
Phytochemical Characterization and In Vitro Antioxidant Properties of Four Brassica Wild Species from Italy

 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
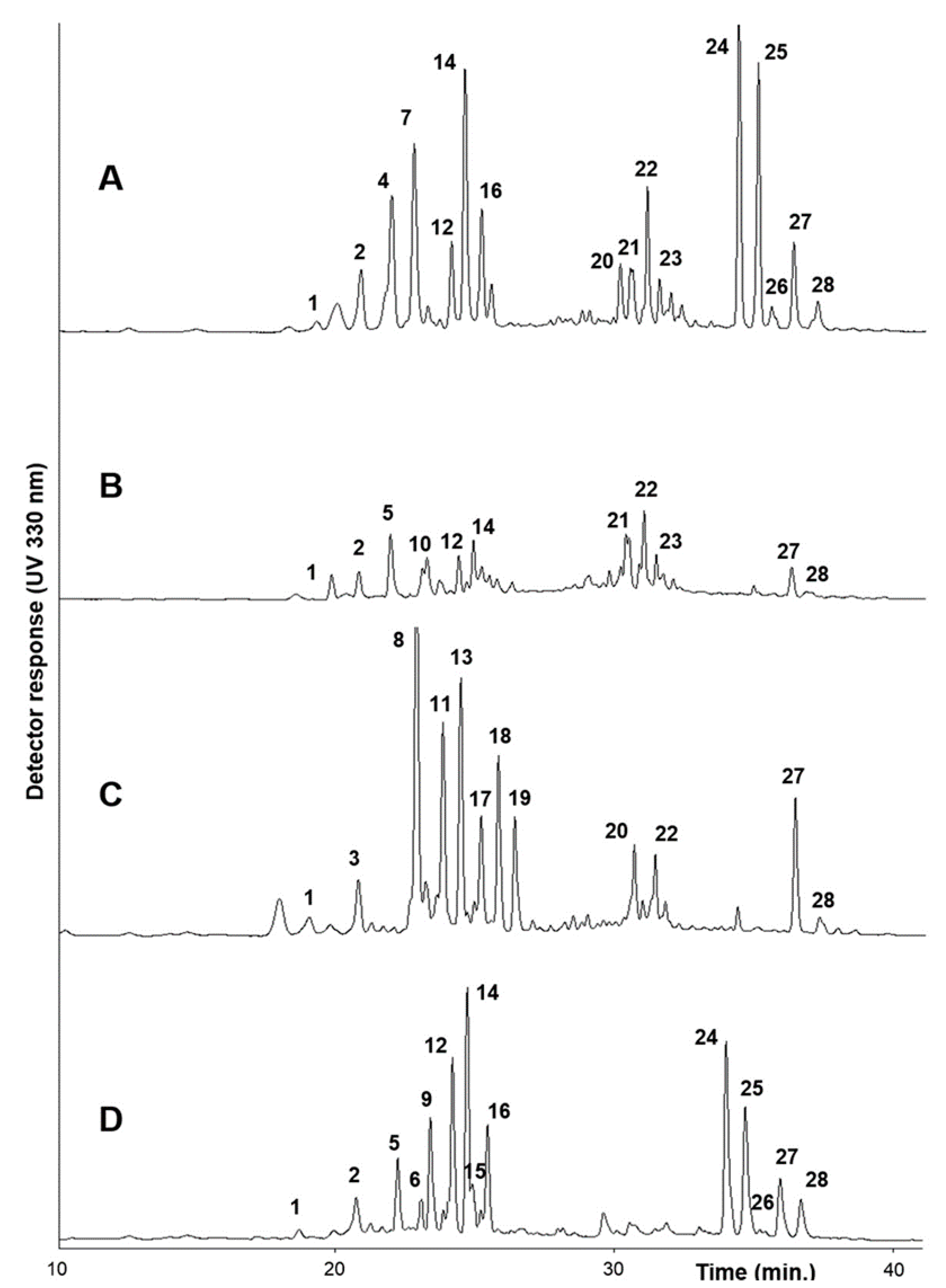
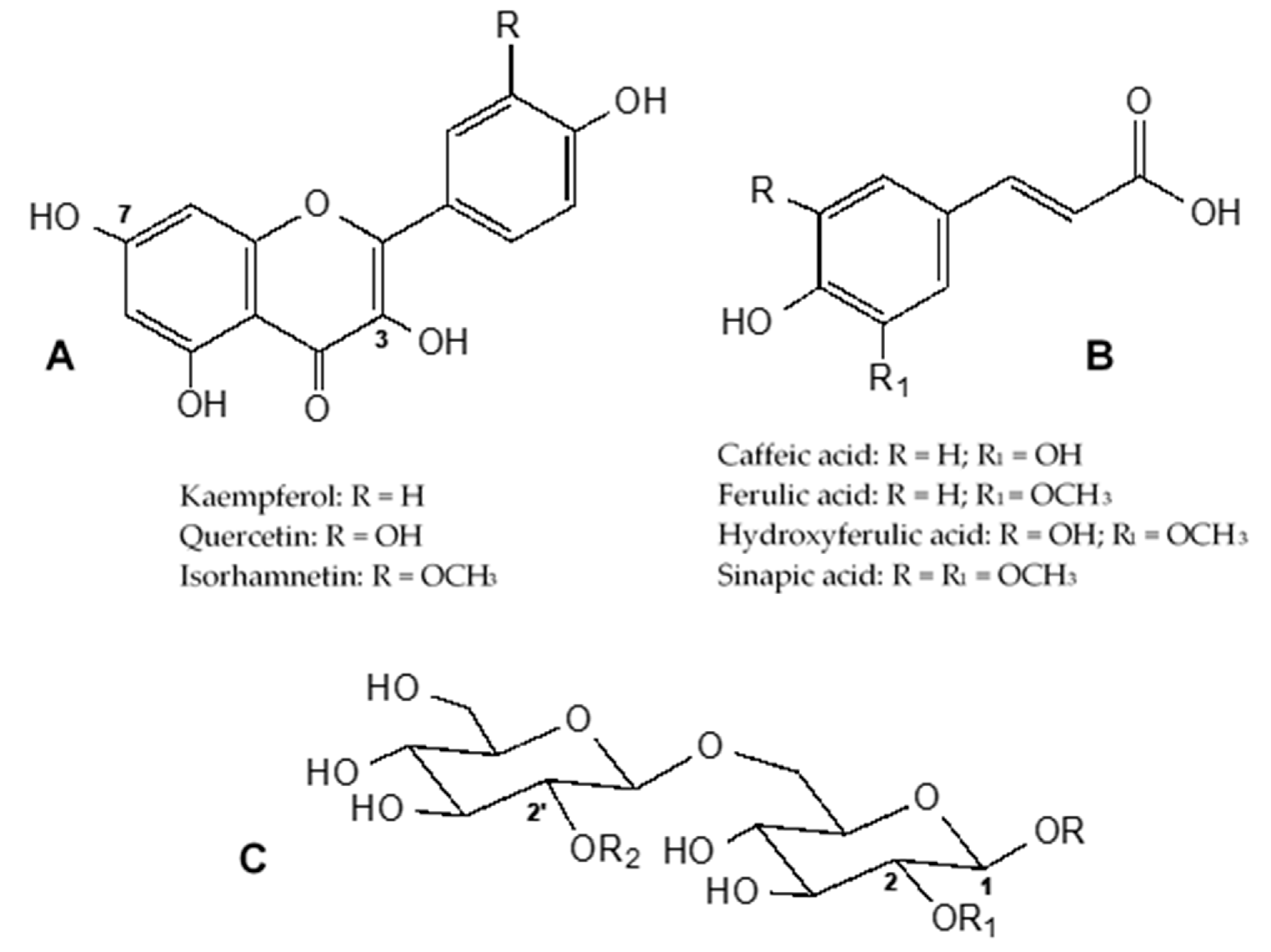
2.1. Phenolic Content and Composition
2.2. Quantification of Phytochemicals and their Fractionation
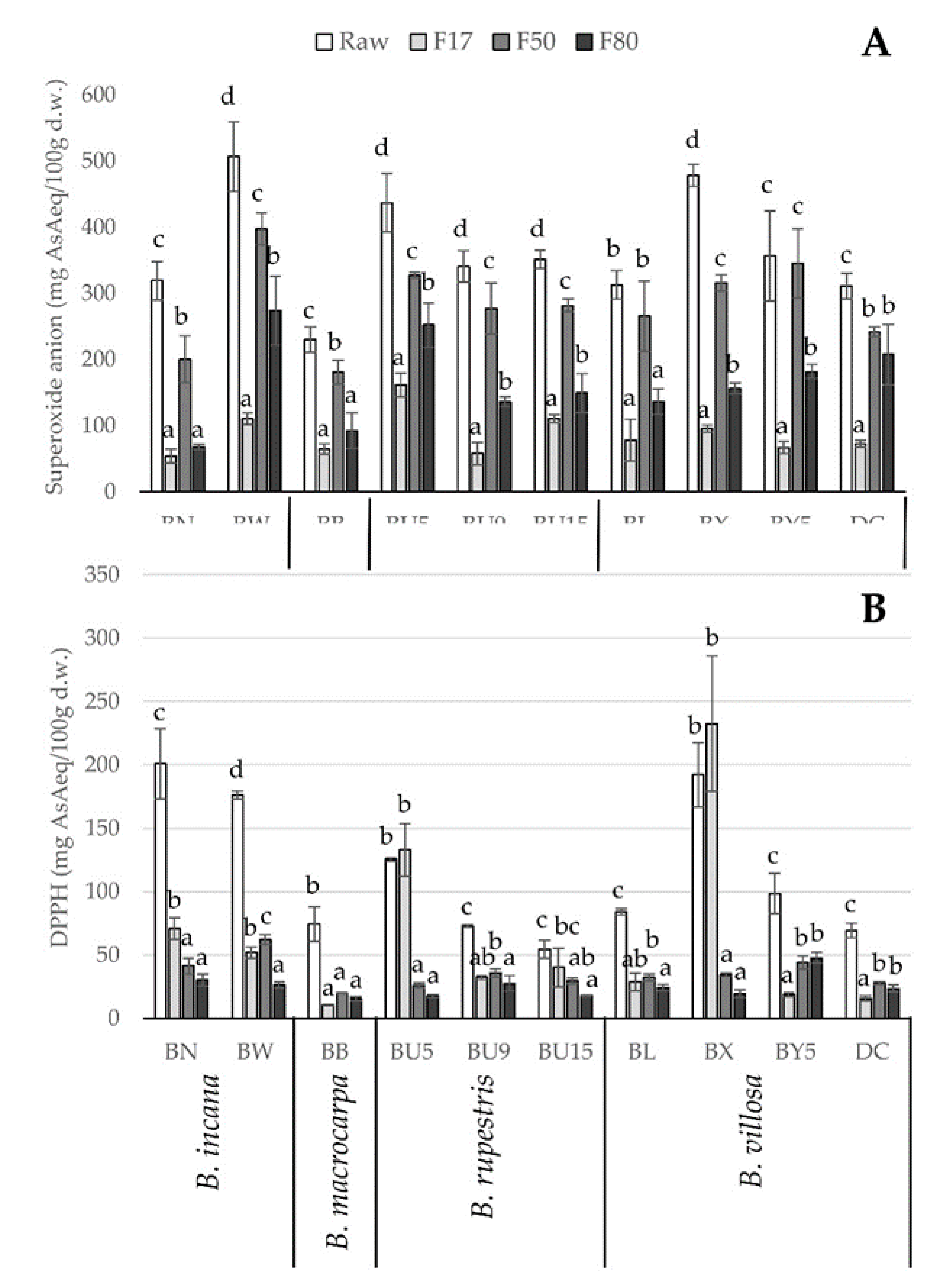
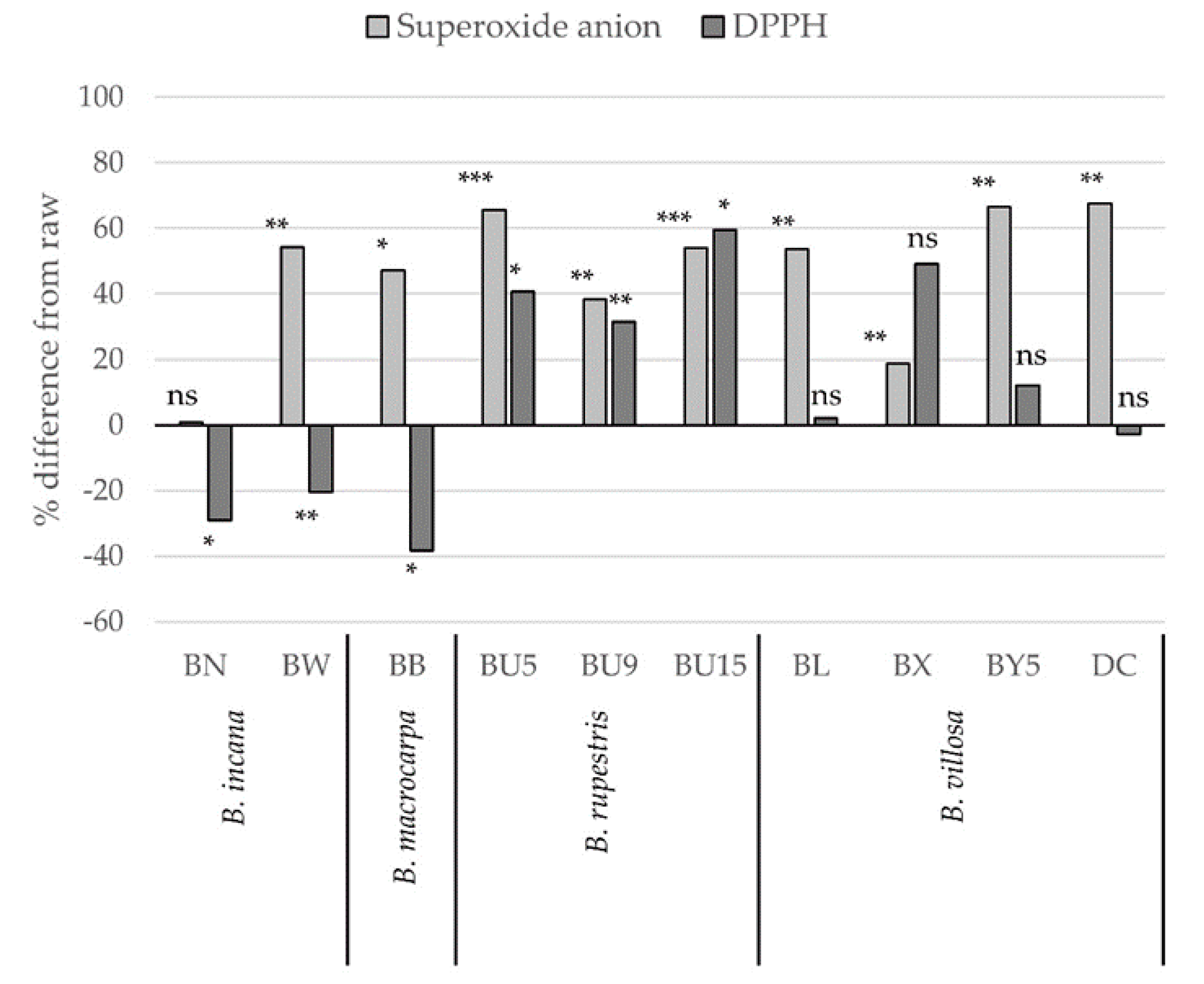
2.3. Antioxidant Capacity
3. Materials and Methods
3.1. Plant Material
3.2. Extraction and Fractionation
3.3. LC/UV–DAD/ESI–MS Analysis
3.4. HPLC-DAD Analyses
3.5. Ascorbic Acid Analysis
3.6. Analysis of Thiol Groups
3.7. Antioxidant Capacity Measurement
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rakow, G. Species origin and economic importance of Brassica. In Brassica. Biotechnology in Agriculture and Forestry; Pua, E.C., Douglas, C.J., Eds.; Springer: Berlin, Germany, 2004; Volume 54, pp. 3–11. [Google Scholar] [CrossRef]
- Bojórquez-Velázquez, E.; Velarde-Salcedo, A.J.; de León-Rodríguez, A.; Jimenez-Islas, H.; Pérez-Torres, J.L.; Herrera-Estrella, A.; Espitia-Rangel, E.; de la Rosa, A.P.B. Morphological, proximal composition, and bioactive compounds characterization of wild and cultivated amaranth (Amaranthus spp.) species. J. Cereal Sci. 2011, 83, 222–228. [Google Scholar] [CrossRef]
- Aguinagalde, I.; Gomez-Campo, C.; Sanchez-Yelamo, M.D. A chemosystematic survey on wild relatives of Brassica oleracea L. Bot. J. Linn. Soc. 1992, 109, 57–67. [Google Scholar] [CrossRef]
- Sánchez-Mata, M.C.; Cabrera Loera, R.D.; Morales, P.; Fernández-Ruiz, V.; Cámara, M.; Díez Marqués, C.; Pardo-de-Santayana, M.; Tardío, J. Wild vegetables of the Mediterranean area as valuable sources of bioactive compounds. Genet. Resour. Crop. Evol. 2012, 59, 431–443. [Google Scholar] [CrossRef]
- Castillo-Lorenzo, E.; Finch-Savage, W.E.; Seal, C.E.; Pritchard, H.W. Adaptive significance of functional germination traits in crop wild relatives of Brassica. Agric. Meteorol. 2019, 264, 343–350. [Google Scholar] [CrossRef]
- Podsędek, A.; Sosnowska, D.; Redzynia, M.; Anders, B. Antioxidant capacity and content of Brassica oleracea dietary antioxidants. Int. J. Food Sci. Technol. 2006, 41, 49–58. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomás-Barberán, F.A.; Garciá-Viguera, C. Health-promoting compounds in broccoli as affected by refrigerated transport and retail sale period. J. Agric. Food. Chem. 2003, 51, 3029–3034. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef]
- Podsędek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Vincieri, F.F.; Romani, A. Antiradical activity and polyphenol composition of local Brassicaceae edible varieties. Food Chem. 2006, 99, 464–469. [Google Scholar] [CrossRef]
- Francisco, M.; Moreno, D.A.; Cartea, M.E.; Ferreres, F.; García-Viguera, C.; Velasco, P. Simultaneous identification of glucosinolates and phenolic compounds in a representative collection of vegetable Brassica rapa. J. Chromatogr. A 2009, 1216, 6611–6619. [Google Scholar] [CrossRef]
- Gratacoós-Cubarsí, M.; Ribas-Agustí, A.; García-Regueiro, J.A.; Castellari, M. Simultaneous evaluation of intact glucosinolates and phenolic compounds by UPLC-DAD-MS/MS in Brassica oleracea L. var. botrytis. Food Chem. 2010, 121, 257–263. [Google Scholar] [CrossRef]
- Nielsen, J.K.; Olsen, C.E.; Petersen, M.K. Acylated flavonol glycosides from cabbage leaves. Phytochemistry 1993, 34, 539–544. [Google Scholar] [CrossRef]
- Llorach, R.; Espin, J.C.; Tomás-Barberán, F.A.; Ferreres, F. Valorization of cauliflower (Brassica oleracea L. var. botrytis) by-products as a source of antioxidant phenolics. J. Agric. Food. Chem. 2003, 51, 2181–2187. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Harnly, J.M. Identification of the phenolic components of Collard greens, Kale and Chinese broccoli. J. Agric. Food. Chem. 2009, 57, 7401–7408. [Google Scholar] [CrossRef]
- Lin, L.Z.; Harnly, J.M. Phenolic component profiles of Mustard greens, Yu Choy and 15 other Brassica vegetables. J. Agric. Food. Chem. 2010, 58, 6850–6857. [Google Scholar] [CrossRef] [PubMed]
- Fiol, M.; Adermann, S.; Neugart, S.; Rohn, S.; Mügge, C.; Schreiner, M.; Krumbein, A.; Kroh, L.W. Highly glycosylated and acylated flavonols isolated from kale (Brassica oleracea var. sabellica)—Structure–antioxidant activity relationship. Food Res. Int. 2012, 47, 80–89. [Google Scholar] [CrossRef]
- Soengas, P.; Cartea, M.E.; Francisco, M.; Sotelo, T.; Velasco, P. New insights into antioxidant activity of Brassica crops. Food Chem. 2012, 134, 725–733. [Google Scholar] [CrossRef]
- Demirkol, O.; Adams, C.; Ercal, N. Biologically important thiols in various vegetables and fruits. J. Agric. Food. Chem. 2004, 52, 8151–8154. [Google Scholar] [CrossRef]
- Shigenaga, T.; Yamauchi, N.; Funamoto, Y.; Shigyo, M. Effects of heat treatment on an ascorbate–glutathione cycle in stored broccoli (Brassica oleracea L.) florets. Postharvest Biol. Technol. 2005, 38, 152–159. [Google Scholar] [CrossRef]
- Zacharis, C.K.; Tzanavaras, P.D.; Zotou, A. Ethyl propiolate as a post-column derivatization reagent for thiols: Development of a green liquid chromatographic method for the determination of glutathione in vegetables. Anal. Chim. Acta 2011, 690, 122–128. [Google Scholar] [CrossRef]
- Sousa, C.; Valentão, P.; Ferreres, F.; Seabra, R.M.; Andrade, P.B. Tronchuda cabbage (Brassica oleracea L. var. costata DC): Scavenger of reactive nitrogen species. J. Agric. Food. Chem. 2008, 56, 4205–4211. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, R.; Sehwag, S.; Singh, S.P. Antioxidant activity and polyphenol content of Brassica oleracea varieties. Int. J. Veg. Sci. 2016, 22, 353–363. [Google Scholar] [CrossRef]
- Branca, F.; Ragusa, L.; Tribulato, A.; Di Gaetano, C.; Calì, F. Genetic relationships of Brassica vegetables and wild relatives in Southern Italy determined by five SSR. Acta Hortic. 2013, 1005, 189–196. [Google Scholar] [CrossRef]
- Branca, F.; Ragusa, L.; Tribulato, A.; Bagatta, M.; Lo Scalzo, R.; Picchi, V. Evaluation of sicilian wild brassica species (n=9) for glucosinolate profile and antioxidant compounds. Acta Hortic. 2013, 1005, 181–188. [Google Scholar] [CrossRef]
- Ferreres, F.; Sousa, C.; Pereira, D.M.; Valentao, P.; Taveira, M.; Martins, A.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Screening of antioxidant phenolic compounds produced by in vitro shoots of Brassica oleracea L. var. costata DC. Comb. Chem. High Throughput Screen. 2009, 12, 230–240. [Google Scholar] [CrossRef]
- Lin, L.Z.; Sun, J.; Chen, P.; Harny, J. UHPLC-PDA-ESI/HRMS/MSn analysis of anthocyanins, flavonol, glucosides, and hydroxycinnamic acid derivatives in red mustard greens (Brassica juncea Coss variety). J. Agric. Food. Chem. 2011, 59, 12059–12072. [Google Scholar] [CrossRef]
- Velasco, P.; Francisco, M.; Moreno, D.A.; Ferreres, F.; Garcia-Viguera, C.; Cartea, M.E. Phytochemical fingerprinting of vegetable Brassica oleracea and Brassica napus by simultaneous identification of glucosinolates and phenolics. Phytochem. Anal. 2011, 22, 144–152. [Google Scholar] [CrossRef]
- Yang, S.C.; Arasu, M.V.; Chun, J.H.; Jang, Y.S.; Lee, Y.H.; Kim, I.H.; Lee, K.T.; Hong, S.T.; Kim, S.J. Identification and determination of phenolic compounds in rapeseed meals (Brassica napus L.). J. Agric. Chem. Env. 2015, 4, 14–23. [Google Scholar] [CrossRef]
- Romani, A.; Vignolini, P.; Isolani, L.; Ieri, F.; Heimler, D. HPLC-DAD/MS characterization of flavonoids and hydroxycinnamic derivatives in turnip tops (Brassica rapa L. subsp. sylvestris L.). J. Agric. Food. Chem. 2006, 54, 1342–1346. [Google Scholar] [CrossRef]
- Miceli, N.; Cavò, E.; Ragusa, M.; Cacciola, F.; Mondello, L.; Dugo, L.; Acquaviva, R.; Malfa, G.A.; Marino, A.; D’Arrigo, M.; et al. Brassica incana Ten. (Brassicaceae): Phenolic constituents, antioxidant and cytotoxic properties of the leaf and flowering top extracts. Molecules 2020, 25, 1461. [Google Scholar] [CrossRef]
- Marín, F.R.; Frutos, M.J.; Pérez-Alvarez, J.A.; Martinez-Sánchez, F.; del Río, J.A. Flavonoids as nutraceuticals: Structural related antioxidant properties and their role on ascorbic acid preservation. Stud. Nat. Prod. Chem. 2002, 26, 741–778. [Google Scholar] [CrossRef]
- Raseetha, S.; Leong, S.Y.; Burritt, D.J.; Indrawati, O. Understanding the degradation of ascorbic acid and glutathione in relation to the levels of oxidative stress biomarkers in broccoli (Brassica oleracea L. italica cv. Bellstar) during storage and mechanical processing. Food Chem. 2013, 138, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.V.; Van Montagu, M.; Inzé, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.J.; Strain, J.J.; Favell, D.; Fletcher, J. Plant L-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Mills, B.J.; Stinson, C.T.; Liu, M.C.; Lang, C.A. Glutathione and cyst(e)ine profiles of vegetables using high performance liquid chromatography with dual electrochemical detection. J. Food Comp. Anal. 1997, 10, 90–101. [Google Scholar] [CrossRef]
- Tabart, J.; Kevers, C.; Pincemail, J.; Defraigne, J.O.; Dommes, J. Comparative antioxidant capacities of phenolic compounds measured by various tests. Food Chem. 2009, 113, 1226–1233. [Google Scholar] [CrossRef]
- Zietz, M.; Weckmüller, A.; Schmidt, S.; Rohn, S.; Schreiner, M.; Krumbein, A.; Kroh, L.W. Genotypic and climatic influence on the antioxidant activity of flavonoids in kale (Brassica oleracea var. sabellica). J. Agric. Food Chem. 2010, 58, 2123–2130. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Rösch, D.; Bergmann, M.; Knorr, D.; Kroh, L.W. Structure antioxidant efficiency relationships of phenolic compounds and their contribution to the antioxidant activity of Sea Buckthorn juice. J. Agric. Food Chem. 2003, 51, 4233–4239. [Google Scholar] [CrossRef]
- Lo Scalzo, R.; Picchi, V.; Migliori, C.A.; Campanelli, G.; Leteo, F.; Ferrari, V.; Di Cesare, L.F. Variations in the phytochemical contents and antioxidant capacity of organically and conventionally grown Italian cauliflower (Brassica oleracea L. subsp. botrytis): Results from a three-year field study. J. Agric. Food Chem. 2013, 61, 10335–10344. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Fujisawa, S.; Kadoma, Y. Anti- and pro-oxidant effects of oxidized quercetin, curcumin or curcumin-related compounds with thiols or ascorbate as measured by the induction period method. In Vivo 2006, 20, 39–44. [Google Scholar] [PubMed]
- Hajimehdipoor, H.; Shahrestani, R.; Shekarchi, M. Investigating the synergistic antioxidant effects of some flavonoid and phenolic compounds. Res. J. Pharm. 2014, 1, 35–40. [Google Scholar]
- Hidalgo, M.; Sánchez-Moreno, C.; de Pascual-Teresa, S. Flavonoid–flavonoid interaction and its effect on their antioxidant activity. Food Chem. 2010, 121, 691–696. [Google Scholar] [CrossRef]
- Kurin, E.; Mučaji, P.; Nagy, M. In vitro antioxidant activities of three red wine polyphenols and their mixtures: An interaction study. Molecules 2012, 17, 14336–14348. [Google Scholar] [CrossRef]
- Rodríguex, I.; Llompart, M.P.; Cela, R. Solid-phase extraction of phenols. J. Chromatogr. A 2000, 885, 291–304. [Google Scholar] [CrossRef]
- Picchi, V.; Migliori, C.A.; Lo Scalzo, R.; Campanelli, G.; Ferrari, V.; di Cesare, L.F. Phytochemical content in organic and conventionally grown Italian cauliflower. Food Chem. 2012, 130, 501–509. [Google Scholar] [CrossRef]
- Hawrylak, B.; Szymanska, M. Selenium as a sulphydrylic group inductor in plant. Cell. Mol. Biol. Lett. 2004, 9, 329–336. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | UV λmax (nm) | [M + H]+/ [M − H]- | MS2 (m/z) (%) | Compound a | B.incana | B. macrocarpa | B. rupestris | B. villosa | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BN | BW | BB | BU5 | BU9 | BU15 | BL | BX | BY5 | DC | ||||||||
| 1 | 235, 263, 319 | 951/949 | 625(100); 301(5) | Q-3-O-diglucoside- 7-O-diglucoside | 2.3 ± 0.0 | 9.6 ± 0.1 | 18.1 ± 0.1 | 6.0 ± 0.4 | 69.2 ± 10.7 | 2.5 ± 0.02 | 13.0 ± 0.2 | 3.6 ± 0.3 | 5.7 ± 0.6 | 6.6 ± 0.0 | |||
| 2 | 239, 263, 338 | 935/933 | 701(100); 285(2) | K-3-O-triglucoside- 7-O-glucoside | 51.2 ± 1.9 | 3.3 ± 0.5 | 23.7 ± 6.6 | - | - | - | 50.2 ± 3.1 | 46.9 ± 2.7 | 80.4 ± 8.6 | 66.9 ± 3.1 | |||
| 3 | 237, 255, 330 | 1275/1273 | 1111(100); 949(43); 787(68); 301(1) | Q-3-O-caffeoyltriglucoside- 7-O-diglucoside | - | - | - | 15.1 ± 1.1 | 59.8 ± 9.1 | 18.8 ± 7.4 | - | - | - | - | |||
| 4 | 239, 267, 331 | 1127/1125 | 801(100); 609(27); 285(1) | K-3-O-hydroxyferuloyl diglucoside-7-O-diglucoside | 52.3 ± 1.9 | 99.7 ± 1.3 | - | - | - | - | - | - | - | - | |||
| 5 | 235, 263, 330 | 935/933 | 609(100);285(2) | K-3-O-diglucoside- 7-O-diglucoside | - | - | 28.1 ± 1.9 | - | - | - | 52.9 ± 3.0 | 36.0 ± 2.2 | 16.3 ± 2.0 | 50.1 ± 1.3 | |||
| 6 | 239, 267, 331 | 995/993 | 831(100); 787(88); 625(56); 301(1) | Q-3-O-sinapoyldiglucoside- 7-O-glucoside | - | - | - | - | - | - | 16.5 ± 0.5 | 9.1 ± 1.2 | 21.2 ± 2.1 | 30.7 ± 0.1 | |||
| 7 | 243, 267, 331 | 1097/1095 | 771(100); 609(17) | K-3-O-caffeoyldiglucoside- 7-O-diglucoside | 62.5 ± 2.1 | 43.5 ± 2.2 | - | - | - | - | - | - | - | - | |||
| 8 | 245, 340 | 1319/1317 | 1111(100); 993(72); 787(87); 301(2) | Q-3-O-sinapoyltriglucoside- 7-O-diglucoside | - | - | - | 54.0 ± 2.3 | 73.7 ± 9.6 | 12.5 ± 0.0 | - | - | - | - | |||
| 9 | 237, 339 | 1157/1155 | 949(100); 831(32); 625(35); 301(1) | Q-3-O-sinapoyldiglucoside- 7-O-diglucoside | - | - | - | - | - | - | 76.1 ± 2.4 | 38.9 ± 2.4 | 17.7 ± 2.6 | 11.6 ± 0.5 | |||
| 10 | 235, 340 | 951/949 | 787(100); 625(52); 301(18) | Q-3-O-caffeoyldiglucoside- 7-O-glucoside | - | - | 9.2 ± 0.6 | - | - | - | - | - | - | - | |||
| 11 | 253, 347 | 1289/1287 | 1111(93); 963(75); 787(100); 301(1) | K-3-O-feruloyltriglucoside- 7-O-diglucoside | - | - | - | 45.4 ± 12.3 | 62.1 ± 9.3 | 74.8 ± 0.2 | - | - | - | - | |||
| 12 | 239, 267, 331 | 979/977 | 815(100); 771(2); 609(7); 285(2) | K-3-O-sinapoyldiglucoside- 7-O-glucoside | 27.7 ± 0.3 | 28.2 ± 0.7 | 7.0 ± 0.1 | - | - | - | 40.0 ± 1.3 | 86.1 ± 6.1 | 24.8 ± 3.7 | 31.3 ± 0.6 | |||
| 13 | 239, 267, 330 | 1303/1301 | 1095(7); 977(100); 771(75); 285(2) | K-3-O-sinapoyltriglucoside- 7-O-diglucoside | - | - | - | 62.0 ± 2.2 | 19.5 ± 2.9 | 36.7 ± 0.4 | - | - | - | - | |||
| 14 | 243, 267, 331 | 1141/1139 | 815(100); 609(28); 285(1) | K-3-O-sinapoyldiglucoside- 7-O-diglucoside | 81.3 ± 1.8 | 67.5 ± 0.7 | 7.7 ± 0.4 | - | - | - | 27.1 ± 0.9 | 81.4 ± 4.4 | 37.1 ± 3.9 | 25.8 ± 1.3 | |||
| 15 | 239, 264, 331 | 949/947 | 785(100); 609(7); 285(2) | K-3-O-feruloyldiglucoside- 7-O-glucoside | - | - | - | - | - | - | 21.5 ± 0.6 | 10.1 ± 1.2 | 43.3 ± 4.2 | 32.1 ± 0.1 | |||
| 16 | 239, 267, 331 | 1111/1109 | 785(100); 609(21); 285(1) | K-3-O-feruloyldiglucoside- 7-O-diglucoside | 38.5 ± 1.1 | 25.3 ± 1.5 | - | - | - | - | 34.3 ± 1.5 | 30.9 ± 1.9 | 30.6 ± 3.0 | 42.3 ± 0.2 | |||
| 17 | 239, 265, 332 | 1273/1271 | 1095(10); 947(100); 771(64); 285(3) | K-3-O-feruloyltriglucoside- 7-O-diglucoside | - | - | - | 19.5 ± 0.1 | 26.2 ± 4.0 | 16.7 ± 0.1 | - | - | - | - | |||
| 18 | 239, 263, 343 | 935/933 | 771(100); 609(10); 285(5) | K-3-O-caffeoyldiglucoside- 7-O-glucoside | - | - | - | 29.4 ± 0.3 | 39.0 ± 5.8 | 18.6 ± 0.3 | - | - | - | - | |||
| 19 | 239, 253, 353 | 965/963 | 801(100); 625(9); 301(3) | Q-3-O-feruloyldiglucoside- 7-O-glucoside | - | - | - | 12.6 ± 0.1 | 31.9 ± 4.8 | 10.6 ± 0.2 | - | - | - | - | |||
| 20 | 239, 269 sh, 329 | 1495/1493 | 1169(100); 609(39); 285(7) | K-3-O-sinapoylhydroxyferuloyl triglucoside-7-O-diglucoside | 10.4 ± 0.3 | 20.0 ± 2.1 | - | 19.1 ± 0.2 | 20.0 ± 2.9 | 11.1 ± 0.0 | - | - | - | - | |||
| 21 | 239, 269 sh, 330 | 1465/1463 | 1139(100); 609(21); 285(3) | K-3-O-feruloylhydroxyferuloyl triglucoside-7-O-diglucoside | 5.6 ± 0.2 | 31.3 ± 2.7 | 30.7 ± 1.8 | - | - | - | - | - | - | - | |||
| 22 | 239, 267, 331 | 1509/1507 | 1301(7); 1183(100); 609(27); 285(5) | K-3-O-disinapoyltriglucoside- 7-O-diglucoside | 22.4 ± 1.3 | 53.0 ± 2.7 | 18.2 ± 1.5 | 18.9 ± 0.3 | 24.0 ± 3.8 | 16.2 ± 1.8 | - | - | - | - | |||
| 23 | 239, 267 sh, 327 | 1479/1477 | 1153(100); 947(25); 609(33); 285(4) | K-3-O-feruloylsinapoyl triglucoside-7-O-diglucoside | 16.6 ± 0.4 | 37.7 ± 3.0 | 10.9 ± 1.3 | - | - | - | - | - | - | - | |||
| 24 | 240, 331 | 777 b/753 | 529(100) | 1,2-disinapoyl gentiobioside | 57.2 ± 0.0 | 51.3 ± 2.4 | - | - | - | - | 27.8 ± 0.7 | 50.6 ± 2.4 | 39.4 ± 3.7 | 17.1 ± 0.2 | |||
| 25 | 240, 327 | 747 b/723 | 529(38); 499(100) | 1-sinapoyl-2-feruloyl gentiobioside | 55.6 ± 0.7 | 55.6 ± 1.3 | - | - | - | - | 15.5 ± 0.1 | 36.9 ± 3.6 | 48.7 ± 3.9 | 33.3 ± 0.7 | |||
| 26 | 239, 294 sh, 327 | 717 b/693 | 499(100) | 1,2-diferuloyl gentiobioside | 9.0 ± 0.0 | 1.6 ± 0.8 | - | - | - | - | 0.94 ± 0.0 | 0.7 ± 0.0 | 45.4 ± 4.2 | 32.4 ± 0.8 | |||
| 27 | 240, 327 | 983 b/959 | 735(100); 511(21) | 1,2,2′-trisinapoyl gentiobioside | 20.9 ± 0.7 | 5.4 ± 1.2 | 8.9 ± 0.7 | 34.5 ± 0.5 | 28.1 ± 4.0 | 18.3 ± 0.3 | 55.1 ± 3.2 | 19.5 ± 0.9 | 45.7 ± 4.2 | 66.3 ± 1.8 | |||
| 28 | 239, 327 | 953 b/929 | 735(98); 705(100); 511(13); 481(7) | 1,2′-disinapoyl-2-feruloyl gentiobioside | 14.8 ± 0.1 | 0.98 ± 0.0 | 1.3 ± 0.1 | 3.4 ± 0.04 | 7.6 ± 1.1 | 0.75 ± 0.0 | 33.7 ± 0.5 | 12.8 ± 2.5 | 8.3 ± 0.7 | 5.2 ± 0.5 | |||
| B.incana | B. macrocarpa | B. rupestris | B. villosa | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BN | BW | BB | BU5 | BU9 | BU15 | BL | BX | BY5 | DC | |||||
| Flavonol | Raw | 370.6 ± 6.8 g | 419.0 ± 17.2 h | 153.7 ± 0.9 a | 282.0 ± 13.7 c | 430.2 ± 30.2 h | 218.6 ± 10.5 b | 325.5 ± 6.1 de | 343.0 ± 22.4 ef | 277.0 ± 61.5 c | 297.4 ± 5.7 cd | |||
| derivatives | F50 | 284.6 ± 38.1 | 275.1 ± 5.0 | 109.1 ± 6.4 | 262.1 ± 10.7 | 152.8 ± 21.0 | 166.9 ± 2.6 | 344.3 ± 17.6 | 338.4 ± 10.5 | 101.3 ± 19.2 | 159.0 ± 19.2 | |||
| F80 | n.d. | 69.9 ± 7.7 | 27.3 ± 2.7 | 33.2 ± 2.1 | 33.5 ± 4.2 | n.d. | 57.5 ± 17.0 | 38.7 ± 6.8 | 76.2 ± 16.4 | 8.3 ± 0.3 | ||||
| Hydroxy- | Raw | 157.4 ± 1.5 e | 114.9 ± 5.8 c | 10.2 ± 0.8 a | 38.0 ± 13.7 b | 37.5 ± 2.6 b | 19.0 ± 0.3 a | 133.0 ± 4.5 d | 120.5 ± 9.4 cd | 187.4 ± 33.4 f | 154.2 ± 2.6 e | |||
| cinnamoyl | F50 | 9.3 ± 0.3 | 21.6 ± 1.8 | 1.7 ± 0.2 | 3.2 ± 0.4 | 8.8 ± 0.8 | 1.0 ± 0.2 | 10.7 ± 2.5 | 10.2 ± 2.2 | 24.8 ± 2.0 | 18.1 ± 8.0 | |||
| gentiobiosides | F80 | 113.0 ± 13.3 | 93.7 ± 7.3 | 8.9 ± 0.6 | 32.4 ± 0.2 | 51.6 ± 7.7 | 18.3 ± 0.6 | 91.1 ± 4.2 | 121.0 ± 3.0 | 181.0 ± 20.4 | 116.4 ± 4.2 | |||
| Total phenols | Raw | 531.0 ± 4.2 e | 533.9 ± 23.0 e | 163.9 ± 1.8 a | 319.9 ± 13.2 c | 444.5 ± 65.6 d | 237.6 ± 10.7 b | 454.7 ± 5.3 d | 463.5 ± 31.8 d | 464.4 ± 94.9 d | 451.6 ± 8.3 d | |||
| AsA | Raw | 355.7 ± 60.8 f | 249.2 ± 42.6 e | 103.9 ± 17.8 b | 214.8 ± 36.7 de | 30.1 ± 12.2 a | 119.8 ± 20.5 bc | 167.0 ± 28.5 cd | 375.8 ± 64.2 f | 7.6 ± 1.3 a | 38.2 ± 6.5 a | |||
| F17 | 122.8 ± 3.5 | 96.5 ± 3.9 | 37.0 ± 10.5 | 101.5 ± 6.5 | 14.6 ± 0.2 | 70.0 ± 7.2 | 76.4 ± 21.9 | 131.0 ± 9.1 | 1.1 ± 0.1 | 5.7 ± 2.1 | ||||
| %* | - 65.5 | - 61.3 | - 64.4 | - 52.8 | - 51.6 | - 41.6 | - 54.2 | - 65.1 | - 85.2 | - 85.1 | ||||
| Biothiols | Raw | 3.83 ± 1.02 d | 1.88 ± 0.5 c | 0.59 ± 0.16 a | 1.19 ± 0.45 abc | 1.7 ± 0.0 bc | 1.02 ± 0.27 abc | 0.89 ± 0.24 abc | 5.13 ± 1.37 e | 0.67 ± 0.18 ab | 0.83 ± 0.22 ab | |||
| Species | Code | Origin | Coordinates | Accession Code |
|---|---|---|---|---|
| B. incana | BN | Agnone Bagni (SR) | 37°19′ N 15°05′ E | UNICT4419 |
| BW | Sortino (SR) | 37°10′ N 15°20′ E | UNICT4158 | |
| B. macrocarpa | BB | Favignana (TP) | 37°55′ N 12°19′ E | UNICT4801 |
| B. rupestris | BU5 | Roccella Valdemone (ME) | 37°56′ N 15°10′ E | UNICT3405 |
| BU9 | Ragusa Ibla (RG) | 36°56′ N 14°45′ E | UNICT3458 | |
| BU15 | Stilo (RC) | 38°29′ N 16°28′ E | UNICT3411 | |
| B. villosa | BL | Caltabellotta (AG) | 37°34′ N 13°13′ E | UNICT5031 |
| BX | Marianopoli (CL) | 37°36′ N 13°55′ E | UNICT3944 | |
| BY5 | Agnone Bagni (SR) | 37°19′ N 15°05′ E | UNICT4581 | |
| DC | Lago Albano (RM) | 41°45′ N12°39′ E | UNICT4202 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Picchi, V.; Lo Scalzo, R.; Tava, A.; Doria, F.; Argento, S.; Toscano, S.; Treccarichi, S.; Branca, F. Phytochemical Characterization and In Vitro Antioxidant Properties of Four Brassica Wild Species from Italy. Molecules 2020, 25, 3495. https://doi.org/10.3390/molecules25153495
Picchi V, Lo Scalzo R, Tava A, Doria F, Argento S, Toscano S, Treccarichi S, Branca F. Phytochemical Characterization and In Vitro Antioxidant Properties of Four Brassica Wild Species from Italy. Molecules. 2020; 25(15):3495. https://doi.org/10.3390/molecules25153495
Chicago/Turabian StylePicchi, Valentina, Roberto Lo Scalzo, Aldo Tava, Filippo Doria, Sergio Argento, Stefania Toscano, Simone Treccarichi, and Ferdinando Branca. 2020. "Phytochemical Characterization and In Vitro Antioxidant Properties of Four Brassica Wild Species from Italy" Molecules 25, no. 15: 3495. https://doi.org/10.3390/molecules25153495
APA StylePicchi, V., Lo Scalzo, R., Tava, A., Doria, F., Argento, S., Toscano, S., Treccarichi, S., & Branca, F. (2020). Phytochemical Characterization and In Vitro Antioxidant Properties of Four Brassica Wild Species from Italy. Molecules, 25(15), 3495. https://doi.org/10.3390/molecules25153495