
Comprehensive Metabolome Analysis of Fermented Aqueous Extracts of Viscum album L. by Liquid Chromatography−High Resolution Tandem Mass Spectrometry

 ,
,
Abstract
1. Introduction
2. Results and Discussion
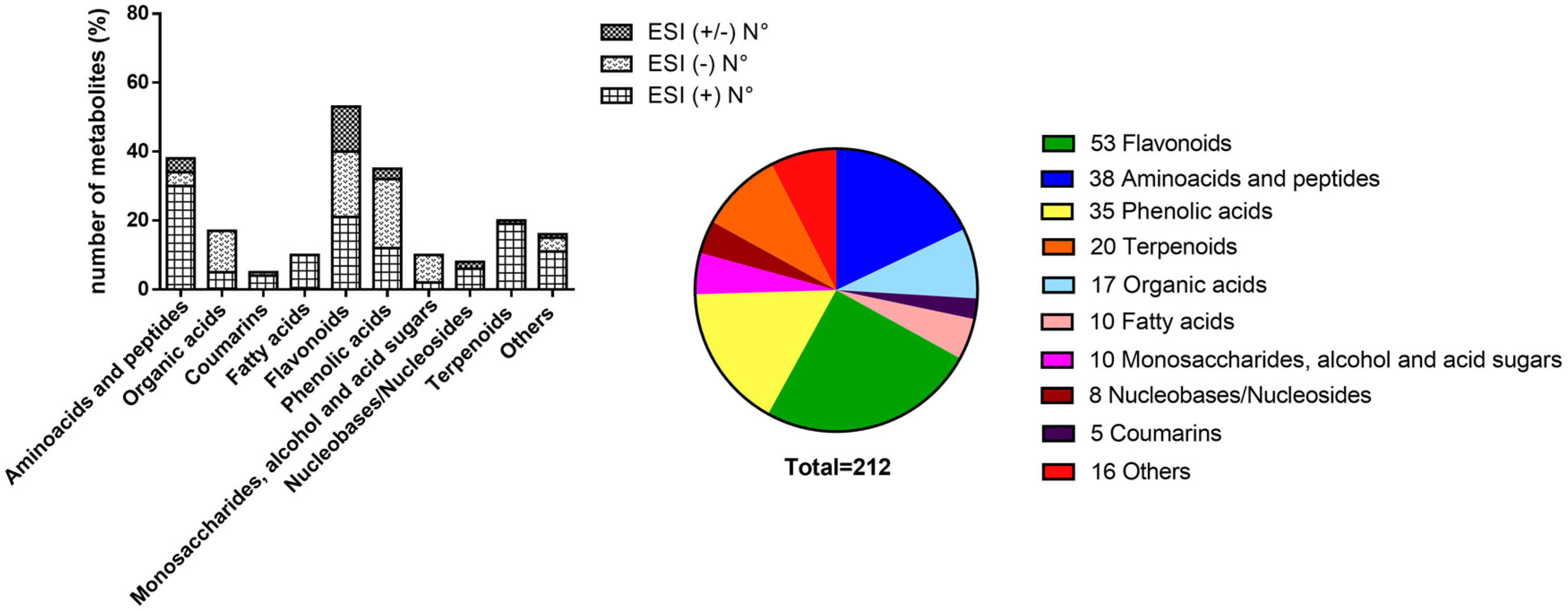
2.1. LC-HRMS/MS Analysis and Compound Annotation
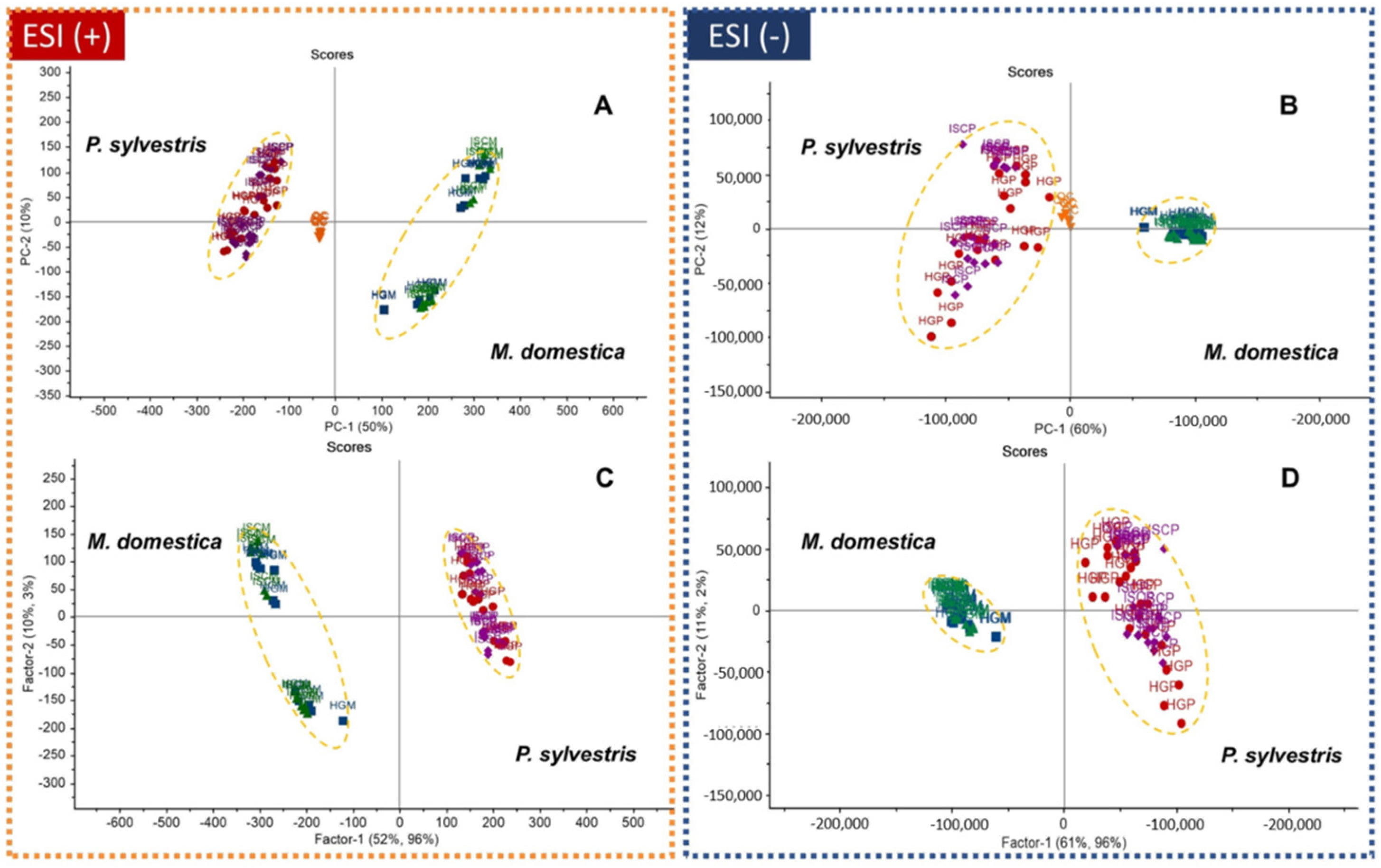
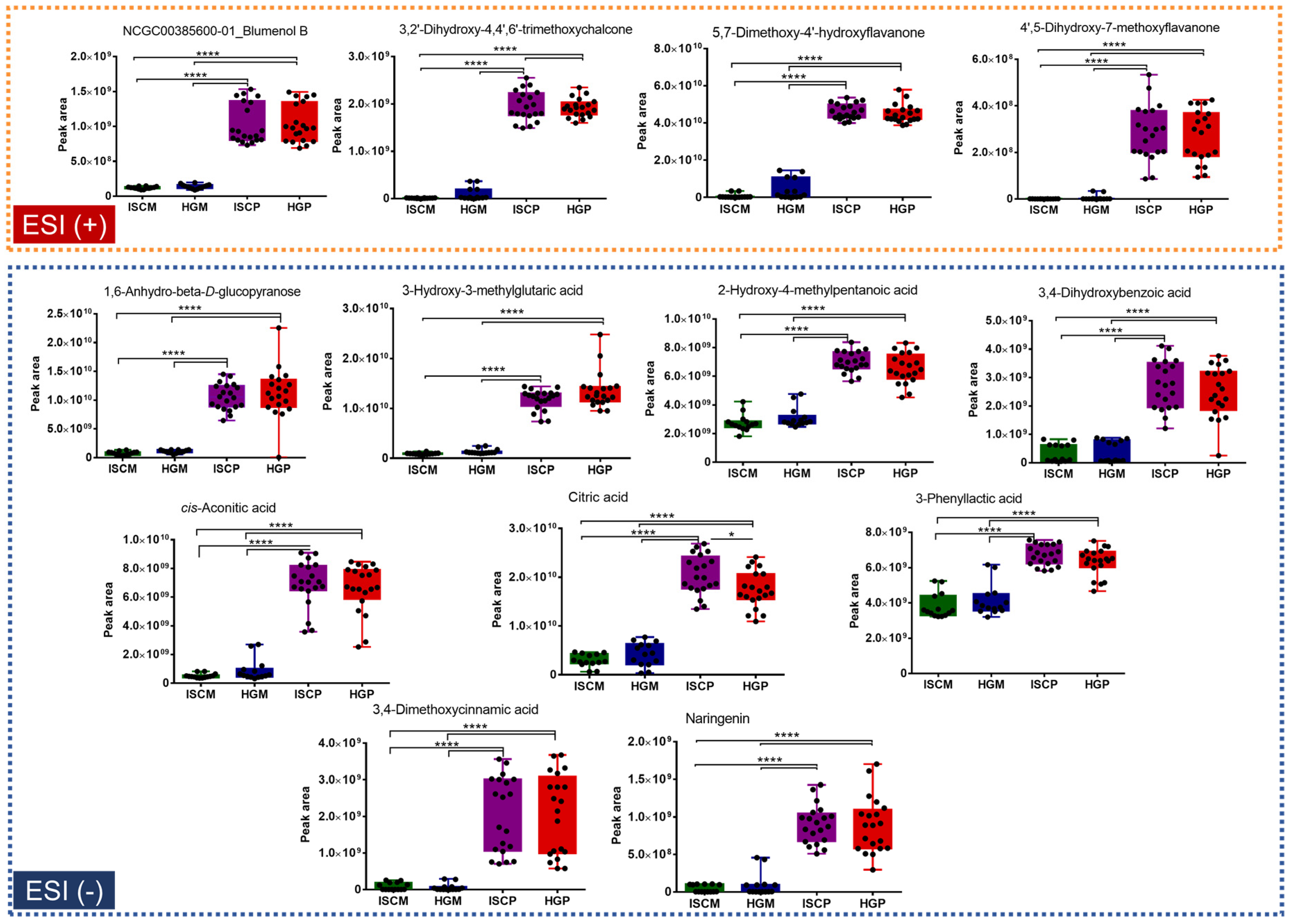
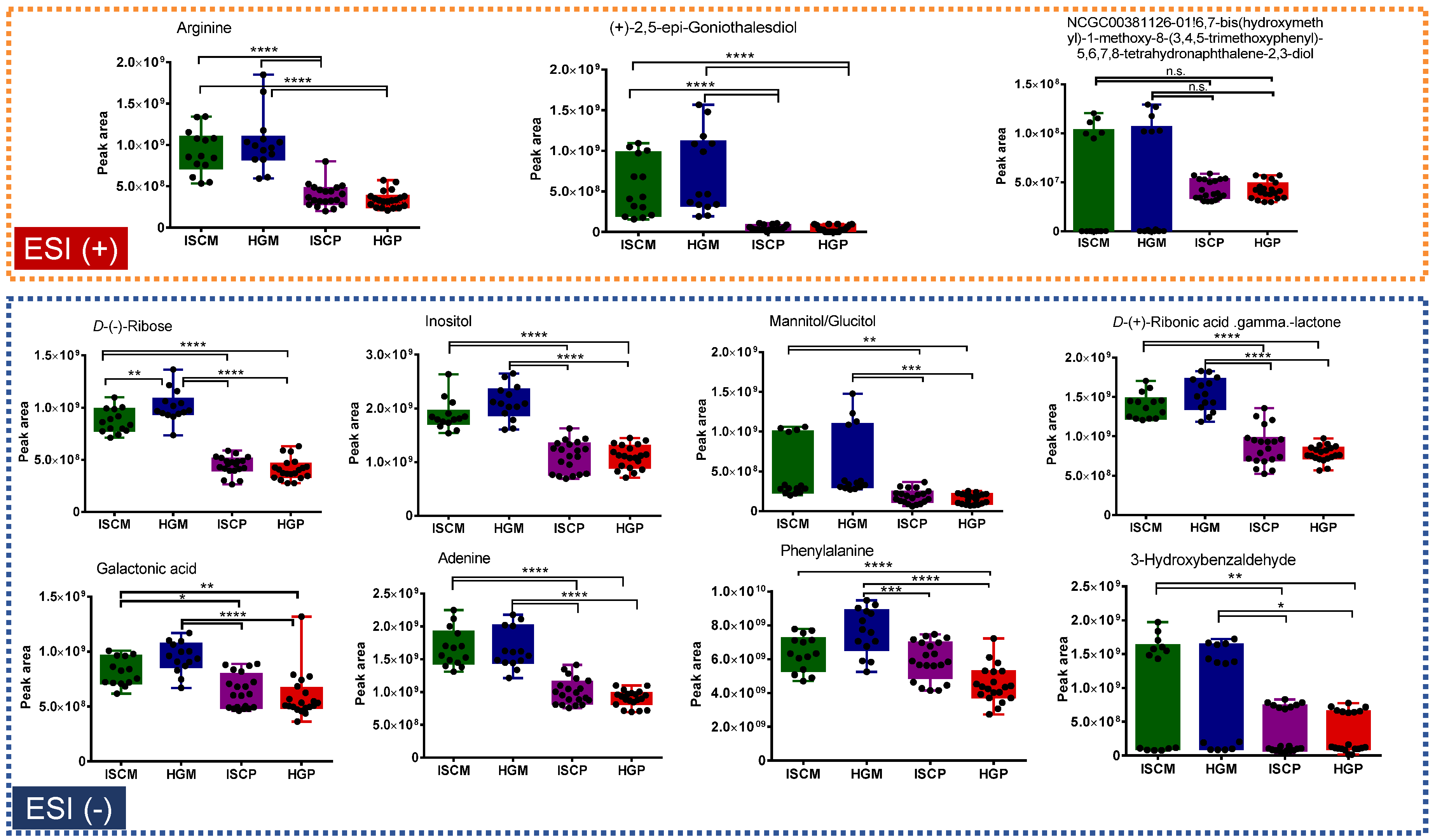
2.2. Multivariate Data Analysis
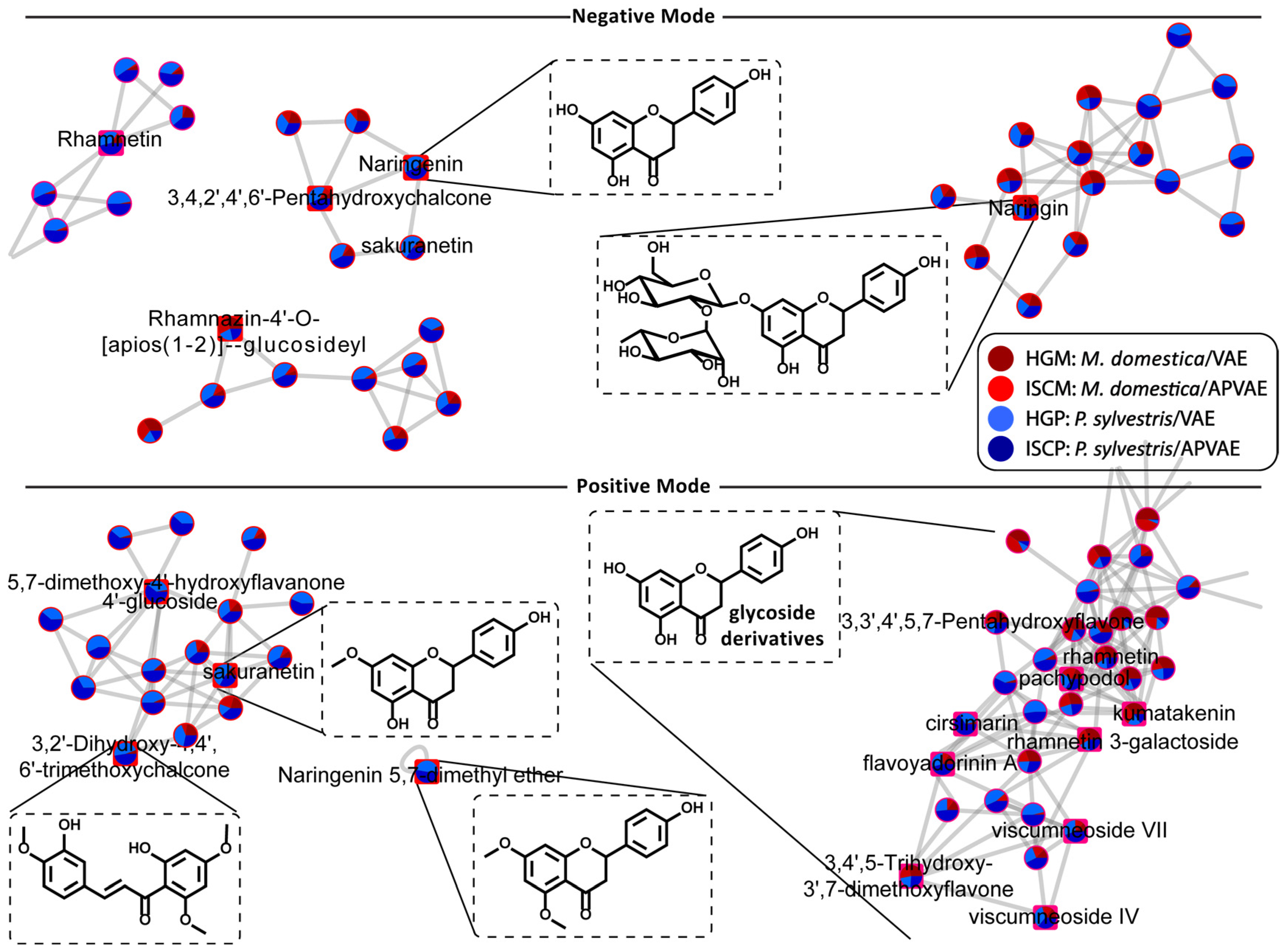
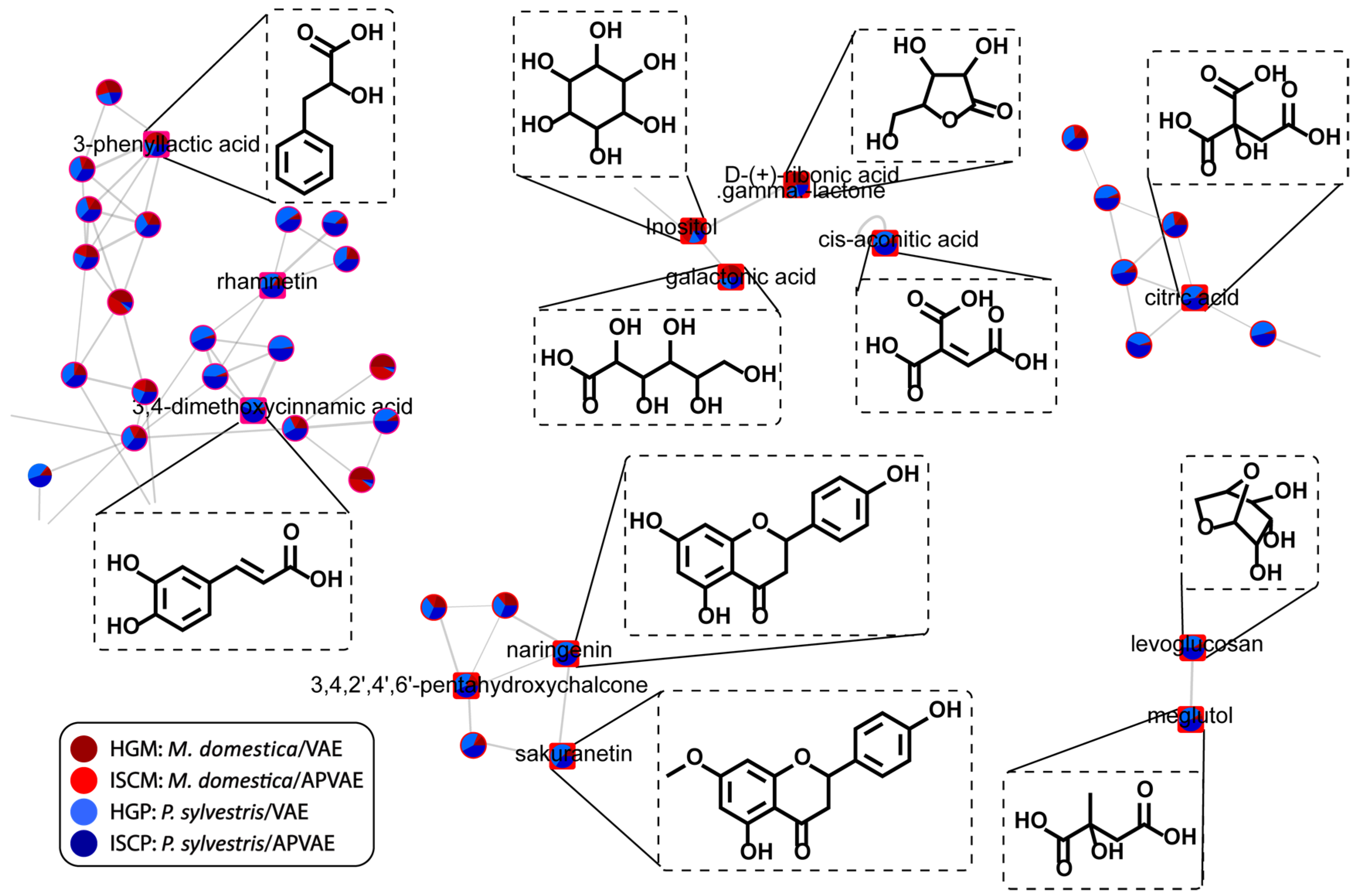
2.3. Chemical Space by Molecular Networking
3. Materials and Methods
3.1. Chemicals and Materials
3.2. Sample Preparation
3.3. LC-HRMS/MS Analysis
3.4. Data Processing and Metabolite Identification
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vergara-Barberán, M.; Lerma-García, M.J.; Nicoletti, M.; Simó-Alfonso, E.F.; Herrero-Martínez, J.M.; Fasoli, E.; Righetti, P.G. Proteomic fingerprinting of mistletoe (Viscum album L.) via combinatorial peptide ligand libraries and mass spectrometry analysis. J. Proteom. 2017, 164, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Ramm, H.; Urech, K.; Scheibler, M.; Grazi, G. Cultivation and Development of Viscum album L. In Mistletoe: The genus Viscum; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 75–94. ISBN 978-90-5823-092-9. [Google Scholar]
- Zuber, D. Biological flora of Central Europe: Viscum album L. Flora-Morphol. Distrib. Funct. Ecol. Plants 2004, 199, 181–203. [Google Scholar] [CrossRef]
- Becker, H. European mistletoe: Taxonomy, host trees, parts used, physiology. In Mistletoe: The Genus Viscum; Harwood Academic Publishers: Amsterdam, The Netherlands, 2000; pp. 31–43. ISBN 90-5823-092-9. [Google Scholar]
- Nazaruk, J.; Orlikowski, P. Phytochemical profile and therapeutic potential of Viscum album L. Nat. Prod. Res. 2016, 30, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Saha, C.; Galun, D.; Upreti, D.K.; Bayry, J.; Kaveri, S.V. European Viscum album: A potent phytotherapeutic agent with multifarious phytochemicals, pharmacological properties and clinical evidence. R. Soc. Chem. 2016, 6, 23837–23857. [Google Scholar] [CrossRef]
- Bar-Sela, G. White-Berry Mistletoe (Viscum album L.) as complementary treatment in cancer: Does it help? Eur. J. Integr. Med. 2011, 3, e55–e62. [Google Scholar] [CrossRef]
- Urech, K.; Baumgartner, S. Chemical Constituents of Viscum album L.: Implications for the Pharmaceutical Preparation of Mistletoe. In Mistletoe-From Mythology to Evidence-Based Medicine; Karger: Basel, Switzerland, 2015; Volume 4, pp. 11–23. ISBN 978-3-318-05444-6. [Google Scholar]
- Eggenschwiler, J.; von Balthazar, L.; Stritt, B.; Pruntsch, D.; Ramos, M.; Urech, K.; Rist, L.; Simões-Wüst, A.P.; Viviani, A. Mistletoe lectin is not the only cytotoxic component in fermented preparations of Viscum album from white fir (Abies pectinata). BMC Complementary Altern. Med. 2007, 7, 2–7. [Google Scholar] [CrossRef]
- Bonamin, L.V.; Cunha de Carvalho, A.; Waisse, S. Viscum album L. in experimental animal tumors: A meta-analysis. Experimental and Therapeutic medicine. Exp. Ther. Med. 2017, 13, 2723–2740. [Google Scholar] [CrossRef]
- Loef, M.; Walach, H. Quality of life in cancer patients treated with mistletoe: A systematic review and meta-analysis. BMC Complementary Med. Ther. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Ostermann, T.; Appelbaum, S.; Poier, D.; Boehm, K.; Raak, C.; Büssing, A. A Systematic Review and Meta-Analysis on the Survival of Cancer Patients Treated with a Fermented Viscum album L. Extract (Iscador): An Update of Findings. Complementary Med. Res. 2020, 1–12. [Google Scholar] [CrossRef]
- Melzer, J.; Iten, F.; Hostanska, K.; Saller, R. Efficacy and Safety of Mistletoe Preparations (Viscum album) for Patients with Cancer Diseases. Forsch. Komplementarmed. 2009, 16, 217–226. [Google Scholar] [CrossRef]
- Matthes, H.; Thronicke, A.; Hofheinz, R.-D.; Baars, E.; Martin, D.; Huber, R.; Breitkreuz, T.; Bar-Sela, G.; Galun, D.; Schad, F. Statement to an Insufficient Systematic Review on Viscum album L. Therapy. Evid. Based Complement. Altern. Med. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bussing, A. Mistletoe: The Genus Viscum; Medicinal and Aromatic Plants-Industrial Profiles; Harwood Academic: Amsterdam, The Netherlands, 2000; ISBN 978-90-5823-092-9. [Google Scholar]
- Urech, K.; Schaller, G.; Jäggy, C. Viscotoxins, mistletoe lectins and their isoforms in mistletoe (Viscum album L.) extracts Iscador. Arzneim. Drug Res. 2006, 56, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Szurpnicka, A.; Zjawiony, J.K.; Szterk, A. Therapeutic potential of mistletoe in CNS-related neurological disorders and the chemical composition of Viscum species. J. Ethnopharmacol. 2019, 231, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Vicas, S.I.; Rugina, D.; Leopold, L.; Pintea, A.; Socaciu, C. HPLC Fingerprint of Bioactive Compounds and Antioxidant Activities of Viscum album from Different Host Trees. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 48–57. [Google Scholar] [CrossRef]
- Panossian, A.; Kocharian, A.; Matinian, K.; Amroyan, E.; Gabrielian, E.; Mayr, C.; Wagner, H. Pharmacological activity of phenylpropanoids of the mistletoe, Viscum album L., host: Pyrus caucasica Fed. Phytomedicine 1998, 5, 11–17. [Google Scholar] [CrossRef]
- Baumgartner, S.; Flückiger, H.; Kunz, M.; Scherr, C.; Urech, K. Evaluation of Preclinical Assays to Investigate an Anthroposophic Pharmaceutical Process Applied to Mistletoe (Viscum album L.) Extracts. Evid. Based Complement. Altern. Med. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Salem, M.A.; Souza, L.P.; Serag, A.; Fernie, A.R.; Farag, M.A.; Ezzat, S.M.; Alseekh, S. Metabolomics in the Context of Plant Natural Products Research: From Sample Preparation to Metabolite Analysis. Metabolites 2020, 10, 37. [Google Scholar] [CrossRef]
- Fiehn, O.; Robertson, D.; Griffin, J.; Werf, M.; Nikolau, B.; Morrison, N.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; Taylor, C.; et al. The metabolomics standards initiative (MSI). Metabolomics 2007, 3, 175–178. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. GigaScience 2013, 2, 1–3. [Google Scholar] [CrossRef]
- Park, B.-J.; Matsuta, T.; Samejima, H.; Park, C.-H.; Sung, I.J.; Lee, B.-D.; Onjo, M. Chemical Constituents of Mistletoe (Viscum album L. var. coloratum Ohwi). IOSR J. Pharm. Biol. Sci. 2017, 12, 19–23. [Google Scholar] [CrossRef]
- Richter, A.; Popp, M. The Physiological Importance of Accumulation of Cyclitols in Viscum album L. New Phytol. 1992, 121, 431–438. [Google Scholar] [CrossRef]
- Richter, A. Viscumitol, a dimethyl-ether of muco-inositol from Viscum album. Phytochem. Lett. 1992, 31, 3925–3927. [Google Scholar] [CrossRef]
- Arda, N.; Koz, Ö.; Onay-Uçar, E.; Kırmızıgül, S. Monosaccharides and polyols from mistletoes (Viscum album L.) growing on two different host species. Biol. Bratisl. 2003, 58, 1037–1041. [Google Scholar]
- Kohl, M.; Bäcker, C.; Wurster, M.; Lindequist, U.; Meyer, U.; Stintzing, F. Comparative investigation of monossacarides and sugar alcohols in mistletoes (Viscum album L.) from different host trees. Phytomedicine 2011, 18S, S20–S21. [Google Scholar] [CrossRef]
- Wurster, M.; Albrecht, S.; Lindequist, U.; Stintzing, F. Comparative investigation of nucleosides in mistletoes (Viscum album L.) from different host trees. Phytomedicine 2015, 22, S23–S26. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Martorell, M.; Valdes, S.E.; Belwal, T.; Tejada, S.; Kamal, M.A. Flavonoids nanoparticles in cancer: Treatment, prevention and clinical prospects. Semin. Cancer Biol. 2019, in press. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Fan, R.; Zhang, Q.; Zhang, Z.; Wang, D.; Xia, Y.; Ma, Y.; Yu, Z.; Zhao, Y. Simultaneous identification and quantification of the common compounds of Viscum coloratum and its corresponding host plants by ultra-high performance liquid chromatography with quadrupole time-of-flight tandem mass spectrometry and triple quadrupole mass spectrometry. J. Chromatogr. B 2017, 1061–1062, 176–184. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Farag, M.A.; Abdel-Naim, A.B.; Ghareib, S.A.; Abdel-Sattar, E.A. Mechanistic Evidence of Viscum schimperi (Viscaceae) Antihyperglycemic Activity: From a Bioactivity-guided Approach to Comprehensive Metabolite Profiling. Phytother. Res. 2015, 29, 1737–1743. [Google Scholar] [CrossRef]
- Patel, B.P.; Singh, B.N. Viscum articulatum Burm. f.: A review on its phytochemistry, pharmacology and traditional uses. J. Pharm. Pharmacol. 2018, 70, 159–177. [Google Scholar] [CrossRef]
- Cebovic, T.; Spasic, S.; Popovic, M. Cytotoxic Effects of the Viscum album L. Extract on Ehrlich Tumour Cells In Vivo. Phytother. Res. 2008, 22, 1097–1103. [Google Scholar] [CrossRef]
- Kurkcuoglu, M.; Demirci, B.; Baser, K.H.C.; Deliorman, D. Composition of the essential oils of Viscum album L. Subspecies from Turkey. J. Fac Pharm Gazi Univ. 2002, 19, 87–92. [Google Scholar]
- Park, S.H.; Kim, D.S.; Kim, S.; Lorz, L.R.; Choi, E.; Lim, H.Y.; Hossain, M.A.; Jang, S.; Choi, Y.I.; Park, K.J.; et al. Loliolide Presents Antiapoptosis and Antiscratching Effects in Human Keratinocytes. Int. J. Mol. Sci. 2019, 20, 651. [Google Scholar] [CrossRef] [PubMed]
- Aïssaoui, H.; Mencherini, T.; Esposito, T.; De Tommasi, N.; Gazzerro, P.; Benayache, S.; Benayache, F.; Mekkiou, R. Heliotropium bacciferum Forssk. (Boraginaceae) extracts: Chemical constituents, antioxidant activity and cytotoxic effect in human cancer cell lines. Nat. Prod. Res. 2019, 33, 1813–1818. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Rasul, A.; Sadiqa, A.; Sarfraz, I.; Hussain, G.; Nageen, B.; Liu, X.; Watanabe, N.; Selamoglu, Z.; Ali, M.; et al. Curcumol: From Plant Roots to Cancer Roots. Int. J. Biol. Sci. 2019, 15, 1600–1609. [Google Scholar] [CrossRef]
- Ren, Y.; Shen, L.; Zhang, D.-W.; Dai, S.-J. Two New Sesquiterpenoids from Solanum lyratum with Cytotoxic Activities. Chem. Pharm. Bull. 2009, 57, 408–410. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, K.; Prakash, M.D.; Kuol, N.; Stojanovska, L.; Apostolopoulos, V. Anti-Tumor Effects of Vitamin B2, B6 and B9 in Promonocytic Lymphoma Cells. Int. J. Mol. Sci. 2019, 20, 3763. [Google Scholar] [CrossRef] [PubMed]
- Abraham, I.; Joshi, R.; Pardasani, P.; Pardasani, R.T. Recent advances in 1,4-benzoquinone chemistry. J. Braz. Chem. Soc. 2011, 22. [Google Scholar] [CrossRef]
- Goss, P.E.; Baptiste, J.; Fernandes, B.; Baker, M.; Dennis, J.W. A Phase I Study of Swainsonine in Patients with Advanced Malignancies. Cancer Res. 1994, 54, 1450–1457. [Google Scholar] [CrossRef]
- Becker, H.; Exner, J. Comparative Studies of Flavonoids and Phenylcarboxylic Acids of Mistletoes from Different Host Trees. Z. Für Pflanzenphysiol. 1980, 97, 417–428. [Google Scholar] [CrossRef]
- Łuczkiewicz, M.; Cisowski, W.; Kaiser, P.; Ochocka, R.; Piotrowski, A. Comparative analysis of phenolic acids in mistletoe plants from various hosts. Acta Pol. Pharm. 2001, 58, 373–379. [Google Scholar]
- Pietrzak, W.; Nowak, R.; Gawlik-Dziki, U.; Lemieszek, M.; Rzeski, W. LC-ESI-MS/MS Identification of Biologically Active Phenolic Compounds in Mistletoe Berry Extracts from Different Host Trees. Molecules 2017, 22, 624. [Google Scholar] [CrossRef] [PubMed]
- Jäger, T.; Holandino, C.; Glauser, G.; Grazi, M.; Ramm, H.; Nonato de Oliveira Melo, M.; Passos Oliveira, A.; Garrett, R.; Baumgartner, S. Metabolic profiling as a tool for differentiating Viscum album ssp. album plants growing on various host trees. Phytomedicine 2019, 61, 153079. [Google Scholar] [CrossRef]
- Gärtner, T.; Link, K.; Müller, M.B.; Stintzing, F.C.; Kammerer, D.R. Phenolic profiles of Viscum album L. subspecies from different host trees. Phytomedicine 2015, 22, S23–S26. [Google Scholar] [CrossRef]
- Deliorman, D.; Çali˝, I.; Ergun, F.; Tamer, U. The comparative studies on phenylpropanoid glycosides of Viscum album subspecies by high performance liquid chromatography. J. Liq. Chromatogr. Relat. Technol. 1999, 22, 3101–3114. [Google Scholar] [CrossRef]
- Nothias, L.F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based Molecular Networking in the GNPS Analysis Environment. BioRxiv Prepr. Serve Biol. 2019, 1–19. [Google Scholar] [CrossRef]
- Resende, J.V.M.; de Sá, N.M.D.; de Oliveira, M.T.L.; Lopes, R.C.; Garrett, R.; Borges, R.M. Chemical profiling of herbarium samples of solanum (Solanaceae) using mass spectrometry. Phytochem. Lett. 2020, 99, 99–105. [Google Scholar] [CrossRef]
- Ernst, M.; Kang, K.B.; Caraballo-Rodríguez, A.M.; Nothias, L.-F.; Wandy, J.; Chen, C.; Wang, M.; Rogers, S.; Medema, M.H.; Dorrestein, P.C.; et al. MolNetEnhancer: Enhanced Molecular Networks by Integrating Metabolome Mining and Annotation Tools. Metabolites 2019, 9, 144. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-diaL: Data-independent ms/ms deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–531. [Google Scholar] [CrossRef]
- Shinbo, Y.; Nakamura, Y.; Altaf-Ul-Amin, M.; Asahi, H.; Kurokawa, K.; Arita, M.; Saito, K.; Ohta, D.; Shibata, D.; Kanaya, S. KNApSAcK: A Comprehensive Species-Metabolite Relationship Database. Available online: http://www.knapsackfamily.com/KNApSAcK/ (accessed on 18 October 2019).
- Taylor & Francis Group Dictionary of Natural Products. Available online: http://dnp.chemnetbase.com/faces/chemical/ChemicalSearch.xhtml;jsessionid=5F8829606AC568E4B726E513EEA0E9CB (accessed on 18 October 2019).
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
Sample Availability: LC-HRMS/MS raw data can be made available to interested researchers upon request. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites Annotation | ESI+ | ESI- | ESI(±) | Total |
|---|---|---|---|---|
| -By MS and MS/MS, metabolites already reported in V. album species | 13 | 20 | 9 | 42 |
| -By MS and MS/MS, metabolites not previously reported in V. album species | 93 | 42 | 7 | 142 |
| -By MS only, metabolites already reported in V. album species | 14 | 6 | 8 | 28 |
| Total | 120 | 68 | 24 | 212 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peñaloza, E.; Holandino, C.; Scherr, C.; Araujo, P.I.P.d.; Borges, R.M.; Urech, K.; Baumgartner, S.; Garrett, R. Comprehensive Metabolome Analysis of Fermented Aqueous Extracts of Viscum album L. by Liquid Chromatography−High Resolution Tandem Mass Spectrometry. Molecules 2020, 25, 4006. https://doi.org/10.3390/molecules25174006
Peñaloza E, Holandino C, Scherr C, Araujo PIPd, Borges RM, Urech K, Baumgartner S, Garrett R. Comprehensive Metabolome Analysis of Fermented Aqueous Extracts of Viscum album L. by Liquid Chromatography−High Resolution Tandem Mass Spectrometry. Molecules. 2020; 25(17):4006. https://doi.org/10.3390/molecules25174006
Chicago/Turabian StylePeñaloza, Evelyn, Carla Holandino, Claudia Scherr, Paula I. P. de Araujo, Ricardo M. Borges, Konrad Urech, Stephan Baumgartner, and Rafael Garrett. 2020. "Comprehensive Metabolome Analysis of Fermented Aqueous Extracts of Viscum album L. by Liquid Chromatography−High Resolution Tandem Mass Spectrometry" Molecules 25, no. 17: 4006. https://doi.org/10.3390/molecules25174006
APA StylePeñaloza, E., Holandino, C., Scherr, C., Araujo, P. I. P. d., Borges, R. M., Urech, K., Baumgartner, S., & Garrett, R. (2020). Comprehensive Metabolome Analysis of Fermented Aqueous Extracts of Viscum album L. by Liquid Chromatography−High Resolution Tandem Mass Spectrometry. Molecules, 25(17), 4006. https://doi.org/10.3390/molecules25174006