
Gold Nanoparticles for Vectorization of Nucleic Acids for Cancer Therapeutics


 and
and 
Abstract
1. Introduction
1.1. Cancer: Worldwide Statistics; Biomarker Detection and Treatment Strategies
1.2. Gene Therapy in Cancer
1.2.1. Genome Editing Tools
CRISPR/Cas9
ZFNs-Zinc Finger Nucleases
TALENs-Transcription Activator-Like Effector Nucleases
Meganucleases
1.2.2. Gene Silencing—RNAi; ASOs; Ribozymes and Riboswitches
RNAi Mechanisms and ASOs
Ribozymes and Riboswitches
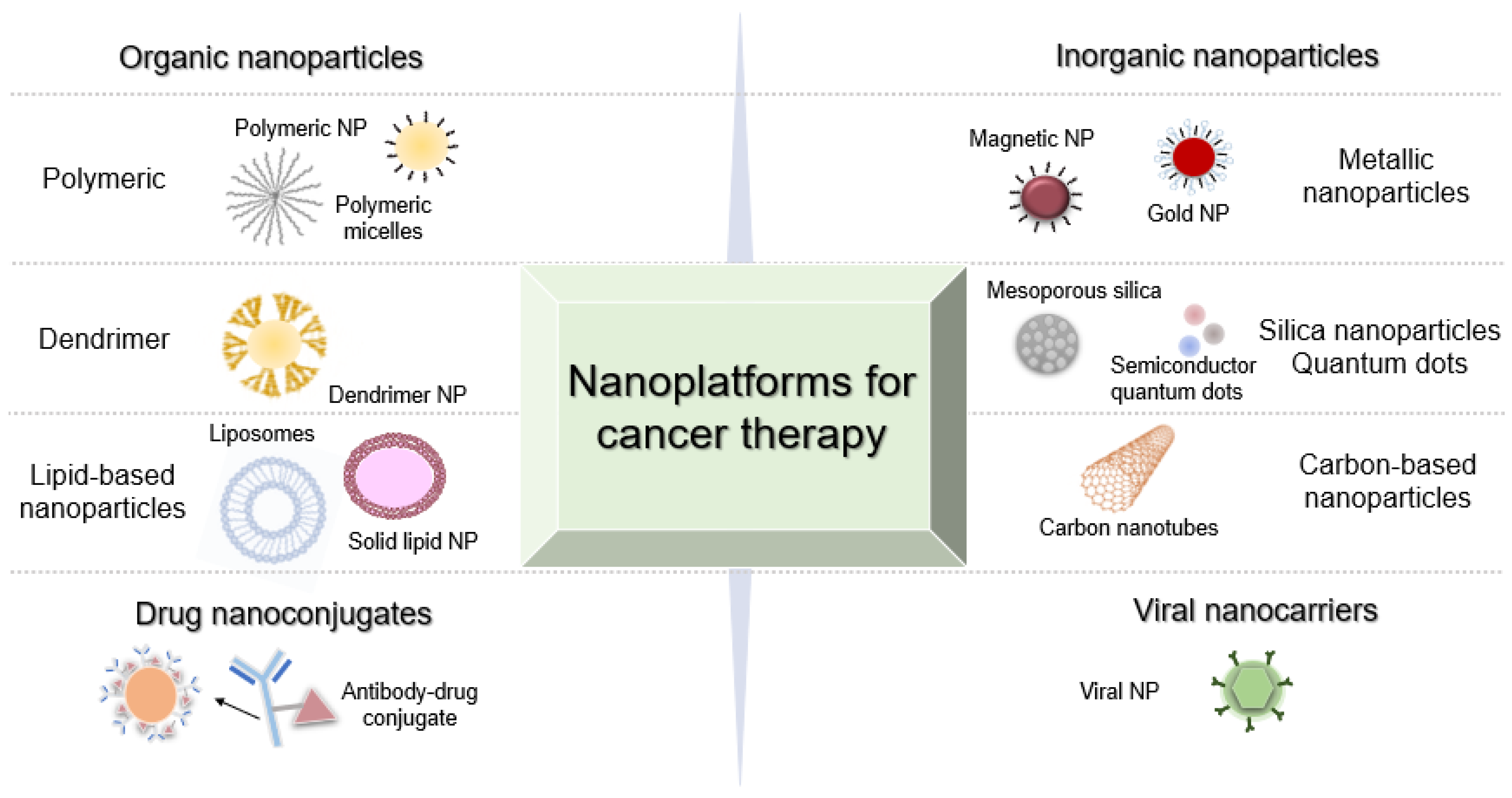
2. Molecular Nanomedicines against Cancer
2.1. Metal NPs for Gene Silencing
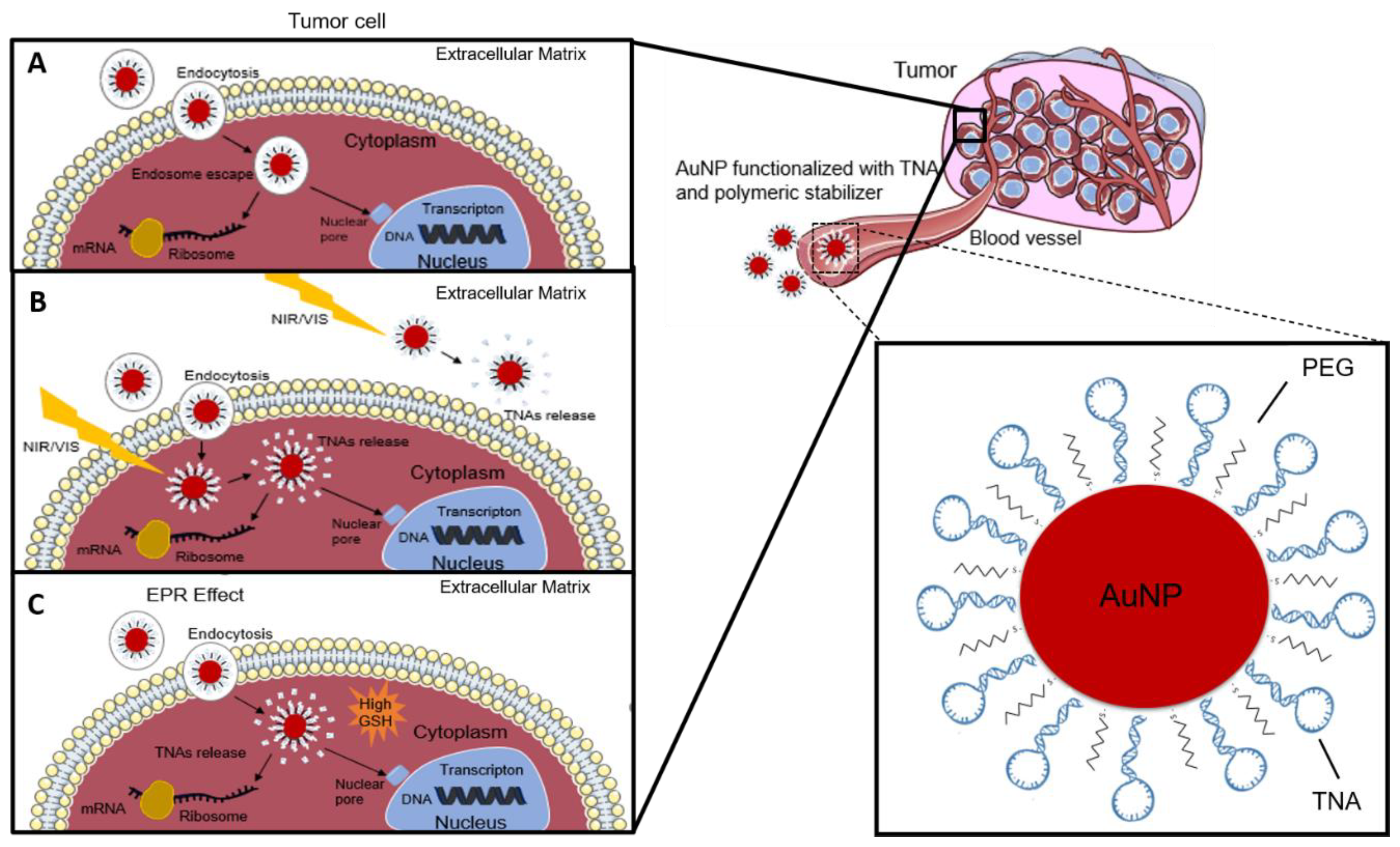
AuNPs for Nucleic Acid Delivery
2.2. Translation to the Clinic
3. Conclusions and Future Perspectives
Funding
Conflicts of Interest
References
- WHO: World Health Organization. Available online: http://www.who.int/en/ (accessed on 26 February 2020).
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qu, X. Cancer Biomarker Detection: Recent Achievements and Challenges. Chem. Soc. Rev. 2015, 44, 2963–2997. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Gamagedara, S. Biomarker Analysis for Oncology. Biomark. Med. 2015, 9, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Kalia, M. Biomarkers for Personalized Oncology: Recent Advances and Future Challenges. Metabolism 2015, 64, S16–S21. [Google Scholar] [CrossRef] [PubMed]
- Henry, N.L.; Hayes, D.F. Cancer Biomarkers. Mol. Oncol. 2012, 6, 140–146. [Google Scholar] [CrossRef]
- Roma-Rodrigues, C.; Rivas-García, L.; Baptista, P.V.; Fernandes, A.R. Gene Therapy in Cancer Treatment: Why Go Nano? Pharmaceutics 2020, 12, 233. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Shukla, P. Gene Editing for Cell Engineering: Trends and Applications. Crit. Rev. Biotechnol. 2017, 37, 672–684. [Google Scholar] [CrossRef]
- Carroll, D. Genome Editing: Past, Present, and Future. Yale J. Biol. Med. 2017, 90, 653–659. [Google Scholar]
- Moon, S.B.; Kim, D.Y.; Ko, J.H.; Kim, Y.S. Recent Advances in the CRISPR Genome Editing Tool Set. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakatura, A. Nucleotide Sequence of the Iap Gene, Responsible for Alkaline Phosphatase Isoenzyme Conversion in Escherichia Coli, and Identification of the Gene Product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An Updated Evolutionary Classification of CRISPR-Cas Systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Bikard, D.; Cox, D.; Zhang, F.; Marraffini, L.A. RNA-Guided Editing of Bacterial Genomes Using CRISPR-Cas Systems. Nat. Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, L.; Liu, H.; Cheng, K. Delivery Strategies of the CRISPR-Cas9 Gene-Editing System for Therapeutic Applications. J. Control. Release 2017, 266, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Biagioni, A.; Laurenzana, A.; Margheri, F.; Chillà, A.; Fibbi, G.; So, M. Delivery Systems of CRISPR/Cas9-Based Cancer Gene Therapy 11 Medical and Health Sciences 1112 Oncology and Carcinogenesis. J. Biol. Eng. 2018, 12. [Google Scholar] [CrossRef]
- Kim, Y.G.; Cha, J.; Chandrasegaran, S. Hybrid Restriction Enzymes: Zinc Finger Fusions to Fok I Cleavage Domain. Proc. Natl. Acad. Sci. USA 1996, 93, 1156–1160. [Google Scholar] [CrossRef] [PubMed]
- Fairall, L.; Schwabe, J.W.R.; Chapman, L.; Finch, J.T.; Rhodes, D. The Crystal Structure of a Two Zinc-Finger Peptide Reveals an Extension to the Rules for Zinc-Finger/DNA Recognition. Nature 1993, 366, 483–487. [Google Scholar] [CrossRef]
- Guo, J.; Gaj, T.; Barbas, C.F. Directed Evolution of an Enhanced and Highly Efficient FokI Cleavage Domain for Zinc Finger Nucleases. J. Mol. Biol. 2010, 400, 96–107. [Google Scholar] [CrossRef]
- Paschon, D.E.; Lussier, S.; Wangzor, T.; Xia, D.F.; Li, P.W.; Hinkley, S.J.; Scarlott, N.A.; Lam, S.C.; Waite, A.J.; Truong, L.N.; et al. Diversifying the Structure of Zinc Finger Nucleases for High-Precision Genome Editing. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Bogdanove, A.J.; Voytas, D.F. TAL Effectors: Customizable Proteins for DNA Targeting. Science 2011, 333, 1843–1846. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Huang, S.; Jiang, W.Z.; Wright, D.; Spalding, M.H.; Weeks, D.P.; Yang, B. TAL Nucleases (TALNs): Hybrid Proteins Composed of TAL Effectors and FokI DNA-Cleavage Domain. Nucleic Acids Res. 2011, 39, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Cermak, T.; Doyle, E.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.; Somia, N.; Bogdanove, A.; Voytas, D. Efficient Design and Assembly of Custom TALEN and Other TAL Effector-Based Constructs for DNA Targeting. Nucleic Acids Res. 2011, 12, e82. [Google Scholar] [CrossRef] [PubMed]
- Reyon, D.; Tsai, S.Q.; Khgayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH Assembly of TALENs for High-Throughput Genome Editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Burgk, J.L.; Schmidt, T.; Kaiser, V.; Höning, K.; Hornung, V. A Ligation-Independent Cloning Technique for High-Throughput Assembly of Transcription Activator-like Effector Genes. Nat. Biotechnol. 2013, 31, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.; Poirot, L.; Galetto, R.; Smith, J.; Montoya, G.; Duchateau, P.; Paques, F. Meganucleases and Other Tools for Targeted Genome Engineering: Perspectives and Challenges for Gene Therapy. Curr. Gene Ther. 2011, 11, 11–27. [Google Scholar] [CrossRef]
- Khan, S.H. Genome-Editing Technologies: Concept, Pros, and Cons of Various Genome-Editing Techniques and Bioethical Concerns for Clinical Application. Mol. Ther. -Nucleic Acids 2019, 16, 326–334. [Google Scholar] [CrossRef]
- Fernandes, A.R.; Baptista, P.V. Gene Silencing Using Multifunctionalized Gold Nanoparticles for Cancer Therapy. Methods Mol. Biol. 2017, 1530, 319–336. [Google Scholar] [CrossRef]
- Kim, D.H.; Rossi, J.J. RNAi Mechanisms and Applications. Biotechniques 2008, 44, 613–616. [Google Scholar] [CrossRef]
- Chery, J. RNA Therapeutics: RNAi and Antisense Mechanisms and Clinical Applications. Postdoc J. 2016, 4. [Google Scholar] [CrossRef]
- Rupp, L.J.; Schumann, K.; Roybal, K.T.; Gate, R.E.; Ye, C.J.; Lim, W.A.; Marson, A. CRISPR/Cas9-Mediated PD-1 Disruption Enhances Anti-Tumor Efficacy of Human Chimeric Antigen Receptor T Cells. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, S.; Hackett, C.S.; Brentjens, R.J. Engineering Strategies to Overcome the Current Roadblocks in CAR T Cell Therapy. Nat. Rev. Clin. Oncol. 2020, 17, 147–167. [Google Scholar] [CrossRef] [PubMed]
- Depil, S.; Duchateau, P.; Grupp, S.A.; Mufti, G.; Poirot, L. ‘Off-the-Shelf’ Allogeneic CAR T Cells: Development and Challenges. Nat. Rev. Drug Discov. 2020, 19, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Serganov, A.; Patel, D.J. Ribozymes, Riboswitches and beyond: Regulation of Gene Expression without Proteins. Nat. Rev. Genet. 2007, 8, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Karimian, A.; Azizian, K.; Parsian, H.; Rafieian, S.; Shafiei-Irannejad, V.; Kheyrollah, M.; Yousefi, M.; Majidinia, M.; Yousefi, B. CRISPR/Cas9 Technology as a Potent Molecular Tool for Gene Therapy. J. Cell. Physiol. 2019, 234, 12267–12277. [Google Scholar] [CrossRef] [PubMed]
- Givens, B.E.; Naguib, Y.W.; Geary, S.M.; Devor, E.J.; Salem, A.K. Nanoparticle-Based Delivery of CRISPR/Cas9 Genome-Editing Therapeutics. AAPS J. 2018, 20, 108. [Google Scholar] [CrossRef] [PubMed]
- Knott, G.J.; Doudna, J.A. CRISPR-Cas Guides the Future of Genetic Engineering. Science 2018, 361, 866–869. [Google Scholar] [CrossRef]
- Lee, K.; Conboy, M.; Park, H.M.; Jiang, F.; Kim, H.J.; Dewitt, M.A.; Mackley, V.A.; Chang, K.; Rao, A.; Skinner, C.; et al. Nanoparticle Delivery of Cas9 Ribonucleoprotein and Donor DNA in Vivo Induces Homology-Directed DNA Repair. Nat. Biomed. Eng. 2017, 1, 889–901. [Google Scholar] [CrossRef]
- Li, H.; Yang, Y.; Hong, W.; Huang, M.; Wu, M.; Zhao, X. Applications of Genome Editing Technology in the Targeted Therapy of Human Diseases: Mechanisms, Advances and Prospects. Signal Transduct. Target. Ther. 2020, 5. [Google Scholar] [CrossRef]
- Stadtmauer, E.A.; Fraietta, J.A.; Davis, M.M.; Cohen, A.D.; Weber, K.L.; Lancaster, E.; Mangan, P.A.; Kulikovskaya, I.; Gupta, M.; Chen, F.; et al. CRISPR-Engineered T Cells in Patients with Refractory Cancer. Science 2020, 367. [Google Scholar] [CrossRef]
- Zhan, T.; Rindtorff, N.; Betge, J.; Ebert, M.P.; Boutros, M. CRISPR/Cas9 for Cancer Research and Therapy. Semin. Cancer Biol. 2019, 55, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Li, L.; Wang, X. Molecular Mechanisms, off-Target Activities, and Clinical Potentials of Genome Editing Systems. Clin. Transl. Med. 2020, 10, 412–426. [Google Scholar] [CrossRef] [PubMed]
- Kher, G.; Trehan, S.; Misra, A. Antisense Oligonucleotides and RNA Interference, 1st ed.; Elsevier Inc.: Burlington, MA, USA, 2011. [Google Scholar] [CrossRef]
- Ryou, S.M.; Kim, J.M.; Yeom, J.H.; Hyun, S.; Kim, S.; Han, M.S.; Kim, S.W.; Bae, J.; Rhee, S.; Lee, K. Gold Nanoparticle-Assisted Delivery of Small, Highly Structured RNA into the Nuclei of Human Cells. Biochem. Biophys. Res. Commun. 2011, 416, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Senapati, D.; Patra, B.C.; Kar, A.; Chini, D.S.; Ghosh, S.; Patra, S.; Bhattacharya, M. Promising Approaches of Small Interfering RNAs (SiRNAs) Mediated Cancer Gene Therapy. Gene 2019, 719. [Google Scholar] [CrossRef]
- Kim, J.H.; Yeom, J.H.; Ko, J.J.; Han, M.S.; Lee, K.; Na, S.Y.; Bae, J. Effective Delivery of Anti-MiRNA DNA Oligonucleotides by Functionalized Gold Nanoparticles. J. Biotechnol. 2011, 155, 287–292. [Google Scholar] [CrossRef]
- Collin, R.W.J.; Garanto, A. Applications of Antisense Oligonucleotides for the Treatment of Inherited Retinal Diseases. Curr. Opin. Ophthalmol. 2017, 28, 260–266. [Google Scholar] [CrossRef]
- Wicki, A.; Witzigmann, D.; Balasubramanian, V.; Huwyler, J. Nanomedicine in Cancer Therapy: Challenges, Opportunities, and Clinical Applications. J. Control. Release 2015, 200, 138–157. [Google Scholar] [CrossRef]
- Ferrari, M. Cancer Nanotechnology: Opportunities and Challenges. Nat. Rev. Cancer 2005, 5, 161–171. [Google Scholar] [CrossRef]
- Peer, D.; Karp, J.M.; Hong, S.; Farokhzad, O.C.; Margalit, R.; Langer, R. Nanocarriers as an Emerging Platform for Cancer Therapy. Nat. Nanotechnol. 2007, 2, 751–760. [Google Scholar] [CrossRef]
- Sumer, B.; Gao, J. Theranostic Nanomedicine for Cancer. Nanomedicine 2008, 3, 137–140. [Google Scholar] [CrossRef]
- Kesharwani, P.; Iyer, A.K. Recent Advances in Dendrimer-Based Nanovectors for Tumor-Targeted Drug and Gene Delivery. Drug Discov. Today 2015, 20, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, E.; Aval, S.F.; Akbarzadeh, A.; Milani, M.; Nasrabadi, H.T.; Joo, S.W.; Hanifehpour, Y.; Nejati-Koshki, K.; Pashaei-Asl, R. Dendrimers: Synthesis, Applications, and Properties. Nanoscale Res. Lett. 2014, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wan, T.; Wang, H.; Zhang, S.; Ping, Y.; Cheng, Y. A Boronic Acid–Rich Dendrimer with Robust and Unprecedented Efficiency for Cytosolic Protein Delivery and CRISPR-Cas9 Gene Editing. Sci. Adv. 2019, 5. [Google Scholar] [CrossRef] [PubMed]
- Bartsevich, V.; Jantz, D.; Smith, J.; Nicholson, M. Treatment of Retinitis Pigmentosa Using Engineered Meganucleases. WO2017044649A1. 2017. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2017044649 (accessed on 30 July 2020).
- Pandey, H.; Rani, R.; Agarwal, V. Liposome and Their Applications in Cancer Therapy. Brazilian Arch. Biol. Technol. 2016, 59. [Google Scholar] [CrossRef]
- Celsion Corporation. ThermoDox. Available online: https://celsion.com/thermodox/ (accessed on 4 May 2020).
- Barba, A.A.; Bochicchio, S.; Dalmoro, A.; Lamberti, G. Lipid Delivery Systems for Nucleic-Acid-Based-Drugs: From Production to Clinical Applications. Pharmaceutics 2019, 11, 360. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, S.; Kim, J.H.; Park, K.; Kim, K.; Kwon, I.C. Polymeric Nanomedicine for Cancer Therapy. Prog. Polym. Sci. 2008, 33, 113–137. [Google Scholar] [CrossRef]
- Yadav, H.K.S.; Almokdad, A.A.; shaluf, S.I.M.; Debe, M.S. Polymer-Based Nanomaterials for Drug-Delivery Carriers; Elsevier Inc.: Cambridge, MA USA, 2019. [Google Scholar] [CrossRef]
- Jo, A.; Ringel-Scaia, V.M.; McDaniel, D.K.; Thomas, C.A.; Zhang, R.; Riffle, J.S.; Allen, I.C.; Davis, R.M. Fabrication and Characterization of PLGA Nanoparticles Encapsulating Large CRISPR-Cas9 Plasmid. J. Nanobiotechnol. 2020, 18. [Google Scholar] [CrossRef]
- Saleem, J.; Wang, L.; Chen, C. Carbon-Based Nanomaterials for Cancer Therapy via Targeting Tumor Microenvironment. Adv. Healthc. Mater. 2018, 7, 1–30. [Google Scholar] [CrossRef]
- Ray, S.C.; Jana, N.R. Application of Carbon-Based Nanomaterials as Drug and Gene Delivery Carrier; Elsevier: Cambridge, MA, USA, 2017. [Google Scholar] [CrossRef]
- Luo, T.Y.; He, X.; Zhang, J.; Chen, P.; Liu, Y.H.; Wang, H.J.; Yu, X.Q. Photoluminescent F-Doped Carbon Dots Prepared by Ring-Opening Reaction for Gene Delivery and Cell Imaging. RSC Adv. 2018, 8, 6053–6062. [Google Scholar] [CrossRef]
- Zhang, H.; Yee, D.; Wang, C. Quantum Dots for Cancer Diagnosis and Therapy: Biological and Clinical Perspectives. Nanomedicine 2008, 3, 83–91. [Google Scholar] [CrossRef]
- Wu, F.-G.; Zhang, X.; Chen, X.; Sun, W.; Bao, Y.-W.; Hua, X.-W.; Gao, G.; Jia, H.-R. Quantum Dots for Cancer Therapy and Bioimaging; Springer International Publishing: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Beik, J.; Khateri, M.; Khosravi, Z.; Kamrava, S.K.; Kooranifar, S.; Ghaznavi, H.; Shakeri-Zadeh, A. Gold Nanoparticles in Combinatorial Cancer Therapy Strategies. Coord. Chem. Rev. 2019, 387, 299–324. [Google Scholar] [CrossRef]
- Vines, J.B.; Yoon, J.H.; Ryu, N.E.; Lim, D.J.; Park, H. Gold Nanoparticles for Photothermal Cancer Therapy. Front. Chem. 2019, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, L.; Zheng, W.; Cong, L.; Guo, Z.; Xie, Y.; Wang, L.; Tang, R.; Feng, Q.; Hamada, Y.; et al. Thermo-Triggered Release of CRISPR-Cas9 System by Lipid-Encapsulated Gold Nanoparticles for Tumor Therapy. Angew. Chemie-Int. Ed. 2018, 57, 1491–1496. [Google Scholar] [CrossRef] [PubMed]
- Hervault, A.; Thanh, N.T.K. Magnetic Nanoparticle-Based Therapeutic Agents for Thermo-Chemotherapy Treatment of Cancer. Nanoscale 2014, 6, 11553–11573. [Google Scholar] [CrossRef]
- Wu, M.; Huang, S. Magnetic Nanoparticles in Cancer Diagnosis, Drug Delivery and Treatment. Mol. Clin. Oncol. 2017, 738–746. [Google Scholar] [CrossRef]
- Rohiwal, S.S.; Dvorakova, N.; Klima, J.; Vaskovicova, M.; Senigl, F.; Slouf, M.; Pavlova, E.; Stepanek, P.; Babuka, D.; Benes, H.; et al. Polyethylenimine Based Magnetic Nanoparticles Mediated Non-Viral CRISPR/Cas9 System for Genome Editing. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Hu, C.-M.; Zhang, L. Therapeutic Nanoparticles to Combat Cancer Drug Resistance. Curr. Drug Metab. 2010, 10, 836–841. [Google Scholar] [CrossRef]
- Huwyler, J.; Cerletti, A.; Fricker, G.; Eberle, A.N.; Drewe, J. By-Passing of P-Glycoprotein Using Immunoliposomes. J. Drug Target. 2002, 10, 73–79. [Google Scholar] [CrossRef]
- Kumar, S.; Diwan, A.; Singh, P.; Gulati, S.; Choudhary, D.; Mongia, A.; Shukla, S.; Gupta, A. Functionalized Gold Nanostructures: Promising Gene Delivery Vehicles in Cancer Treatment. RSC Adv. 2019, 9, 23894–23907. [Google Scholar] [CrossRef]
- Kaestner, L.; Scholz, A.; Lipp, P. Conceptual and Technical Aspects of Transfection and Gene Delivery. Bioorganic Med. Chem. Lett. 2015, 25, 1171–1176. [Google Scholar] [CrossRef]
- Karimi, S.; Fouani, M.H.; Moshaii, A.; Nikkhah, M.; Hosseinkhani, S.; Sheikhnejad, R. Development of Dual Functional Nucleic Acid Delivery Nanosystem for DNA Induced Silencing of BCL-2 Oncogene. Int. J. Nanomed. 2020, 15, 1693–1708. [Google Scholar] [CrossRef] [PubMed]
- Bessis, N.; GarciaCozar, F.J.; Boissier, M.C. Immune Responses to Gene Therapy Vectors: Influence on Vector Function and Effector Mechanisms. Gene Ther. 2004, 11, S10–S17. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorth, M.; Narvekar, A. Non Viral Vectors in Gene Therapy- An Overview. J. Clin. DIAGNOSTIC Res. 2015, 9, GE01. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Stewart, D.J.; Lee, J.J.; Ji, L.; Ramesh, R.; Jayachandran, G.; Nunez, M.I.; Wistuba, I.I.; Erasmus, J.J.; Hicks, M.E.; et al. Phase I Clinical Trial of Systemically Administered TUSC2(FUS1)-Nanoparticles Mediating Functional Gene Transfer in Humans. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Schultheis, B.; Strumberg, D.; Santel, A.; Vank, C.; Gebhardt, F.; Keil, O.; Lange, C.; Giese, K.; Kaufmann, J.; Khan, M.; et al. First-in-Human Phase I Study of the Liposomal RNA Interference Therapeutic Atu027 in Patients with Advanced Solid Tumors. J. Clin. Oncol. 2014, 32, 4141–4148. [Google Scholar] [CrossRef]
- Plank, C.; Schillinger, U.; Scherer, F.; Bergemann, C.; Rémy, J.S.; Krötz, F.; Anton, M.; Lausier, J.; Rosenecker, J. The Magnetofection Method: Using Magnetic Force to Enhance Gene Delivery. Biol. Chem. 2003, 384, 737–747. [Google Scholar] [CrossRef]
- Amendoeira, A.; García, L.R.; Fernandes, A.R.; Baptista, P.V. Light Irradiation of Gold Nanoparticles Toward Advanced Cancer Therapeutics. Adv. Ther. 2020, 3, 1900153. [Google Scholar] [CrossRef]
- Chang, D.; Lim, M.; Goos, J.A.C.M.; Qiao, R.; Ng, Y.Y.; Mansfeld, F.M.; Jackson, M.; Davis, T.P.; Kavallaris, M. Biologically Targeted Magnetic Hyperthermia: Potential and Limitations. Front. Pharmacol. 2018, 9, 831. [Google Scholar] [CrossRef]
- Dulińska-Litewka, J.; Łazarczyk, A.; Hałubiec, P.; Szafrański, O.; Karnas, K.; Karewicz, A. Superparamagnetic Iron Oxide Nanoparticles-Current and Prospective Medical Applications. Materials 2019, 12, 617. [Google Scholar] [CrossRef]
- Beola, L.; Asín, L.; Fratila, R.M.; Herrero, V.; De La Fuente, J.M.; Grazú, V.; Gutiérrez, L. Dual Role of Magnetic Nanoparticles as Intracellular Hotspots and Extracellular Matrix Disruptors Triggered by Magnetic Hyperthermia in 3D Cell Culture Models. ACS Appl. Mater. Interfaces 2018, 10, 44301–44313. [Google Scholar] [CrossRef]
- Cazares-Cortes, E.; Cabana, S.; Boitard, C.; Nehlig, E.; Griffete, N.; Fresnais, J.; Wilhelm, C.; Abou-Hassan, A.; Ménager, C. Recent Insights in Magnetic Hyperthermia: From the “Hot-Spot” Effect for Local Delivery to Combined Magneto-Photo-Thermia Using Magneto-Plasmonic Hybrids. Adv. Drug Deliv. Rev. 2019, 138, 233–246. [Google Scholar] [CrossRef]
- Mello, P.d.A.; Bian, S.; Savio, L.E.B.; Zhang, H.; Zhang, J.; Junger, W.; Wink, M.R.; Lenz, G.; Buffon, A.; Wu, Y.; et al. Hyperthermia and Associated Changes in Membrane Fluidity Potentiate P2X7 Activation to Promote Tumor Cell Death. Oncotarget 2017, 8, 67254. [Google Scholar] [CrossRef]
- Garanina, A.S.; Naumenko, V.A.; Nikitin, A.A.; Myrovali, E.; Petukhova, A.Y.; Klimyuk, S.V.; Nalench, Y.A.; Ilyasov, A.R.; Vodopyanov, S.S.; Erofeev, A.S.; et al. Temperature-Controlled Magnetic Nanoparticles Hyperthermia Inhibits Primary Tumor Growth and Metastases Dissemination. Nanomed. Nanotechnol. Biol. Med. 2020, 25, 102171. [Google Scholar] [CrossRef]
- Conde, J.; Rosa, J.; de la Fuente, J.M.; Baptista, P.V. Gold-Nanobeacons for Simultaneous Gene Specific Silencing and Intracellular Tracking of the Silencing Events. Biomaterials 2013, 34, 2516–2523. [Google Scholar] [CrossRef]
- Pedrosa, P.; Corvo, M.L.; Ferreira-Silva, M.; Martins, P.; Carvalheiro, M.C.; Costa, P.M.; Martins, C.; Martins, L.M.D.R.S.; Baptista, P.V.; Fernandes, A.R. Targeting Cancer Resistance via Multifunctional Gold Nanoparticles. Int. J. Mol. Sci. 2019, 20, 5510. [Google Scholar] [CrossRef]
- Foroozandeh, P.; Aziz, A.A. Insight into Cellular Uptake and Intracellular Trafficking of Nanoparticles. Nanoscale Res. Lett. 2018, 13, 1–12. [Google Scholar] [CrossRef]
- Ding, Y.; Jiang, Z.; Saha, K.; Kim, C.S.; Kim, S.T.; Landis, R.F.; Rotello, V.M. Gold Nanoparticles for Nucleic Acid Delivery. Mol. Ther. 2014, 22, 1075–1083. [Google Scholar] [CrossRef]
- Roma-Rodrigues, C.; Mendes, R.; Baptista, P.V.; Fernandes, A.R. Targeting Tumor Microenvironment for Cancer Therapy. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Barnaby, S.N.; Lee, A.; Mirkin, C.A. Probing the Inherent Stability of SiRNA Immobilized on Nanoparticle Constructs. Proc. Natl. Acad. Sci. USA 2014, 111, 9739–9744. [Google Scholar] [CrossRef]
- Palchetti, S.; Pozzi, D.; Capriotti, A.L.; Barbera, G.L.; Chiozzi, R.Z.; Digiacomo, L.; Peruzzi, G.; Caracciolo, G.; Laganà, A. Influence of Dynamic Flow Environment on Nanoparticle-Protein Corona: From Protein Patterns to Uptake in Cancer Cells. Coll. Surf. B Biointerfaces 2017, 153, 263–271. [Google Scholar] [CrossRef]
- Falahati, M.; Attar, F.; Sharifi, M.; Haertlé, T.; Berret, J.F.; Khan, R.H.; Saboury, A.A. A Health Concern Regarding the Protein Corona, Aggregation and Disaggregation. Biochim. Biophys. Acta -Gen. Subj. 2019, 1863, 971–991. [Google Scholar] [CrossRef]
- Moore, T.L.; Rodriguez-Lorenzo, L.; Hirsch, V.; Balog, S.; Urban, D.; Jud, C.; Rothen-Rutishauser, B.; Lattuada, M.; Petri-Fink, A. Nanoparticle Colloidal Stability in Cell Culture Media and Impact on Cellular Interactions. Chem. Soc. Rev. 2015, 44, 6287–6305. [Google Scholar] [CrossRef]
- Zhang, P.; An, K.; Duan, X.; Xu, H.; Li, F.; Xu, F. Recent Advances in SiRNA Delivery for Cancer Therapy Using Smart Nanocarriers. Drug Discov. Today 2018, 23, 900–911. [Google Scholar] [CrossRef]
- Huang, H.C.; Barua, S.; Kay, D.B.; Rege, K. Simultaneous Enhancement of Photothermal Stability and Gene Delivery Efficacy of Gold Nanorods Using Polyelectrolytes. ACS Nano 2009, 3, 2941–2952. [Google Scholar] [CrossRef]
- Wang, J.; Thomas, M.; Lin, P.; Cheng, J.X.; Matei, D.E.; Wei, A. SiRNA Delivery Using Dithiocarbamate-Anchored Oligonucleotides on Gold Nanorods. Bioconjug. Chem. 2019, 30, 443–453. [Google Scholar] [CrossRef]
- Jo, H.; Ban, C. Aptamer-Nanoparticle Complexes as Powerful Diagnostic and Therapeutic Tools. Exp. Mol. Med. 2016, 48. [Google Scholar] [CrossRef]
- Fukushima, H.; Yamashita, S.; Mori, T.; Katayama, Y.; Niidome, T. Sequential Release of Single-Stranded Dnas from Gold Nanorods Triggered by near-Infrared Light Irradiation. Chem. Lett. 2012, 41, 711–712. [Google Scholar] [CrossRef]
- Mahajan, S.D.; Aalinkeel, R.; Law, W.C.; Reynolds, J.L.; Nair, B.B.; Sykes, D.E.; Yong, K.T.; Roy, I.; Prasad, P.N.; Schwartz, S.A. Anti-HIV-1 Nanotherapeutics: Promises and Challenges for the Future. Int. J. Nanomed. 2012, 7, 5301–5314. [Google Scholar] [CrossRef]
- Silva, J.; Fernandes, A.R.; Baptista, P.V. Application of Nanotechnology in Drug Delivery. In Application of Nanotechnology in Drug Delivery; InTech: London, UK, 2014. [Google Scholar] [CrossRef]
- Leung, J.; Wu, S.; Chou, K.; Signorell, R. Investigation of Sub-100 Nm Gold Nanoparticles for Laser-Induced Thermotherapy of Cancer. Nanomaterials 2013, 3, 86–106. [Google Scholar] [CrossRef]
- Arnida; Janát-Amsbury, M.M.; Ray, A.; Peterson, C.M.; Ghandehari, H. Geometry and Surface Characteristics of Gold Nanoparticles Influence Their Biodistribution and Uptake by Macrophages. Eur. J. Pharm. Biopharm. 2011, 77, 417–423. [Google Scholar] [CrossRef]
- Xia, Y.; Li, W.; Cobley, C.M.; Chen, J.; Xia, X.; Zhang, Q.; Yang, M.; Cho, E.C.; Brown, P.K. Gold Nanocages: From Synthesis to Theranostic Applications. Acc. Chem. Res. 2011, 44, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wang, Z.; Bi, L.; Bi, C.; Du, Q. Gold Nanocage-Based Surface-Enhanced Raman Scattering Probes for Long-Term Monitoring of Intracellular MicroRNA during Bone Marrow Stem Cell Differentiation. Nanoscale 2020, 12, 1513–1527. [Google Scholar] [CrossRef] [PubMed]
- Robinson, R.; Gerlach, W.; Ghandehari, H. Comparative Effect of Gold Nanorods and Nanocages for Prostate Tumor Hyperthermia. J. Control. Release 2015, 220, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Singhana, B.; Slattery, P.; Chen, A.; Wallace, M.; Melancon, M.P. Light-Activatable Gold Nanoshells for Drug Delivery Applications. AAPS PharmSciTech 2014, 15, 741–752. [Google Scholar] [CrossRef]
- Wang, Z.; Li, S.; Zhang, M.; Ma, Y.; Liu, Y.; Gao, W.; Zhang, J.; Gu, Y. Laser-Triggered Small Interfering RNA Releasing Gold Nanoshells against Heat Shock Protein for Sensitized Photothermal Therapy. Adv. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Huschka, R.; Barhoumi, A.; Liu, Q.; Roth, J.A.; Ji, L.; Halas, N.J. Gene Silencing by Gold Nanoshell-Mediated Delivery and Laser-Triggered Release of Antisense Oligonucleotide and SiRNA. ACS Nano 2012, 6, 7681–7691. [Google Scholar] [CrossRef]
- Braun, G.B.; Pallaoro, A.; Wu, G.; Missirlis, D.; Zasadzinski, J.A.; Tirrell, M.; Reich, N.O. Laser-Activated Gene Silencing via Gold Nanoshell-SiRNA Conjugates. ACS Nano 2009, 3, 2007–2015. [Google Scholar] [CrossRef]
- Pannerec-Varna, M.; Ratajczak, P.; Bousquet, G.; Ferreira, I.; Leboeuf, C.; Boisgard, R.; Gapihan, G.; Verine, J.; Palpant, B.; Bossy, E.; et al. In Vivo Uptake and Cellular Distribution of Gold Nanoshells in a Preclinical Model of Xenografted Human Renal Cancer. Gold Bull. 2013, 46, 257–265. [Google Scholar] [CrossRef]
- Yetisgin, A.A.; Cetinel, S.; Zuvin, M.; Kosar, A.; Kutlu, O. Therapeutic Nanoparticles and Their Targeted Delivery Applications. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Grijalvo, S.; Alagia, A.; Jorge, A.F.; Eritja, R. Covalent Strategies for Targeting Messenger and Non-Coding RNAs: An Updated Review on SiRNA, MiRNA and AntimiR Conjugates. Genes (Basel). 2018, 9. [Google Scholar] [CrossRef]
- Rosi, N.L.; Giljohann, D.A.; Thaxton, C.S.; Lytton-Jean, A.K.R.; Han, M.S.; Mirkin, C.A. Oligonucleotide-Modified Gold Nanoparticles for Infracellular Gene Regulation. Science 2006, 312, 1027–1030. [Google Scholar] [CrossRef] [PubMed]
- Remant Bahadur, K.C.; Thapa, B.; Bhattarai, N. Gold Nanoparticle-Based Gene Delivery: Promises and Challenges. Nanotechnol. Rev. 2014, 3, 269–280. [Google Scholar] [CrossRef]
- Thomas, M.; Klibanov, A.M. Conjugation to Gold Nanoparticles Enhances Polyethylenimine’s Transfer of Plasmid Dna into Mammalian Cells. Proc. Natl. Acad. Sci. USA 2003, 100, 9138–9143. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Peng, Q.; Chen, F.; Zhong, Z.; Zhuo, R. Low Molecular Weight Polyethylenimine Conjugated Gold Nanoparticles as Efficient Gene Vectors. Bioconjug. Chem. 2010, 21, 836–843. [Google Scholar] [CrossRef]
- Kong, W.H.; Bae, K.H.; Jo, S.D.; Kim, J.S.; Park, T.G. Cationic Lipid-Coated Gold Nanoparticles as Efficient and Non-Cytotoxic Intracellular SiRNA Delivery Vehicles. Pharm. Res. 2012, 29, 362–374. [Google Scholar] [CrossRef]
- Li, P.; Li, D.; Zhang, L.; Li, G.; Wang, E. Cationic Lipid Bilayer Coated Gold Nanoparticles-Mediated Transfection of Mammalian Cells. Biomaterials 2008, 29, 3617–3624. [Google Scholar] [CrossRef]
- Kawano, T.; Yamagata, M.; Takahashi, H.; Niidome, Y.; Yamada, S.; Katayama, Y.; Niidome, T. Stabilizing of Plasmid DNA in Vivo by PEG-Modified Cationic Gold Nanoparticles and the Gene Expression Assisted with Electrical Pulses. J. Control. Release 2006, 111, 382–389. [Google Scholar] [CrossRef]
- Lee, S.H.; Bae, K.H.; Kim, S.H.; Lee, K.R.; Park, T.G. Amine-Functionalized Gold Nanoparticles as Non-Cytotoxic and Efficient Intracellular SiRNA Delivery Carriers. Int. J. Pharm. 2008, 364, 94–101. [Google Scholar] [CrossRef]
- Baptista, P.V. Gold Nanobeacons: A Potential Nanotheranostics Platform. Nanomedicine 2014, 9, 2247–2250. [Google Scholar] [CrossRef]
- Mirkin, C.A.; Letsinger, R.L.; Mucic, R.C.; Storhoff, J.J. A DNA-Based Method for Rationally Assembling Nanoparticles into Macroscopic Materials. Nature 1996, 382, 607–609. [Google Scholar] [CrossRef]
- Carnerero, J.M.; Jimenez-Ruiz, A.; Castillo, P.M.; Prado-Gotor, R. Covalent and Non-Covalent DNA–Gold-Nanoparticle Interactions: New Avenues of Research. ChemPhysChem 2017, 18, 17–33. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, C.M.; Esposito, E.A.; Boal, A.K.; Simard, J.M.; Martin, C.T.; Rotello, V.M. Inhibition of DNA Transcription Using Cationic Mixed Monolayer Protected Gold Clusters. J. Am. Chem. Soc. 2001, 123, 7626–7629. [Google Scholar] [CrossRef]
- Ghosh, P.S.; Kim, C.K.; Han, G.; Forbes, N.S.; Rotello, V.M. Efficient Gene Delivery Vectors by Tuning the Surface Charge Density of Amino Acid-Functionalized Gold Nanoparticles. ACS Nano 2008, 2, 2213–2218. [Google Scholar] [CrossRef] [PubMed]
- Elbakry, A.; Zaky, A.; Liebl, R.; Rachel, R.; Goepferich, A.; Breunig, M. Layer-by-Layer Assembled Gold Nanoparticles for Sirna Delivery. Nano Lett. 2009, 9, 2059–2064. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Huang, Y.; Jiang, Q.; Sun, Y.; Deng, L.; Liang, Z.; Du, Q.; Xing, J.; Zhao, Y.; Wang, P.C.; et al. Enhanced Gene Delivery and SiRNA Silencing by Gold Nanoparticles Coated with Charge-Reversal Polyelectrolyte. ACS Nano 2010, 4, 5505–5511. [Google Scholar] [CrossRef]
- Hong, R.; Han, G.; Fernández, J.M.; Kim, B.J.; Forbes, N.S.; Rotello, V.M. Glutathione-Mediated Delivery and Release Using Monolayer Protected Nanoparticle Carriers. J. Am. Chem. Soc. 2006, 128, 1078–1079. [Google Scholar] [CrossRef]
- Han, G.; You, C.C.; Kim, B.J.; Turingan, R.S.; Forbes, N.S.; Martin, C.T.; Rotello, V.M. Light-Regulated Release of DNA and Its Delivery to Nuclei by Means of Photolabile Gold Nanoparticles. Angew. Chem.-Int. Ed. 2006, 45, 3165–3169. [Google Scholar] [CrossRef]
- Yuan, C.; Tian, T.; Sun, J.; Hu, M.; Wang, X.; Xiong, E.; Cheng, M.; Bao, Y.; Lin, W.; Jiang, J.; et al. Universal and Naked-Eye Gene Detection Platform Based on the Clustered Regularly Interspaced Short Palindromic Repeats/Cas12a/13a System. Anal. Chem. 2020, 92, 4029–4037. [Google Scholar] [CrossRef]
- Shahbazi, R.; Sghia-Hughes, G.; Reid, J.L.; Kubek, S.; Haworth, K.G.; Humbert, O.; Kiem, H.P.; Adair, J.E. Targeted Homology-Directed Repair in Blood Stem and Progenitor Cells with CRISPR Nanoformulations. Nat. Mater. 2019, 18, 1124–1132. [Google Scholar] [CrossRef]
- Ryu, Y.; Kang, J.A.; Kim, D.; Kim, S.R.; Kim, S.; Park, S.J.; Kwon, S.H.; Kim, K.N.; Lee, D.E.; Lee, J.; et al. Programed Assembly of Nucleoprotein Nanoparticles Using DNA and Zinc Fingers for Targeted Protein Delivery. Small 2018, 14, 1–13. [Google Scholar] [CrossRef]
- Petree, J.R.; Yehl, K.; Galior, K.; Glazier, R.; Deal, B.; Salaita, K. Site-Selective RNA Splicing Nanozyme: DNAzyme and RtcB Conjugates on a Gold Nanoparticle. ACS Chem. Biol. 2018, 13, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Tian, F.; Hernández, Y.; Bao, C.; Cui, D.; Janssen, K.P.; Ibarra, M.R.; Baptista, P.V.; Stoeger, T.; de la Fuente, J.M. In Vivo Tumor Targeting via Nanoparticle-Mediated Therapeutic SiRNA Coupled to Inflammatory Response in Lung Cancer Mouse Models. Biomaterials 2013, 34, 7744–7753. [Google Scholar] [CrossRef] [PubMed]
- Vinhas, R.; Fernandes, A.R.; Baptista, P.V. Gold Nanoparticles for BCR-ABL1 Gene Silencing: Improving Tyrosine Kinase Inhibitor Efficacy in Chronic Myeloid Leukemia. Mol. Ther.-Nucleic Acids 2017, 7, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Shen, N.; Yang, Y.; Yu, H.; Xu, S.; Yang, Y.W.; Liu, S.; Meguellati, K.; Yan, F. Targeting Epigenetic Pathway with Gold Nanoparticles for Acute Myeloid Leukemia Therapy. Biomaterials 2018, 167, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Rahme, K.; Guo, J.; Holmes, J.D. Bioconjugated Gold Nanoparticles Enhance SiRNA Delivery in Prostate Cancer Cells. Methods Mol. Biol. 2019, 1974, 291–301. [Google Scholar] [CrossRef]
- Ryou, S.M.; Park, M.; Kim, J.M.; Jeon, C.O.; Yun, C.H.; Han, S.H.; Kim, S.W.; Lee, Y.; Kim, S.; Han, M.S.; et al. Inhibition of Xenograft Tumor Growth in Mice by Gold Nanoparticle-Assisted Delivery of Short Hairpin RNAs against Mcl-1L. J. Biotechnol. 2011, 156, 89–94. [Google Scholar] [CrossRef]
- Ryou, S.M.; Kim, S.; Jang, H.H.; Kim, J.H.; Yeom, J.H.; Eom, M.S.; Bae, J.; Han, M.S.; Lee, K. Delivery of ShRNA Using Gold Nanoparticle-DNA Oligonucleotide Conjugates as a Universal Carrier. Biochem. Biophys. Res. Commun. 2010, 398, 542–546. [Google Scholar] [CrossRef]
- Pedrosa, P.; Heuer-Jungemann, A.; Kanaras, A.G.; Fernandes, A.R.; Baptista, P.V. Potentiating Angiogenesis Arrest in Vivo via Laser Irradiation of Peptide Functionalised Gold Nanoparticles. J. Nanobiotechnol. 2017, 15, 85. [Google Scholar] [CrossRef]
- Son, S.; Kim, N.; You, D.G.; Yoon, H.Y.; Yhee, J.Y.; Kim, K.; Kwon, I.C.; Kim, S.H. Antitumor Therapeutic Application of Self-Assembled RNAi-AuNP Nanoconstructs: Combination of VEGF-RNAi and Photothermal Ablation. Theranostics 2017, 7, 9–22. [Google Scholar] [CrossRef]
- Roma-Rodrigues, C.; Pereira, F.; Alves de Matos, A.P.; Fernandes, M.; Baptista, P.V.; Fernandes, A.R. Smuggling Gold Nanoparticles across Cell Types-A New Role for Exosomes in Gene Silencing. Nanomedicine 2017, 13, 1389–1398. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic: An Update. Bioeng. Transl. Med. 2019, 4, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic. Bioeng. Transl. Med. 2016, 1, 10–29. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Huang, Q.; Li, C.; Yang, Y.; Wang, X.; Yu, J.; Huang, Y.; Liang, X.J. Improved Nucleic Acid Therapy with Advanced Nanoscale Biotechnology. Mol. Ther. -Nucleic Acids 2020, 19, 581–601. [Google Scholar] [CrossRef] [PubMed]
- Conde, J.; Ambrosone, A.; Sanz, V.; Hernandez, Y.; Marchesano, V.; Tian, F.; Child, H.; Berry, C.C.; Ibarra, M.R.; Baptista, P.V.; et al. Design of Multifunctional Gold Nanoparticles for in Vitro and in Vivo Gene Silencing. ACS Nano 2012, 6, 8316–8324. [Google Scholar] [CrossRef]
- Hirakawa, M.P.; Krishnakumar, R.; Timlin, J.A.; Carney, J.P.; Butler, K.S. Gene Editing and CRISPR in the Clinic: Current and Future Perspectives. Biosci. Rep. 2020, 40. [Google Scholar] [CrossRef]
- Clinical Trials. gov. Available online: https://clinicaltrials.gov/ (accessed on 2 June 2020).
- Hua, S.; de Matos, M.B.C.; Metselaar, J.M.; Storm, G. Current Trends and Challenges in the Clinical Translation of Nanoparticulate Nanomedicines: Pathways for Translational Development and Commercialization. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Baptista, P.; Fernandes, A.; Figueiredo, S.; Vinhas, R.; Cordeiro, M.; Carlos, F.; Mendo, S. Gold Nanoparticle-Based Theranostics: Disease Diagnostics and Treatment Using a Single Nanomaterial. Nanobiosens. Dis. Diagn. 2015, 4, 11. [Google Scholar] [CrossRef]
- Ioannidis, J.P.A.; Kim, B.Y.S.; Trounson, A. How to Design Preclinical Studies in Nanomedicine and Cell Therapy to Maximize the Prospects of Clinical Translation. Nat. Biomed. Eng. 2018, 2, 797–809. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Platform | Condition/Disease | Target | Cells | Vectorization | Clinical Trial Reference (Phase) |
|---|---|---|---|---|---|
| ZFN | HIV-1 infection | CCR5 | T (CD4+) | Adenovirus | NCT01252641 (I/II) |
| T CD4/CD8 | mRNA | NCT02225665 (I/II) | |||
| HSPC (CD34+) | mRNA | NCT02500849 (I) | |||
| HPV-induced precancerous Lesions of cervix | HPV16/18 E7 | Epithelial | DNA | NCT02800369 (I) | |
| Hemophilia B | Factor IX gene | Hepatocytes | AAV | NCT02695160 (I) | |
| TALEN | Relapsed/refractory B-ALL | CD52, TRAC | CAR T | Lentivirus | NCT02808442 (I) |
| HPV-related cervical cancer | HPV16/18 E6/E7 | Epithelial | Plasmid | NCT03226470 (I) | |
| TALENs coupled to CRISPR/Cas9 | HPV-associated cervical cancer | HPV16/18 E6/E7 | Epithelial | Plasmid | NCT03057912 (I) |
| CRISPR/Cas9 | AML | CD123, TRAC | CAR T | mRNA | NCT03190278 (I) |
| Various cancers | PDCD1 | T | DNA | NCT02793856 (I) NCT03081715 (I) | |
| Relapsed/refractory CD19+ leukemia and lymphoma | TRAC, B2M | CAR T | Lentivirus, electroporation | NCT03166878 (I/II) | |
| CD19 and CD20 or CD22, TRAC | CAR T | Lentivirus, electroporation | NCT03398967 (I/II) | ||
| T cell leukemia, lymphoma | CD7, CD28 | CAR T | - | NCT03690011 (I) | |
| β-thalassemia | BCL11A | HSPC (CD34+) | - | NCT03655678 (I/II) | |
| Sickle cell disease | BCL11A | HSPC (CD34+) | - | NCT03745287 (I/II) |
| Gene Therapy Tools | Characteristics | Advantages | Limitations | Application in Cancer Therapy | Ref. |
|---|---|---|---|---|---|
| CRISPR/Cas9 | CRISPR/Cas9 facilitate efficient multiplex genome editing, with the possibility of simultaneous deletion or insertion of multiple DNA sequences. | CRISPR/Cas9 is fast to develop, simple and cost-saving in comparison with other gene editing tools. Potential for simultaneous multiple loci editing. | High frequency of random integration. Activation of non-homologous end joining DNA repair pathway (may lead to microdeletions at the DSB site). | T-Cell modification in cancer therapy. | [15,16,40,41,42] |
| ZFNs | DNA-binding domains recognize trinucleotide DNA sequences (longer DNA sequences may also be targeted). | ZFN may be designed to include a variety of effector domains to recognize virtually any DNA sequence. | Off-targets effects are high. Expensive and hard to use technique. | Clinical application other than cancer (HIV). | [21,43] |
| TALENs | DNA targeting specificity comes from the fused bacterial TALE proteins. TALEN arrays recognize only a single nucleotide (as opposed to ZFNs) and it has no impact on the binding specificity. | TALEN engineered nucleases show better specificity and efficiency than ZFN. | The major limitation is the cloning of the large modules in series and join these modules in designated order by ligase in an efficient way. | Clinical trials -Cervical intraepithelial neoplasia; hematological malignancies. | [9,40,43] |
| RNA interference | RNAi generally used to down-regulate gene expression. Also, the effector molecules (e.g., siRNA) may be vectorized alone or expressed from suitable plasmids. | RNAi uses the cells’ machinery, facilitated by short interfering RNA molecules. | Short half-lives due to RNases. Low biochemical stability. High costs. | Silencing oncogenes in several cancer–clinical use. | [31,44,45,46] |
| Antisense | AON are small DNA or RNA molecules complementary to their target mRNA. Binding to their target result in alteration of mRNA splicing or degradation of target transcripts. | Simple to design and vectorize inside the cell | Obstacles for in vivo delivery (immunogenic) that may cause side effects. Short half-lives due to RNase activity. | Clinical application other than cancer (e.g., thalassemia). | [29,31,47,48] |
| Meganucleases | Meganuclease technology involves re-engineering the DNA-binding specificity that naturally occurs in the family of homing endonucleases. | Meganucleases are the smallest class of engineered nucleases, making them potentially amenable to all standard gene delivery methods since they offer fewer off-targets effects. | Meganucleases are difficult to construct, time-consuming and high costing limiting their use as gene editing tools. | No reports on clinical applications. | [27,43] |
| Type of NPs | Characteristics | Application | Limitations | Examples | Gene Editing Tool | Ref. |
|---|---|---|---|---|---|---|
| Dendrimer | Small (1–15 nm) branched polymeric NPs High water solubility, high cargo capacity. | Targeting cancer cells and injured tissues | Synthesis is quite time-consuming; Some toxicity issues in drug incorporation and release. | PAMAM dendrimers used as drug delivery systems. | Boronic acid-rich dendrimer as vector for CRISPR/Cas9. Meganucleases | [53,54,55,56] |
| Liposome | Nanostructures made of amphiphilic molecules (polymers, lipids) with good biocompatibility and cell uptake. | Delivery of hydrophilic or hydrophobic cargo depending on lipid constitution. | Poor stability, rapid degradation and clearance. May trigger lipid oxidation (long-term). | Doxorubicin in a heat sensitive liposomal formulation-ThermoDox® (Celsion) in phase III clinical trial in primary hepatocellular carcinoma; in phase II for refractory breast cancer and colorectal liver metastasis. | Lipid delivery systems for siRNA delivery. Meganucleases | [56,57,58,59] |
| Polymeric NP | Natural (proteins and polypeptides) or synthetic. Biocompatibility and biodegradable. | Controlled drug release, protection of drug molecules specific targeting. | Low transfection efficiency Some cytotoxicity. | Most used is PEI (Polyethyleneimine) in nanosized ionic complexes (polyplexes). | Delivery of plasmid DNA (CRISPR–Cas9) in PLGA. | [60,61,62] |
| Carbon NP | Carbon dots, graphene, oxides, and carbon nanotubes (CNT). Unique mechanical and optical properties. | Imaging and drug delivery applications. | Cytotoxicity. | Stimuli responsive drug delivery systems. | siRNA delivery and intracellular tracking of siRNA (nanotheranostics). | [63,64,65] |
| Quantum Dots | QDs are luminescent nanoprobes that present high photostability, i.e., no photobleaching. | Used in imaging, detection and targeting. | High toxicity due to the CdSe (metallic core of the NP). | Quantum Dots can act as photosensitizers, producing reactive oxygen species (ROS) upon light irradiation. | No reports on clinical applications. | [66,67] |
| Gold NP (AuNP) | Metallic core NPs with unique optical and physical-chemical properties. | Primarily used for labelling applications; may be used in theranostic tools. | Cytotoxicity, biodistribution, retention time, and physiological response of NPs. | AuNP can be used as photothermal agents in hyperthermia, and nanocarriers for gene silencing. | Deliver of CRISPR-Cas9 with/without external stimuli | [39,68,69,70] |
| Magnetic NP (MNP) | Actuated by an external magnetic field. | Imaging. | Cytotoxicity of ion core. | Ferucarbotran (Resovist®)-clinically approved superparamagnetic iron oxide nanoparticles (SPIONs) coated with carboxydextran for the enhancement of MRI contrast of the liver. | CRISPR/Cas9-PEI-MNP. | [71,72,73] |
| AuNPs | Characteristics | Advantages | Limitations | Applications | TNAs | In Vivo Distribution | Ref. |
|---|---|---|---|---|---|---|---|
| AuNRs | Elongated NPs, showing longitudinal plasmon wavelength with nearly linear dependence on their aspect ratio. | Tunable properties, including SPR. | Low drug loading capacity. Poor control over size distribution. | Tunable optical resonance in the NIR for in vivo applications, such as imaging, photothermy. | siRNA DNA aptamers ASOs RNA decoys | Accumulation in the liver, long circulation time, and high accumulation in the tumors. | [101,102,103,104,105,106,107,108] |
| AuNCus | Hollow with ultrathin and porous walls. Easy to synthesize (scale up). | Tunable sizes and scalability. | Few data about toxicity, biodistribution, and physiological response. | Theranostics (SPR can be tuned between 600–1200 nm); hollow interiors allow encapsulation; porous walls for ease drug release. | siRNAs miRNAs | Medium level accumulation in the liver, kidneys and spleen. Rapidly excreted. | [102,106,109,110,111] |
| AuNShs | Spherical with a dielectric core covered by a thin gold shell. | Improve in vivo bioavailability and controlled drug release. | Lack of targeting efficacy. Limited tracking/monitoring in vivo. | Controlled/triggered drug delivery via irradiation (NIR). | siRNA ASOs | Short circulation times, accumulation in liver and spleen. No induction of tissue damage (necrosis, inflammatory infiltrate or fibrosis) liver, spleen, kidney or bone marrow. | [112,113,114,115,116] |
| AuNSs | Spherical solid. | Ease of functionalization. Enhanced cellular uptake. | Prone to aggregation. Multitude of possible cell uptake routes render difficult to control. | Extremely versatile for photothermy, and combined therapy. | ASOs siRNAs miRNA | Short circulation time and accumulation in the liver with low accumulation in the tumors. | [91,117,118,119] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, D.; Fontinha, D.; Martins, C.; Pires, D.; Fernandes, A.R.; Baptista, P.V. Gold Nanoparticles for Vectorization of Nucleic Acids for Cancer Therapeutics. Molecules 2020, 25, 3489. https://doi.org/10.3390/molecules25153489
Ferreira D, Fontinha D, Martins C, Pires D, Fernandes AR, Baptista PV. Gold Nanoparticles for Vectorization of Nucleic Acids for Cancer Therapeutics. Molecules. 2020; 25(15):3489. https://doi.org/10.3390/molecules25153489
Chicago/Turabian StyleFerreira, Daniela, David Fontinha, Catarina Martins, David Pires, Alexandra R. Fernandes, and Pedro V. Baptista. 2020. "Gold Nanoparticles for Vectorization of Nucleic Acids for Cancer Therapeutics" Molecules 25, no. 15: 3489. https://doi.org/10.3390/molecules25153489
APA StyleFerreira, D., Fontinha, D., Martins, C., Pires, D., Fernandes, A. R., & Baptista, P. V. (2020). Gold Nanoparticles for Vectorization of Nucleic Acids for Cancer Therapeutics. Molecules, 25(15), 3489. https://doi.org/10.3390/molecules25153489