
Production of Encecalin in Cell Cultures and Hairy Roots of Helianthella quinquenervis (Hook.) A. Gray

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Results and Discussion
2.1. Germination Treatments and Callus Induction
2.2. Characterization of Cell Suspension Cultures
2.3. Hairy Root Induction
2.4. PCR for the Confirmation of Root Transformation
2.5. Detection and Quantification of Chromenes In Vitro Cultured Cells of H. quinquenervis
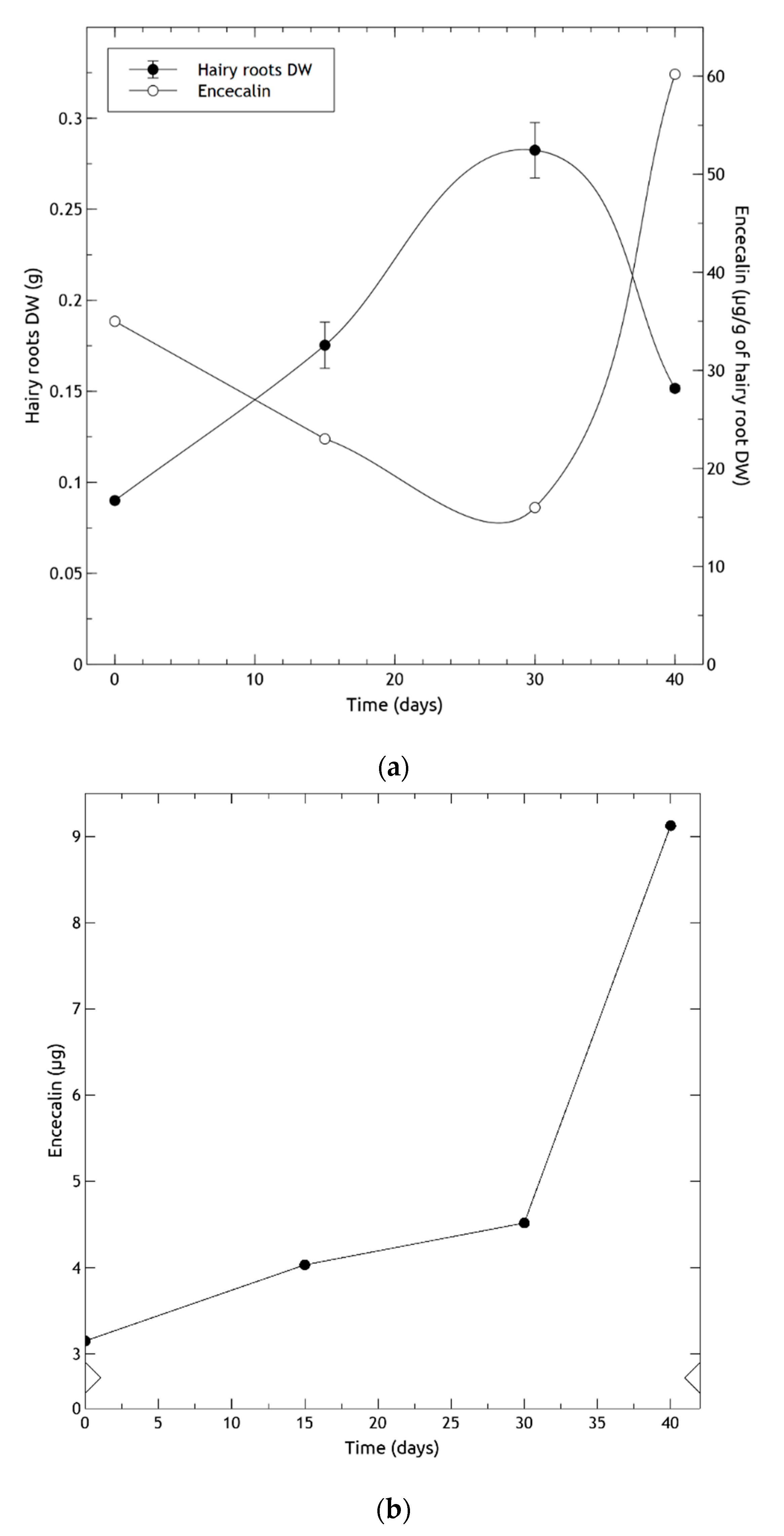
2.6. Hairy Root Growth and Encecalin Quantitative Analysis
3. Materials and Methods
3.1. Seed Germination and Plant In Vitro Growth
3.2. Calli and Cell Suspension Cultures
3.3. Hairy Roots Induction
3.4. Use of PCR for Transformation Analysis
3.5. Establishment of Hairy Roots in Liquid Medium and Growth Analysis
3.6. Detection and Quantification of Encecalin
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weber, W.A. The Genus Helianthella (Compositae). Am. Midl. Nat. 1952, 48, 1–35. [Google Scholar] [CrossRef]
- Bye, R. Medicinal Plants of the Tarahumara Indians of Chihuahua, Mexico. In Two Mummies from Chihuahua: A Multidisciplinary Study; Tyson, R.A., Elñerick, D.V., Eds.; San Diego Museum papers: San Diego, CA, USA, 1985; pp. 77–104. [Google Scholar]
- Rojas, I.S.; Lotina-Hennsen, B.; Mata, R. Effect of lichen metabolites on thylakoid electron transport and photophosphorylation in isolated spinach chloroplasts. J. Nat. Prod. 2000, 63, 1396–1399. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Lugo, M.; Barrientos-Benítez, T.; Luna, B.; Ramirez-Gama, R.; Bye, R.; Linares, E.; Mata, R. Antimicrobial and cytotoxic activities of some crude drug extracts from Mexican medicinal plants. Phytomedicine 1996, 2, 341–347. [Google Scholar] [CrossRef]
- Tapia-Pérez, M.; Tapia-Contreras, A.; Cedillo-Rivera, R.; Osuna, L.; Meckes, M. Screening of Mexican Medicinal Plants for Antiprotozoal Activity—Part II. Pharm. Boil. 2003, 41, 180–183. [Google Scholar] [CrossRef]
- Castañeda, P.; Gómez, L.; Mata, R.; Lotina-Hennsen, B.; Anaya, A.L.; Bye, R. Phytogrowth-Inhibitory and Antifungal Constituents of Helianthella quinquenervis†. J. Nat. Prod. 1996, 59, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Herz, W.; Kulanthaivel, P. Chromans from Helianthella quinquenervis. Phytochemistry 1984, 23, 435–437. [Google Scholar] [CrossRef]
- Gómez, L. Estudio Químico y Biológico Preliminar de Helianthella quinquenervis. Ph.D. Thesis, Faculty of Chemistry, National Autonomous University of Mexico, Mexico City, Mexico, 1993. [Google Scholar]
- Proksch, P.; Rodriguez, E. Chromenes and benzofurans of the Asteraceae, their chemistry and biological significance. Phytochemistry 1983, 22, 2335–2348. [Google Scholar] [CrossRef]
- Isman, M.B. Toxicity and Fate of Acetylchromenes in Pest Insects. In ACS Symposium Series; Amasov, J.T., Philogène, J.R., Morgan, P., Eds.; American Chemical Society: Washington, DC, USA, 1989; pp. 44–58. [Google Scholar]
- Kunze, A.; Aregullin, M.; Rodriguez, E.; Proksch, P. Fate of the chromene encecalin in the interaction of Encelia farinosa and its specialized herbivore Trirhabda geminata. J. Chem. Ecol. 1996, 22, 491–498. [Google Scholar] [CrossRef]
- Alonso-Castro, A.J.; Villarreal, M.L.; Salazar-Olivo, L.A.; Gómez-Sánchez, M.; Dominguez, F.; García-Carrancá, A. Mexican medicinal plants used for cancer treatment: Pharmacological, phytochemical and ethnobotanical studies. J. Ethnopharmacol. 2011, 133, 945–972. [Google Scholar] [CrossRef]
- Lotina-Hennsen, B.; King-Díaz, B.; Aguilar, M.; Terrones, M.G.H. Plant secondary metabolites. Targets and mechanisms of allelopathy. In Allelopathy: A Physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., González, L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 229–265. [Google Scholar]
- Merrill, G.B. Eupatoriochromene and encecalin, plant growth regulators from yellow starthistle (Centaurea solstitialis L.). J. Chem. Ecol. 1989, 15, 2073–2087. [Google Scholar] [CrossRef]
- Harel, D.; Schepmann, D.; Prinz, H.; Brun, R.; Schmidt, T.J.; Wünsch, B. Natural Product Derived Antiprotozoal Agents: Synthesis, Biological Evaluation, and Structure–Activity Relationships of Novel Chromene and Chromane Derivatives. J. Med. Chem. 2013, 56, 7442–7448. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Chappell, J. Metabolic engineering of natural products in plants; tools of the trade and challenges for the future. Curr. Opin. Biotechnol. 2008, 19, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Balandrin, M.; Klocke, J.; Wurtele, E.S.; Bollinger, W. Natural plant chemicals: Sources of industrial and medicinal materials. Science 1985, 228, 1154–1160. [Google Scholar] [CrossRef]
- Vanisree, M.; Lee, C.Y.; Lo, S.F.; Nalawade, S.M.; Lin, C.Y.; Tsay, H.S. Studies on the production of some important secondary metabolites from medicinal plants by plant tissue cultures. Bot. Bull. Acad. Sin. 2004, 45, 1–22. [Google Scholar]
- Ochoa-Villarreal, M.; Howat, S.; Hong, S.; Jang, M.O.; Jin, Y.-W.; Lee, E.-K.; Loake, G.J. Plant cell culture strategies for the production of natural products. BMB Rep. 2016, 49, 149–158. [Google Scholar] [CrossRef]
- Roychowdhury, D.; Majumder, A.; Jha, S. Agrobacterium rhizogenes-Mediated Transformation in Medicinal Plants: Prospects and Challenges. In Biotechnology for Medicinal Plants; Chandra, S., Lata, H., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 29–68. [Google Scholar]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.-M.; Ritala, A.; Cardon, F. Hairy Root Cultures-A Versatile Tool with Multiple Applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef]
- Christey, M.; Braun, R.H. Production of hairy root cultures and transgenic plants by Agrobacterium rhizogenes-mediated transformation. In Methods in Molecular Biology; Peña, L., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2005; pp. 47–60. [Google Scholar]
- Nilsson, O.; Olsson, O. Getting to the root: The role of the Agrobacterium rhizogenes rol genes in the formation of hairy roots. Physiol. Plant. 1997, 100, 463–473. [Google Scholar] [CrossRef]
- Guillon, S.; Trémouillaux-Guiller, J.; Pati, P.K.; Rideau, M.; Gantet, P. Hairy root research: Recent scenario and exciting prospects. Curr. Opin. Plant Boil. 2006, 9, 341–346. [Google Scholar] [CrossRef]
- Chandra, S.; Chandra, R. Engineering secondary metabolite production in hairy roots. Phytochem. Rev. 2011, 10, 371–395. [Google Scholar] [CrossRef]
- Wilczańska-Barska, A.; Królicka, A.; Głód, D.; Majdan, M.; Kawiak, A.; Krauze-Baranowska, M. Enhanced accumulation of secondary metabolites in hairy root cultures of Scutellaria lateriflora following elicitation. Biotechnol. Lett. 2012, 34, 1757–1763. [Google Scholar] [CrossRef]
- Aguado, M.; Martínez-Sánchez, J.J.; Reig-Armiñana, J.; García-Breijo, F.J.; Franco, J.A.; Vicente, M.J. Morphology, anatomy and germination response of heteromorphic achenes of Anthemis chrysantha J. Gay (Asteraceae), a critically endangered species. Seed Sci. Res. 2011, 21, 283–294. [Google Scholar] [CrossRef]
- Mathur, S.; Shekhawat, G.S. Establishment and characterization of Stevia rebaudiana (Bertoni) cell suspension culture: An in vitro approach for production of stevioside. Acta Physiol. Plant. 2012, 35, 931–939. [Google Scholar] [CrossRef]
- Yue, W.; Ming, Q.-L.; Lin, B.; Rahman, K.; Zheng, C.-J.; Han, T.; Qin, L.-P. Medicinal plant cell suspension cultures: Pharmaceutical applications and high-yielding strategies for the desired secondary metabolites. Crit. Rev. Biotechnol. 2016, 36, 215–232. [Google Scholar] [CrossRef] [PubMed]
- Udomsin, O.; Yusakul, G.; Kitisripanya, T.; Juengwatanatrakul, T.; Putalun, W. The Deoxymiroestrol and Isoflavonoid Production and Their Elicitation of Cell Suspension Cultures of Pueraria candollei var. mirifica: From Shake Flask to Bioreactor. Appl. Biochem. Biotechnol. 2019, 190, 57–72. [Google Scholar] [CrossRef]
- Srinivasan, A.; Vijayakumar, S.; Raman, K. Rational metabolic engineering for enhanced alpha-tocopherol production in Helianthus annuus cell culture. Biochem. Eng. J. 2019, 151, 107256. [Google Scholar] [CrossRef]
- Georgiev, M.I.; Ludwig-Müller, J.; Alipieva, K.; Lippert, A. Sonication-assisted Agrobacterium rhizogenes-mediated transformation of Verbascum xanthophoeniceum Griseb. for bioactive metabolite accumulation. Plant Cell Rep. 2011, 30, 859–866. [Google Scholar] [CrossRef]
- Georgiev, M.; Georgiev, V.; Weber, J.; Bley, T.H.; Ilieva, M.; Pavlov, A. Agrobacterium Rhizogenes-Mediated Genetic Transformations: A Powerful Tool for the Production of Metabolites; Wolf, T., Koch, J., Eds.; Nova Science Publishers: New York, NY, USA, 2008. [Google Scholar]
- Thwe, A.; Arasu, M.V.; Li, X.; Park, C.H.; Kim, J.-B.; Al-Dhabi, N.A.; Park, S.U. Effect of Different Agrobacterium rhizogenes Strains on Hairy Root Induction and Phenylpropanoid Biosynthesis in Tartary Buckwheat (Fagopyrum tataricum Gaertn). Front. Microbiol. 2016, 7, 1–59. [Google Scholar] [CrossRef]
- Vojin, T.; Snežana, M.; Aleksandar, C.; Marija, P.; Milana, T.; Dragana, A.; Jovan, T.; Angelina, S. Production of hairy root cultures of lettuce (Lactuca sativa L.). Open Life Sci. 2014, 9, 1196–1205. [Google Scholar] [CrossRef]
- Dilshad, E.; Ismail, H.; Haq, I.U.; Cusido, R.M.; Palazon, J.; Ramirez-Estrada, K.; Mirza, B. Rol genes enhance the biosynthesis of antioxidants in Artemisia carvifolia Buch. BMC Plant Boil. 2016, 16, 125. [Google Scholar] [CrossRef]
- Sarkar, S.; Ghosh, I.; Roychowdhury, D.; Jha, S. The Effects of rol Genes of Agrobacterium rhizogenes on Morphogenesis and Secondary Metabolite Accumulation in Medicinal Plants. In Biotechnological Approaches for Medicinal and Aromatic Plants: Conservation, Genetic Improvement and Utilization; Kumar, N., Ed.; Springer: Singapore, 2018; pp. 27–51. [Google Scholar]
- Bhojwani, S.S.; Dantu, P.K. Plant Tissue Culture: An Introductory Text; Springer: London, UK, 2013. [Google Scholar]
- Saad, I.; Díaz, E.; Chávez, I.; Reyes-Chilpa, R.; Rubluo, A.; Jiménez-Estrada, M. Antifungal monoterpene production in elicited cell suspension cultures of Piqueria trinervia. Phytochemistry 2000, 55, 51–57. [Google Scholar] [CrossRef]
- Łuczkiewicz, M.; Zarate, R.; Dembińska-Migas, W.; Migas, P.; Verpoorte, R. Production of pulchelin E in hairy roots, callus and suspension cultures of Rudbeckia hirta L. Plant Sci. 2002, 163, 91–100. [Google Scholar] [CrossRef]
- Kawoosa, T.; Singh, H.; Kumar, A.; Sharma, S.K.; Devi, K.; Dutt, S.; Vats, S.K.; Sharma, M.; Ahuja, P.S.; Kumar, S. Light and temperature regulated terpene biosynthesis: Hepatoprotective monoterpene picroside accumulation in Picrorhiza kurrooa. Funct. Integr. Genom. 2010, 10, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Trick, H.N.; Finer, J. SAAT: Sonication-assisted Agrobacterium-mediated transformation. Transg. Res. 1997, 6, 329–336. [Google Scholar] [CrossRef]
- Santarém, E.R.; Trick, H.N.; Essig, J.S.; Finer, J. Sonication-assisted Agrobacterium- mediated transformation of soybean immature cotyledons: Optimization of transient expression. Plant Cell Rep. 1998, 17, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Beranová, M.; Rakouský, S.; Vávrová, Z.; Skalicky, T. Sonication assisted Agrobacterium-mediated transformation enhances the transformation efficiency in flax (Linum usitatissimum L.). Plant Cell Tissue Organ Cult. 2008, 94, 253–259. [Google Scholar] [CrossRef]
- Wu, X.; Shi, H.; Chen, X.; Liu, Y.; Guo, Z. Establishment of Agrobacterium-mediated transformation of seashore paspalum (Paspalum vaginatum O. Swartz). Vitr. Cell. Dev. Boil.—Anim. 2018, 54, 545–552. [Google Scholar] [CrossRef]
- Danphitsanuparn, P.; Boonsnongcheep, P.; Boriboonkaset, T.; Chintapakorn, Y.; Prathanturarug, S. Effects of Agrobacterium rhizogenes strains and other parameters on production of isoflavonoids in hairy roots of Pueraria candollei Grah. ex Benth. var. candollei. Plant Cell Tissue Organ Cult. 2012, 111, 315–322. [Google Scholar] [CrossRef]
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Malik, S.; Cusido, R.M.; Mirjalili, M.H.; Moyano, E.; Palazon, J.; Bonfill, M. Production of the anticancer drug taxol in Taxus baccata suspension cultures: A review. Process. Biochem. 2011, 46, 23–34. [Google Scholar] [CrossRef]
- Stafford, A. Natural Products and Metabolites from Plants and Plant Tissue Cultures. In Plant Cell and Tissue Culture; Stafford, A., Warren, G., Eds.; Open University Press: Buckingham, UK, 1991; pp. 124–238. [Google Scholar]
- Danova, K.; Cellarova, E.; Macková, A.; Daxnerová, Z.; Kapchina-Toteva, V. In vitro culture of Hypericum rumeliacum Boiss. and production of phenolics and flavonoids. Vitr. Cell. Dev. Boil.—Anim. 2010, 46, 422–429. [Google Scholar] [CrossRef]
- Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Proksch, P.; Wray, V.; Isman, M.B.; Rahaus, I. Ontogenetic variation of biologically active natural products in Ageratina adenophora. Phytochemistry 1990, 29, 453–457. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Duncan, D.R.; Widholm, J.M.; Pollard, J.W.; Walker, J.M. Techniques for Selecting Mutants from Plant Tissue Cultures. In Plant Cell and Tissue Culture; Pollard, J.W., Walker, J.M., Eds.; The Humana Press: Clifton, NJ, USA, 2003; pp. 443–454. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Boil. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Takashi, A.; Takashi, H.; Satoshi, T.; Atsuhiro, O. Putative start codon TTG for the regulatory protein VirG of the hairy-root-inducing plasmid pRiA4. Gene 1989, 78, 173–178. [Google Scholar] [CrossRef]
Sample Availability: Not available. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method of Transformation | Number of Explants Transformed/Total Explant | Range of Roots/Explant |
|---|---|---|
| Prick | 6/35 | 1–3 |
| Sonication | 18/45 | 1–5 |
| Co-culture | 20/30 | 1–9 |
| Culture | Encecalin (µg/g FW) |
|---|---|
| Calli 1 (photoperiod) | 165 ± 7 |
| Calli (darkness) | 290 ± 3 |
| Cell suspension culture | 327 ± 14 |
| Hairy roots (co-culture) | 1086 ± 31 |
| Gene | Expected Amplified Product (bp) | Sequence |
|---|---|---|
| rolC | 534 | 1: 5′-TAA CAT GGC TGA AGA CGA CC-3′ 2: 5′-AAA CTT GCA CTC GCC ATG CC-3′ |
| aux1 | 350 | 1: 5′-TTC GAA GGA AGC TTG TCA GAA-3′ 2: 5′-CTT AAA TCC GTC TGA CCA TAG-3′ |
| virG1 | 350 | 1: 5′- ACT GAA TAT CAG GCA ACG CC-3′ 2: 5′- GCG TCA AAG AAA TAG CCA GC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Altamirano, J.M.; Ugidos, I.F.; Palazón, J.; Bonfill, M.; García-Angulo, P.; Álvarez, J.; Acebes, J.L.; Bye, R.; Encina, A. Production of Encecalin in Cell Cultures and Hairy Roots of Helianthella quinquenervis (Hook.) A. Gray. Molecules 2020, 25, 3231. https://doi.org/10.3390/molecules25143231
Hernández-Altamirano JM, Ugidos IF, Palazón J, Bonfill M, García-Angulo P, Álvarez J, Acebes JL, Bye R, Encina A. Production of Encecalin in Cell Cultures and Hairy Roots of Helianthella quinquenervis (Hook.) A. Gray. Molecules. 2020; 25(14):3231. https://doi.org/10.3390/molecules25143231
Chicago/Turabian StyleHernández-Altamirano, J. Mabel, Irene F. Ugidos, Javier Palazón, Mercedes Bonfill, Penélope García-Angulo, Jesús Álvarez, José L. Acebes, Robert Bye, and Antonio Encina. 2020. "Production of Encecalin in Cell Cultures and Hairy Roots of Helianthella quinquenervis (Hook.) A. Gray" Molecules 25, no. 14: 3231. https://doi.org/10.3390/molecules25143231
APA StyleHernández-Altamirano, J. M., Ugidos, I. F., Palazón, J., Bonfill, M., García-Angulo, P., Álvarez, J., Acebes, J. L., Bye, R., & Encina, A. (2020). Production of Encecalin in Cell Cultures and Hairy Roots of Helianthella quinquenervis (Hook.) A. Gray. Molecules, 25(14), 3231. https://doi.org/10.3390/molecules25143231