Clove Oil (Syzygium aromaticum L.) Activity against Alicyclobacillus acidoterrestris Biofilm on Technical Surfaces

 , , and
, , and
Abstract
1. Introduction
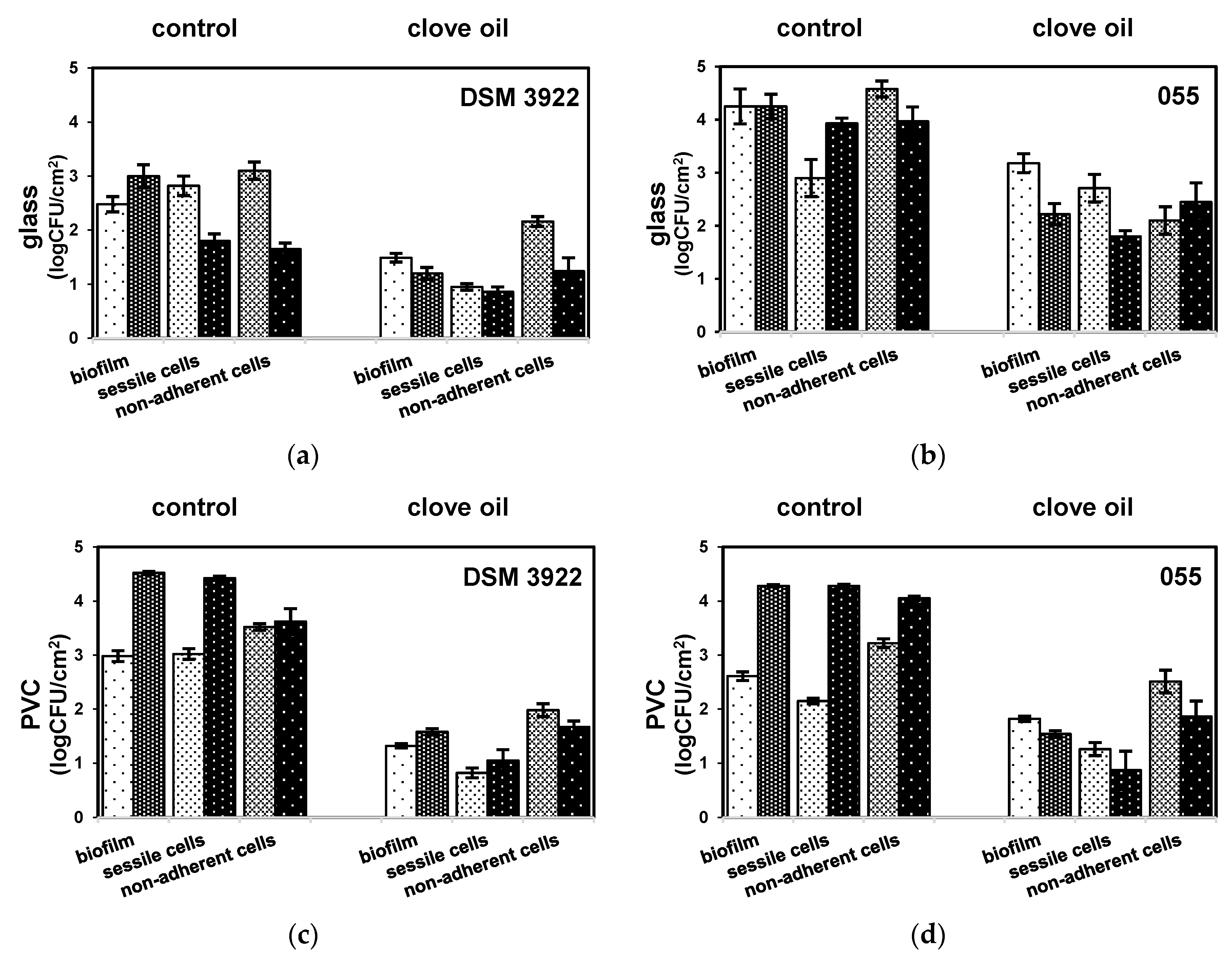
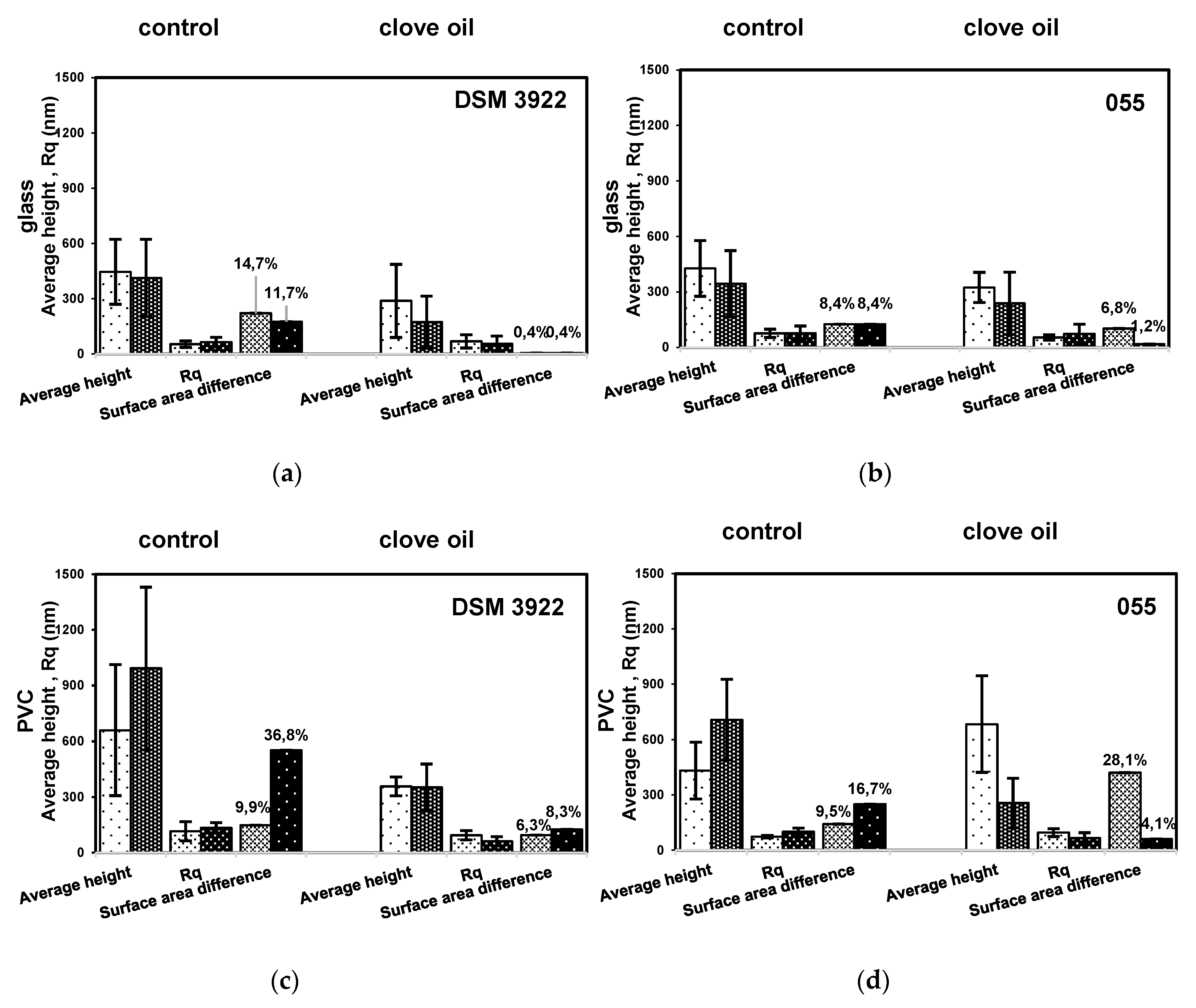
2. Results
3. Discussion
4. Materials and Methods
4.1. Microbial Strains
4.2. Clove Essential Oil and Its Chemical Analysis
4.3. Preparation of Abiotic Surfaces, Biofilm Growth Conditions, and Quantification
4.4. Biofilm Surface Characterization by AFM
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Abee, T.; Kovács, Á.T.; Kuipers, O.P.; van der Veen, S. Biofilm formation and dispersal in Gram-positive bacteria. Curr. Opin. Biotech. 2011, 22, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Bogino, P.C.; Oliva, M.D.M.; Sorroche, F.G.; Giordano, W. The role of bacterial biofilms and surface components in plant-bacterial associations. Int. J. Mol. Sci. 2013, 14, 15838–15859. [Google Scholar] [CrossRef] [PubMed]
- Czaczyk, K.; Myszka, K. Biosynthesis of extracellular polymeric substances (EPS) and its role in microbial biofilm formation. Pol. J. Environ. Stud. 2007, 16, 799–806. [Google Scholar]
- Vu, B.; Chen, M.; Crawford, R.J.; Ivanova, E.P. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009, 14, 2535–2554. [Google Scholar] [CrossRef]
- Auger, S.; Ramarao, N.; Faille, C.; Fouet, A.; Aymerich, S.; Gohar, M. Biofilm formation and cell surface properties among pathogenic and nonpathogenic strains of the Bacillus cereus group. Appl. Environ. Microbiol. 2009, 75, 6616–6618. [Google Scholar] [CrossRef]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 1–18. [Google Scholar] [CrossRef]
- Han, Q.; Song, X.; Zhang, Z.; Fu, J.; Wang, X.; Malakar, P.K.; Liu, H.; Pan, Y.; Zhao, Y. Removal of foodborne pathogen biofilms by acidic electrolyzed water. Front. Microbiol. 2017, 8, 988. [Google Scholar] [CrossRef]
- Srey, S.; Jahid, I.K.; Ha, S.D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Steyn, C.E.; Cameron, M.; Witthuhn, R.C. Occurrence of Alicyclobacillus in the fruit processing environment—A review. Int. J. Food Microbiol. 2011, 147, 1–11. [Google Scholar] [CrossRef]
- Danyluk, M.D.; Friedrich, L.M.; Jouquand, C.; Goodrich-Schneider, R.; Parish, M.E.; Rouseff, R. Prevalence, concentration, spoilage, and mitigation of Alicyclobacillus spp. in tropical and subtropical fruit juice concentrates. Food Microbiol. 2011, 28, 472–477. [Google Scholar] [CrossRef]
- McKnight, I.C.; Eiroa, M.N.U.; Sant’Ana, A.S.; Massaguer, P.R. Alicyclobacillus acidoterrestris in pasteurized exotic Brazilian fruit juices: Isolation, genotypic characterization and heat resistance. Food Microbiol. 2010, 27, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yue, T.; Yuan, Y. Alicyclobacillus contamination in the production line of kiwi products in China. PLoS ONE 2013, 8, e67704. [Google Scholar] [CrossRef] [PubMed]
- Anjos, M.M.; dos Ruiz, S.P.; Nakamura, C.V.; de Abreu Filho, B.A. Resistance of Alicyclobacillus acidoterrestris spores and biofilm to industrial sanitizers. J. Food Protect. 2013, 76, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, M.; Pasvolsky, R.; Zakin, V. External pH is a cue for the behavioral switch that determines surface motility and biofilm formation of Alicyclobacillus acidoterrestris. J. Food Prot. 2014, 77, 1418–1423. [Google Scholar] [CrossRef]
- Tyfa, A.; Kunicka-Styczyńska, A.; Zabielska, J. Evaluation of hydrophobicity and quantitative analysis of biofilm formation by Alicyclobacillus sp. Acta Biochim. Pol. 2015, 62, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zu, Y.; Chen, L.; Shi, X.; Wang, Z.; Sun, Z.; Effert, T. Antimicrobial activity of clove and rosemary essential oils alone and in combination. Phytother. Res. 2007, 21, 989–994. [Google Scholar] [CrossRef]
- Nuñez, L.; D’Aquino, M. Microbicide activity of clove essential oil (Eugenia caryophyllata). Braz. J. Microbiol. 2012, 43, 1255–1260. [Google Scholar] [CrossRef]
- Xu, J.G.; Liu, T.; Hu, Q.P.; Cao, X.M. Chemical composition, antibacterial properties and mechanism of action of essential oil from clove buds against Staphylococcus aureus. Molecules 2016, 21, 1194. [Google Scholar] [CrossRef]
- Gupta, C.; Garg, A.; Uniyal, R.; Gupta, S. Comparison of antimicrobial activities of clove oil and its extract on some food borne microbes. Internet J. Microbiol. 2009, 7, 1–6. [Google Scholar]
- Pinto, E.; Vale-Silva, L.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the clove essential oil from Syzygium aromaticum on Candida, Aspergillus and dermatophyte species. J. Med. Microbiol. 2009, 58, 1454–1462. [Google Scholar] [CrossRef]
- Hoque, M.M.; Bari, M.L.; Juneja, V.K.; Kawamoto, S. Antimicrobial activity of cloves and cinnamon extracts against food borne pathogens and spoilage bacteria, and inactivation of Listeria monocytogenes in ground chicken meat with their essential oils. Rep. National Food Res. Inst. 2008, 72, 9–21. [Google Scholar]
- Kim, Y.-G.; Lee, J.-H.; Gwon, G.; Kim, S.-I.; Park, J.G.; Lee, J. Essential oils and eugenols inhibit biofilm formation and the virulence of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 36377. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.M.; Ahmad, I.; Asif, M.; Tahseen, Q. Influence of clove oil on certain quorum-sensing-regulated functions and biofilm of Pseudomonas aeruginosa and Aeromonas hydrophila. J. Biosci. 2013, 38, 835–844. [Google Scholar] [CrossRef]
- Cui, H.; Zhang, C.; Li, C.; Lin, L. Inhibition of Escherichia coli O157:H7 biofilm on vegetable surface by solid liposomes of clove oil. LWT 2020, 117, 108656. [Google Scholar] [CrossRef]
- Sellamuthu, R. Eugenol. In Encyclopedia of Toxicology. Reference Module in Biomedical Sciences, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 539–541. [Google Scholar]
- Nowak, K.; Ogonowski, J.; Jaworska, M.; Grzesik, K. Clove oil—Properties and applications. Chemik 2012, 66, 145–152. [Google Scholar]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Use of essential oils to inhibit Alicyclobacillus acidoterrestris: A short overview of the literature. Front. Microbiol. 2011, 2, 195. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Campaniello, D.; Speranza, B.; Sinigaglia, M.; Corbo, M.R. Control of Alicyclobacillus acidoterrestris in apple juice by citrus extracts and a mild heat-treatment. Food Control 2013, 31, 553–559. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Combining eugenol and cinnamaldehyde to control the growth of Alicyclobacillus acidoterrestris. Food Control 2010, 21, 172–177. [Google Scholar] [CrossRef]
- Huertas, J.P.; Esteban, M.D.; Antolinos, V.; Palop, A. Combined effect of natural antimicrobials and thermal treatments on Alicyclobacillus acidoterrestris spores. Food Control 2014, 35, 73–78. [Google Scholar] [CrossRef]
- Maldonado, M.C.; Aban, M.P.; Navarro, A.R. Chemicals and lemon essential oil effect on Alicyclobacillus acidoterrestris viability. Braz. J. Microbiol. 2013, 44, 1133–1137. [Google Scholar] [CrossRef]
- Orr, R.V.; Beuchat, L.R. Efficacy of disinfectants in killing spores of Alicyclobacillus acidoterrestris and performance of media for supporting colony development by survivors. J. Food Prot. 2000, 63, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Podolak, R.; Elliott, P.H.; Taylor, B.J.; Khurana, A.; Black, D.G. Destruction of Alicyclobacillus acidoterrestris spores in apple juice on stainless steel surfaces by chemical disinfectants. J. Food Prot. 2009, 72, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Tyfa, A.; Kunicka-Styczyńska, A.; Dąbrowska, J. Activity of compounds of natural origin against Alicyclobacillus acidoterrestris, a common fruit juices contaminant. Biotechnol. Food Sci. 2015, 79, 9–22. [Google Scholar]
- European Pharmacopoeia, 5th ed.; European Union: Brussels, Belgium, 2005.
- Prado, D.B.; dos Anjos Szczerepa, M.M.; Capeloto, O.A.; Astrath, N.G.C.; dos Santos, N.C.A.; Previdelli, I.T.S.; Nakamura, C.V.; Mikcha, J.M.G.; Abreu Filho, B.A. Effect of ultraviolet (UV-C) radiation on spores and biofilms of Alicyclobacillus spp. in industrialized orange juice. Int. J. Food Microbiol. 2019, 305, e108238. [Google Scholar] [CrossRef] [PubMed]
- Prado, D.B.; Fernandes, M.S.; Anjos, M.M.; Tognim, M.C.B.; Nakamura, C.V.; Machinski Jr, M.; Mikcha, J.M.G.; Abreu Filho, B.A. Biofilm-forming ability of Alicyclobacillus spp. isolates from orange juice concentrate processing plant. J. Food Safety 2018, 38, e12466. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Corbo, M.R.; Sinigaglia, M. Inhibition of Alicyclobacillus acidoterrestris spores by natural compounds. Int. J. Food Sci. Technol. 2008, 43, 1271–1275. [Google Scholar] [CrossRef]
- Molva, C.; Baysal, A. Antimicrobial activity of grape seed extract on Alicyclobacillus acidoterrestris DSM 3922 vegetative cells and spores in apple juice. Food Sci. Technol. 2015, 60, 238–245. [Google Scholar] [CrossRef]
- Molva, C.; Baysal, A.H. Evaluation of bioactivity of pomegranate fruit extract against Alicyclobacillus acidoterrestris DSM 3922 vegetative cells and spores in apple juice. Food Sci. Technol. 2015, 62, 989–995. [Google Scholar] [CrossRef]
- de Pascoli, I.C.; dos Anjos, M.M.; da Silva, A.A.; Lorenzetti, F.B.; Cortez, D.A.G.; Mikcha, J.M.G.; Nakamura, T.U.; Nakamura, C.V.; de Abreu Filho, B.A. Piperaceae extracts for controlling Alicyclobacillus acidoterrestris growth in commercial orange juice. Ind. Crop. Prod. 2018, 116, 224–230. [Google Scholar] [CrossRef]
- Piskernik, S.; Klančnik, A.; Demšar, L.; Smole Možina, S.; Jeršek, B. Control of Alicyclobacillus spp. vegetative cells and spores in apple juice with rosemary extracts. Food Control 2016, 60, 205–214. [Google Scholar] [CrossRef]
- Takahashi, T.; Kokubo, R.; Sakaino, M. Antimicrobial activities of eucalyptus leaf extracts and flavonoids from Eucalyptus maculata. Lett. Appl. Microbiol. 2004, 39, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Basson, A.; Flemming, L.A.; Chenia, H.Y. Evaluation of adherence, hydrophobicity, aggregation, and biofilm development of Flavobacterium johnsoniae-like isolates. Microb. Ecol. 2008, 55, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, D.; Brözel, V.S.; von Holy, A. Spore formation in Bacillus subtilis biofilms. J. Food Protect. 2005, 68, 860–865. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Ciuffreda, E.; Sinigaglia, M.; Corbo, M.R. Effects of lysozyme on Alicyclobacillus acidoterrestris under laboratory conditions. Food Sci. Technol. 2014, 49, 224–229. [Google Scholar]
- De Carvalho, A.A.T.; Vanetti, M.C.D.; Mantovani, H.C. Bovicin HC5 reduces thermal resistance of Alicyclobacillus acidoterrestris in acidic mango pulp. J. Appl. Microbiol. 2008, 104, 1685–1691. [Google Scholar] [CrossRef][Green Version]
- Molva, C.; Baysal, A.H. Modeling growth of Alicyclobacillus acidoterrestris DSM 3922 type strain vegetative cells in the apple juice with nisin and lysozyme. Microbiology 2017, 3, 315–322. [Google Scholar] [CrossRef]
- Pei, J.; Yue, T.; Yuan, Y. Control of Alicyclobacillus acidoterrestris in fruit juices by a newly discovered bacteriocin. World J. Microb. Biot. 2014, 30, 855–863. [Google Scholar] [CrossRef]
- Yamazaki, K.; Murakami, M.; Inoue, N.; Matsuda, T. Use of nisin for inhibition of Alicyclobacillus acidoterrestris in acidic drinks. Food Microbiol. 2000, 17, 315–320. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef]
- Smigielski, K.; Raj, A.; Krosowiak, K.; Gruska, R. Chemical composition of the essential oil of Lavandula angustifolia cultivated in Poland. J. Essent. Oil Bear. Plants 2009, 12, 338–347. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corp: Carol Stream, IL, USA, 2007. [Google Scholar]
- Marques, S.C.; Rezende, J.D.G.O.S.; Alves, L.A.D.F.; Silva, B.C.; Alves, E.; Abreu, L.R.D.; Piccoli, R.H. Formation of biofilms by Staphylococcus aureus on stainless steel and glass surfaces and its resistance to some selected chemical sanitizers. Braz. J. Microbiol. 2007, 38, 538–543. [Google Scholar] [CrossRef]
Sample Availability: Samples of the clove buds (Syzygium aromaticum (L.) Merr. and Perry) essential oil are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Compound | RI | Concentration (%) |
|---|---|---|
| Eugenol | 1346 | 86.99 ± 0.09 |
| α- Copaene | 1377 | 00.07 ± 0.01 |
| β-Caryophyllene | 1422 | 08.76 ± 0.04 |
| Cadina-1(6),4-diene | 1468 | 00.04 ± 0.01 |
| α-Humulene | 1452 | 01.91 ± 0.03 |
| ɣ-Muurolene | 1470 | 00.02 ± 0.01 |
| β-Selinene | 1482 | 00.02 ± 0.01 |
| α-Selinene | 1491 | 00.05 ± 0.02 |
| β-Farnesene | 1494 | 00.06 ± 0.01 |
| (Z)-Calamenene | 1509 | 00.11 ± 0.02 |
| δ-Cadinene | 1513 | 00.27 ± 0.03 |
| Cadina-1,4-diene | 1524 | 00.03 ± 0.01 |
| Humulene epoxide | 1594 | 00.11 ± 0.01 |
| Total | 98.44 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunicka-Styczyńska, A.; Tyfa, A.; Laskowski, D.; Plucińska, A.; Rajkowska, K.; Kowal, K. Clove Oil (Syzygium aromaticum L.) Activity against Alicyclobacillus acidoterrestris Biofilm on Technical Surfaces. Molecules 2020, 25, 3334. https://doi.org/10.3390/molecules25153334
Kunicka-Styczyńska A, Tyfa A, Laskowski D, Plucińska A, Rajkowska K, Kowal K. Clove Oil (Syzygium aromaticum L.) Activity against Alicyclobacillus acidoterrestris Biofilm on Technical Surfaces. Molecules. 2020; 25(15):3334. https://doi.org/10.3390/molecules25153334
Chicago/Turabian StyleKunicka-Styczyńska, Alina, Agnieszka Tyfa, Dariusz Laskowski, Aleksandra Plucińska, Katarzyna Rajkowska, and Krystyna Kowal. 2020. "Clove Oil (Syzygium aromaticum L.) Activity against Alicyclobacillus acidoterrestris Biofilm on Technical Surfaces" Molecules 25, no. 15: 3334. https://doi.org/10.3390/molecules25153334
APA StyleKunicka-Styczyńska, A., Tyfa, A., Laskowski, D., Plucińska, A., Rajkowska, K., & Kowal, K. (2020). Clove Oil (Syzygium aromaticum L.) Activity against Alicyclobacillus acidoterrestris Biofilm on Technical Surfaces. Molecules, 25(15), 3334. https://doi.org/10.3390/molecules25153334