Google Scholar]
- Zhu, B.; He, H.; Hou, T. A comprehensive review of corn protein-derived bioactive peptides: Production, characterization, bioactivities, and transport pathways: Review of corn proteins-derived bioactive peptides. Compr. Rev. Food Sci. Food Saf. 2019, 18, 329–345. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, X.; Kopparapu, N.; Cong, W.; Deng, Y.; Sun, X.; Liu, X. Purification and evaluation of a novel antioxidant peptide from corn protein hydrolysate. Process Biochem. 2014, 49, 1562–1569. [Google Scholar] [CrossRef]
- Huang, W.; Sun, J.; He, H.; Dong, H.; Li, J. Antihypertensive effect of corn protein hydrolysates, produced by a continuous production in enzymatic membrane reactor, in spontaneously hypertensive rats. Food Chem. 2011, 128, 968–973. [Google Scholar] [CrossRef]
- Li, H.; Guo, P.; Hu, X.; Fang, P.; Li, X.; Zhang, X. Antioxidant properties and possible mode of action of corn protein peptides and zein peptides. Food Biochem. 2010, 34, 44–60. [Google Scholar] [CrossRef]
- Sun, P.; Tian, X.; Shi, R. Effects of dietary corn peptide and exercise on hepatic ATGL and TNF-α in obese rats. Chin. J. Physiol. 2017, 33, 117–120. [Google Scholar]
- Jin, D.; Liu, X.; Zheng, X.; Wang, X.; He, J. Preparation of antioxidative corn protein hydrolysates, purification and evaluation of three novel corn antioxidant peptides. Food Chem. 2016, 204, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zheng, X.; Song, Z.; Liu, X.; Kopparapu, N.K.; Wang, X.; Zheng, Y. Preparation of enzymatic pretreated corn gluten meal hydrolysate and in vivo evaluation of its antioxidant activity. Funct. Foods 2015, 18, 1147–1157. [Google Scholar] [CrossRef]
- Gu, R.; Li, C.; Liu, W.; Yi, W.; Cai, M. Angiotensin I-converting enzyme inhibitory activity of low-molecular-weight peptides from Atlantic salmon (Salmo salar L.) skin. Food Res. Int. 2011, 44, 1536–1540. [Google Scholar] [CrossRef]
- Qin, L.; Zhu, B.; Zhou, D.; Wu, H.; Tan, H.; Yang, J.; Li, D.; Dong, X.; Murata, Y. Preparation and antioxidant activity of enzymatic hydrolysis from purple sea urchin (Stronglyocentrotus nudus) gonad. Lwt-Food Sci. Technol. 2011, 44, 1113–1118. [Google Scholar] [CrossRef]
- Zhuang, L.; Tang, N.; Dong, S.; Sun, B.; Liu, J. Optimization of antioxidant peptide preparation from corn gluten meal. Sci. Food Agric. 2013, 93, 3264–3270. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilpour, M.; Ehsani, M.R.; Aminlari, M.; Shekarforoush, S.; Hoseini, E. Antimicrobial activity of peptides derived from enzymatic hydrolysis of goat milk caseins. Comp. Clin. Pathol. 2016, 25, 599–605. [Google Scholar] [CrossRef]
- Feijoo-Siota, L.; Villa, T.G.B.V.J.; Barros Velazquez, J. Native and biotechnologically engineered plant proteases with industrial applications. Food Bioprocess Technol. 2011, 4, 1066–1088. [Google Scholar] [CrossRef]
- Holyavka, M.; Pankova, S.; Koroleva, V.; Vyshkvorkina, Y.; Lukin, A.; Kondratyev, M.; Artyukhov, V. Influence of UV radiation on molecular structure and catalytic activity of free and immobilized bromelain, ficin and papain. Photochem. Photobiol. 2019, 201, 111681. [Google Scholar] [CrossRef]
- Wharton, W.C. The structure and mechanism of stem bromelain evaluation of the homogeneity of purified stem bromelain, determination of the molecular weight and kinetic analysis of the bromelain-catalysed hydrolysis of Nbenzyloxycarbonyl-L-phenylalanyl-L-serine methyl ester. Biochemical 1974, 143, 575–586. [Google Scholar]
- Selamassakul, O.; Laohakunjit, N.; Kerdchoechuen, O.; Yang, L.; Maier, C.S. Bioactive peptides from brown rice protein hydrolyzed by bromelain: Relationship between biofunctional activities and flavor characteristics. Food Sci. 2020, 85, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cheng, X.; Wang, X.; Wu, S. Free-radical-scavenging and anti-inflammatory effect of yak milk casein before and after enzymatic hydrolysis. Food Chem. 2011, 126, 484–490. [Google Scholar] [CrossRef]
- Zhu, K.; Zhou, H.; Qian, H. Antioxidant and free radical-scavenging activities of wheat germ protein hydrolysates (WGPH) prepared with alcalase. Process Biochem. 2006, 41, 1296–1302. [Google Scholar] [CrossRef]
- Zhou, K.; Canning, C.; Sun, S. Effects of rice protein hydrolysates prepared by microbial proteases and ultrafiltration on free radicals and meat lipid oxidation. Food Sci. Technol. 2013, 50, 331–335. [Google Scholar] [CrossRef]
- Hidalgo, M.E.; Folmer Côrrea, A.P.; Mancilla Canales, M.; Joner Daroit, D.; Brandelli, A.; Risso, P. Biological and physicochemical properties of bovine sodium caseinate hydrolysates obtained by a bacterial protease preparation. Food Hydrocoll. 2015, 43, 510–520. [Google Scholar] [CrossRef]
- Oliveira, C.F.; Coletto, D.; Correa, A.P.; Daroit, D.J.; Toniolo, R.; Cladera-Olivera, F.; Brandelli, A. Antioxidant activity and inhibition of meat lipid oxidation by soy protein hydrolysates obtained with a microbial protease. Int. Food Res. 2014, 21, 775–781. [Google Scholar]
- Chi, C.; Wang, B.; Hu, F.; Wang, Y.; Zhang, B.; Deng, S.; Wu, C. Purification and identification of three novel antioxidant peptides from protein hydrolysate of bluefin leatherjacket (Navodon septentrionalis) skin. Food Res. Int. 2015, 73, 124–129. [Google Scholar] [CrossRef]
- Liu, F.; Chen, Z.; Shao, J.; Wang, C.; Zhang, C. Effect of fermentation on the peptide content, phenolics and antioxidant activity of defatted wheat germ. Food Biosci. 2017, 20, 141–148. [Google Scholar] [CrossRef]
- Zhuang, H.; Tang, N.; Yuan, Y. Purification and identification of antioxidant peptides from corn gluten meal. Funct. Foods 2013, 5, 1810–1821. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Kou, X.; Gao, J.; Xue, X.; Zhang, Z.; Wang, H.; Wang, X. Purification and identification of antioxidant peptides from chickpea (Cicer arietinum L.) albumin hydrolysates. Lmt-Food Sci. Technol. 2013, 50, 591–598. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, J.; Tang, X.; Xiong, L. Reducing, radical scavenging, and chelation properties of in vitro digests of alcalase-treated zein hydrolysate. Agric. Food Chem. 2008, 56, 2714–2721. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.R.; Burney, J.D.; Black, K.W.; Zaloga, G.P. Effect of chain length on absorption of biologically active peptides from the gastrointestinal tract. Digestion 1999, 60, 332–337. [Google Scholar] [CrossRef]
- Zhou, C.; Hu, J.; Ma, H.; Yagoub, A.E.A.; Yu, X.; Owusu, J.; Ma, H.; Qin, X. Antioxidant peptides from corn gluten meal: Orthogonal design evaluation. Food Chem. 2015, 187, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Shen, Y.; Li, Y. Antioxidant activities of sorghum kafirin Alcalase hydrolysates and membrane/gel filtrated fractions. Antioxidants 2019, 8, 131. [Google Scholar] [CrossRef]
- Agrawal, H.; Joshi, R.; Gupta, M. Isolation and characterization of enzymatic hydrolyzed peptides with antioxidant activities from green tender sorghum. Lwt-Food Sci. Technol. 2017, 84, 608–616. [Google Scholar] [CrossRef]
- Hogan, S.; Zhang, L.; Li, J.; Wang, H.; Zhou, K. Development of antioxidant rich peptides from milk protein microbial proteases and analysis of their effects on lipid peroxidation in cooked beef. Food Chem. 2009, 117, 438–443. [Google Scholar] [CrossRef]
- Sonklin, C.; Laohakunjit, N.; Kerdcheochuen, O. Assessment of antioxidant properties of membrane ultrafiltration peptides from mungbean protein hydrolysates. Natl. Inst. Health 2018, 6, e5337. [Google Scholar] [CrossRef]
- Li, H.; Guo, P.; Hu, X.; Xu, L.; Zhang, X. Preparation of corn (Zea mays) peptides and their protective effect against alcohol-induced acute hepatic injury in NH mice. Biotechnol. Appl. Biochem. 2007, 47, 169–174. [Google Scholar]
- Hu, R.; Dunmire, K.M.; Truelock, C.N.; Paulk, C.B.; Aldrich, G.; Li, Y. Antioxidant performances of corn gluten meal and DDGS protein hydrolysates in food, pet food, and feed systems. J. Agric. Food Res. 2020, 2, 100030. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Jung, W.; Je, J.; Kim, S.K. Purification of a radical scavenging peptide from fermented mussel sauce ant its antioxidant properties. Food Res. Int. 2005, 38, 175–182. [Google Scholar] [CrossRef]
- Harman, L.S.; Mottley, C.; Mason, R.P. Free radical metabolites of L-cysteine oxidation. Biol. Chem. 1984, 259, 5606–5611. [Google Scholar]
- Zhou, K.; Sun, S.; Canning, C. Production and functional characterization of antioxidative hydrolysates from corn protein via enzymatic hydrolysis and ultrafiltration. Food Chem. 2012, 135, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Shen, Y.; Xu, J.; Qi, G.; Chen, G.; Wang, W.; Sun, X.; Li, Y. Antioxidant and anticancer effects in human hepatocarcinoma (HepG2) cells of papain-hydrolyzed sorghum kafirin hydrolysates. Funct. Foods 2019, 58, 375–382. [Google Scholar] [CrossRef]
- Moure, A.; Domínguez, H.; Parajó, J.C. Antioxidant properties of ultrafiltration-recovered soy protein fractions from industrial effluents and their hydrolysates. Process. Biochem. 1991, 41, 447–456. [Google Scholar] [CrossRef]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Hayes, K.D. Comparative study on antioxidant activity of yellow strip trevally protein hydrolysate produced from alcalase and flavourzyme. Int. J. Food Sci. Technol. 2008, 43, 1019–1026. [Google Scholar] [CrossRef]
- Jadhav, S.J.; Nimbalkar, S.S.; Kulkarni, A.D.; Madhavi, D.L. Lipid oxidation in biological and food systems. In Food Antioxidants; Taylor & Francis Group: Abingdon, UK, 1996; pp. 5–63. [Google Scholar]
- Cheetangdee, N.; Benjakul, S. Antioxidant activities of rice bran protein hydrolysates in bulk oil and oil-in-water emulsion. Sci. Agric. 2015, 95, 1461–1468. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Thamnarathip, P.; Jangchud, K.; Nitisinprasert, S.; Vardhanabhuti, B. Identification of peptide molecular weight from rice bran protein hydrolysate with high antioxidant activity. Cereal Sci. 2016, 69, 329–335. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Hawkins Byrne, D. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Zhang, L.; Li, J.; Zhou, K. Chelating and radical scavenging activities of soy protein hydrolysates prepared from microbial proteases and their effect on meat lipid peroxidation. Bioresour. Technol. 2010, 101, 2084–2089. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Type | Enzyme Amount, mg/g of Protein | pH | Temperature, °C | Time, h |
|---|---|---|---|---|
| Papain | 40 | 6.5 | 50 | 0.5–5 |
| Ficin | 225.5 | 6.0 | 50 | 0.5–5 |
| Bromelain | 150 | 5.0 | 50 | 0.5–5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, R.; Chen, G.; Li, Y. Production and Characterization of Antioxidative Hydrolysates and Peptides from Corn Gluten Meal Using Papain, Ficin, and Bromelain. Molecules 2020, 25, 4091. https://doi.org/10.3390/molecules25184091
Hu R, Chen G, Li Y. Production and Characterization of Antioxidative Hydrolysates and Peptides from Corn Gluten Meal Using Papain, Ficin, and Bromelain. Molecules. 2020; 25(18):4091. https://doi.org/10.3390/molecules25184091
Chicago/Turabian StyleHu, Ruijia, Gengjun Chen, and Yonghui Li. 2020. "Production and Characterization of Antioxidative Hydrolysates and Peptides from Corn Gluten Meal Using Papain, Ficin, and Bromelain" Molecules 25, no. 18: 4091. https://doi.org/10.3390/molecules25184091
APA StyleHu, R., Chen, G., & Li, Y. (2020). Production and Characterization of Antioxidative Hydrolysates and Peptides from Corn Gluten Meal Using Papain, Ficin, and Bromelain. Molecules, 25(18), 4091. https://doi.org/10.3390/molecules25184091