Lignans, Amides, and Saponins from Haplophyllum tuberculatum and Their Antiprotozoal Activity

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results and Discussion
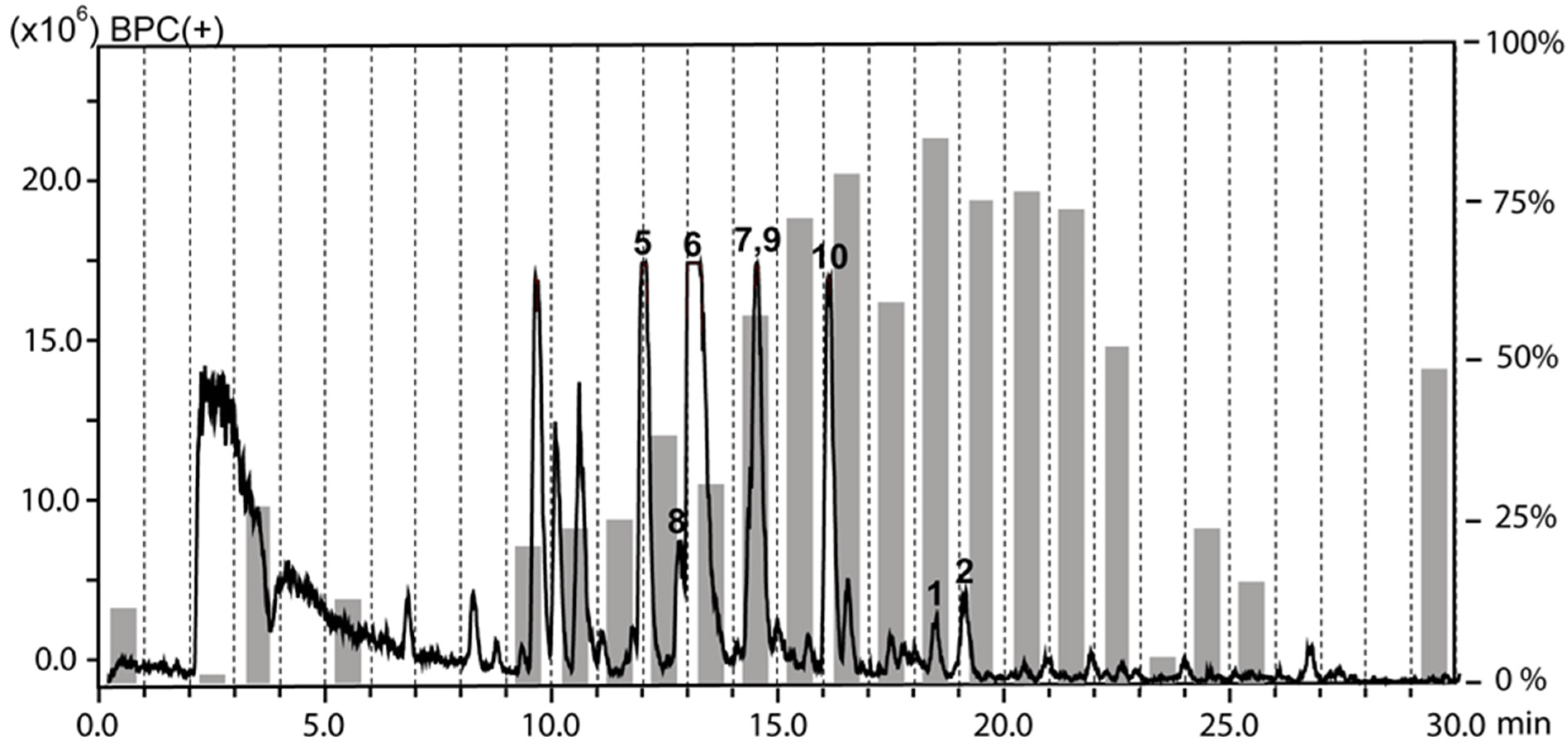
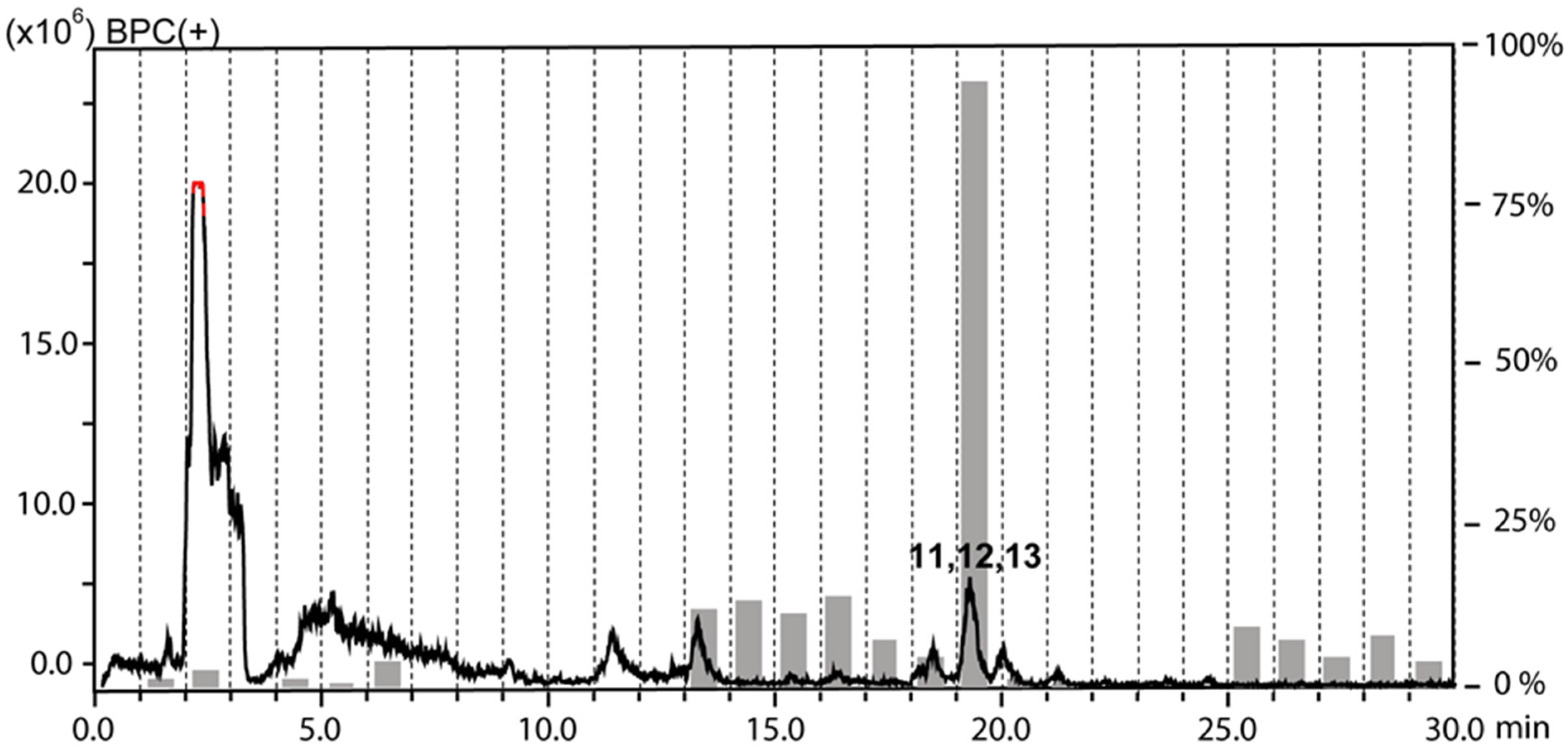
2.1. Extraction and HPLC-based Activity Profiling
2.2. Compound Isolation and Structure Elucidation
2.3. Comparison to Previously Reported Compounds
2.4. Biological Testing
2.4.1. Activity against Leishmania Donovani Axenic Amastigotes
2.4.2. Activity against Plasmodium Falciparum
2.4.3. Activity against Trypanosoma Brucei Rhodesiense
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Material
3.3. Extraction
3.4. Microfractionation for Activity Profiling
3.5. Preparative Isolation
3.6. Activity against Leishmania Donovani Axenic Amastigotes
3.7. Activity against Leishmania Donovani Intramacrophage Amastigotes
3.8. Activity against Plasmodium Falciparum
3.9. Activity against Trypanosoma Brucei Rhodesiense
3.10. In Vitro Cytotoxicity with L-6 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hotez, P.J.; Alvarado, M.; Basáñez, M.-G.; Bolliger, I.; Bourne, R.; Boussinesq, M.; Brooker, S.J.; Brown, A.S.; Buckle, G.; Budke, C.M.; et al. The global burden of disease study 2010: Interpretation and implications for the neglected tropical diseases. PLoS Negl. Trop. Dis. 2014, 8, e2865. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Vos, T.; Lozano, R.; Naghavi, M.; Flaxman, A.D.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S. Disability-adjusted life years (DALYs) for 291 diseases and injuries in 21 regions, 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2197–2223. [Google Scholar] [CrossRef]
- Alves, F.; Bilbe, G.; Blesson, S.; Goyal, V.; Monnerat, S.; Mowbray, C.; Ouattara, G.M.; Pécoul, B.; Rijal, S.; Rode, J. Recent development of visceral leishmaniasis treatments: Successes, pitfalls, and perspectives. Clin. Microbiol. Rev. 2018, 31, e00048-18. [Google Scholar] [CrossRef] [PubMed]
- Balasegaram, M.; Ritmeijer, K.; Lima, M.A.; Burza, S.; Ortiz Genovese, G.; Milani, B.; Gaspani, S.; Potet, J.; Chappuis, F. Liposomal amphotericin B as a treatment for human leishmaniasis. Expert Opin. Emerg. Drugs 2012, 17, 493–510. [Google Scholar] [CrossRef] [PubMed]
- WHO|Leishmaniasis in High-Burden Countries: An Epidemiological Update Based on Data Reported in 2014. Available online: http://www.who.int/leishmaniasis/resources/who_wer9122/en/ (accessed on 12 May 2020).
- Kato, K.C.; Morais-Teixeira, E.; Reis, P.G.; Silva-Barcellos, N.M.; Salaün, P.; Campos, P.P.; Dias Corrêa-Junior, J.; Rabello, A.; Demicheli, C.; Frézard, F. Hepatotoxicity of pentavalent antimonial drug: Possible role of residual Sb(III) and protective effect of ascorbic acid. Antimicrob. Agents Chemother. 2014, 58, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, D.H.; Savioli, L.; Engels, D. Neglected tropical diseases: Progress towards addressing the chronic pandemic. Lancet Lond. Engl. 2017, 389, 312–325. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet Lond. Engl. 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Mahmoud, A.B.; Mäser, P.; Kaiser, M.; Hamburger, M.; Khalid, S. Mining Sudanese Medicinal Plants for Antiprotozoal Agents. Front. Pharmacol. 2020, 11, 865. [Google Scholar] [CrossRef]
- Hamdi, A.; Majouli, K.; Abdelhamid, A.; Marzouk, B.; Belghith, H.; Chraief, I.; Bouraoui, A.; Marzouk, Z.; Heyden, Y.V. Pharmacological activities of the organic extracts and fatty acid composition of the petroleum ether extract from Haplophyllum tuberculatum leaves. J. Ethnopharmacol. 2018, 216, 97–103. [Google Scholar] [CrossRef]
- Eissa, T.F.; González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. Biological activity of HPLC-characterized ethanol extract from the aerial parts of Haplophyllum tuberculatum. Pharm. Biol. 2014, 52, 151–156. [Google Scholar] [CrossRef]
- Al-Burtamani, S.K.S.; Fatope, M.O.; Marwah, R.G.; Onifade, A.K.; Al-Saidi, S.H. Chemical composition, antibacterial and antifungal activities of the essential oil of Haplophyllum tuberculatum from Oman. J. Ethnopharmacol. 2005, 96, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Al-Rehaily, A.J.; Al-Howiriny, T.A.; Ahmad, M.S.; Al-Yahya, M.A.; El-Feraly, F.S.; Hufford, C.D.; McPhail, A.T. Alkaloids from Haplophyllum tuberculatum. Phytochemistry 2001, 57, 597–602. [Google Scholar] [CrossRef]
- Hamdi, A.; Viane, J.; Mahjoub, M.A.; Majouli, K.; Gad, M.H.H.; Kharbach, M.; Demeyer, K.; Marzouk, Z.; Heyden, Y.V. Polyphenolic contents, antioxidant activities and UPLC-ESI-MS analysis of Haplophyllum tuberculatum A. Juss leaves extracts. Int. J. Biol. Macromol. 2018, 106, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, S.; Seradj, H. Justicidin B: A Promising Bioactive Lignan. Mol. Basel Switz. 2016, 21, 820. [Google Scholar] [CrossRef] [PubMed]
- El-Tahir, A.; Satti, G.M.; Khalid, S.A. Antiplasmodial activity of selected sudanese medicinal plants with emphasis on Acacia nilotica. Phytother. Res. PTR 1999, 13, 474–478. [Google Scholar] [CrossRef]
- Khalid, S.A.; Farouk, A.; Geary, T.G.; Jensen, J.B. Potential antimalarial candidates from African plants: An in vitro approach using Plasmodium falciparum. J. Ethnopharmacol. 1986, 15, 201–209. [Google Scholar] [CrossRef]
- Hamdi, A.; Bero, J.; Beaufay, C.; Flamini, G.; Marzouk, Z.; Vander Heyden, Y.; Quetin-Leclercq, J. In vitro antileishmanial and cytotoxicity activities of essential oils from Haplophyllum tuberculatum A. Juss leaves, stems and aerial parts. BMC Complement. Altern. Med. 2018, 18, 60. [Google Scholar] [CrossRef]
- Gertsch, J.; Tobler, R.T.; Brun, R.; Sticher, O.; Heilmann, J. Antifungal, antiprotozoal, cytotoxic and piscicidal properties of Justicidin B and a new arylnaphthalide lignan from Phyllanthus piscatorum. Planta Med. 2003, 69, 420–424. [Google Scholar]
- Potterat, O.; Hamburger, M. Combined use of extract libraries and HPLC-based activity profiling for lead discovery: Potential, challenges, and practical considerations. Planta Med. 2014, 80, 1171–1181. [Google Scholar] [CrossRef]
- Nguyen, P.H.; Le, T.V.T.; Kang, H.W.; Chae, J.; Kim, S.K.; Kwon, K.; Seo, D.B.; Lee, S.J.; Oh, W.K. AMP-activated protein kinase (AMPK) activators from Myristica fragrans (nutmeg) and their anti-obesity effect. Bioorg. Med. Chem. Lett. 2010, 20, 4128–4131. [Google Scholar] [CrossRef]
- Hattori, M.; Hada, S.; KAWATA, Y.; TEZUKA, Y.; KIKUCHI, T.; NAMBA, T. New 2, 5-bis-aryl-3, 4-dimethyltetrahydrofuran lignans from the aril of Myristica fragrans. Chem. Pharm. Bull. (Tokyo) 1987, 35, 3315–3322. [Google Scholar] [CrossRef]
- Nakano, Y.; Nasu, M.; Kano, M.; Kameoka, H.; Okuyama, T.; Nishizawa, M.; Ikeya, Y. Lignans from guaiac resin decrease nitric oxide production in interleukin 1β-treated hepatocytes. J. Nat. Med. 2017, 71, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Chang, F.-R.; Yen, H.-F.; Wu, Y.-C. Amides from stems of Annona cherimola. Phytochemistry 1998, 49, 1443–1447. [Google Scholar] [CrossRef]
- King, R.R.; Calhoun, L.A. Characterization of cross-linked hydroxycinnamic acid amides isolated from potato common scab lesions. Phytochemistry 2005, 66, 2468–2473. [Google Scholar] [CrossRef] [PubMed]
- Iwasa, K.; Takahashi, T.; Nishiyama, Y.; Moriyasu, M.; Sugiura, M.; Takeuchi, A.; Tode, C.; Tokuda, H.; Takeda, K. Online structural elucidation of alkaloids and other constituents in crude extracts and cultured cells of Nandina domestica by combination of LC-MS/MS, LC-NMR, and LC-CD analyses. J. Nat. Prod. 2008, 71, 1376–1385. [Google Scholar] [CrossRef]
- Maciel, J.K.S.; Chaves, O.S.; Brito Filho, S.G.; Teles, Y.C.F.; Fernandes, M.G.; Assis, T.S.; Fernandes, P.D.; de Andrade, A.P.; Felix, L.P.; Silva, T.M.S.; et al. New Alcamide and Anti-oxidant Activity of Pilosocereus gounellei A. Weber ex K. Schum. Bly. ex Rowl. (Cactaceae). Mol. Basel Switz. 2015, 21, 11. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, H.-Y.; Wang, Y.-M.; Tian, T.; Wu, W.-M.; Zhou, M.; Meng, X.-G.; Ruan, H.-L. Neuroprotective Lignans from the Fruits of Schisandra bicolor var. tuberculata. J. Nat. Prod. 2017, 80, 1117–1124. [Google Scholar] [CrossRef]
- Agrawal, P.K.; Jain, D.C.; Pathak, A.K. NMR spectroscopy of steroidal sapogenins and steroidal saponins: An update. Magn. Reson. Chem. 1995, 33, 923–953. [Google Scholar] [CrossRef]
- Nuevo, M.; Cooper, G.; Sandford, S.A. Deoxyribose and deoxysugar derivatives from photoprocessed astrophysical ice analogues and comparison to meteorites. Nat. Commun. 2018, 9, 5276. [Google Scholar] [CrossRef]
- Farid, H.; Haslinger, E.; Kunert, O.; Wegner, C.; Hamburger, M. New steroidal glycosides from Balanites aegyptiaca. Helv. Chim. Acta 2002, 85, 1019–1026. [Google Scholar] [CrossRef]
- Pettit, G.R.; Doubek, D.L.; Herald, D.L.; Numata, A.; Takahasi, C.; Fujiki, R.; Miyamoto, T. Isolation and structure of cytostatic steroidal saponins from the African medicinal plant Balanites aegyptica. J. Nat. Prod. 1991, 54, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Rédei, D.; Kúsz, N.; Jedlinszki, N.; Blazsó, G.; Zupkó, I.; Hohmann, J. Bioactivity-Guided Investigation of the Anti-Inflammatory Activity of Hippophae rhamnoides Fruits. Planta Med. 2018, 84, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.A.; Waterman, P.G. Alkaloid, lignan and flavonoid constituents of Haplophyllum tuberculatum from Sudan. Planta Med. 1981, 43, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Sheriha, G.M.; Amer, K.M.A. Lignans of haplophyllum tuberculatum. Phytochemistry 1984, 23, 151–153. [Google Scholar] [CrossRef]
- Martin-Tanguy, J.; Cabanne, F.; Perdrizet, E.; Martin, C. The distribution of hydroxycinnamic acid amides in flowering plants. Phytochemistry 1978, 17, 1927–1928. [Google Scholar] [CrossRef]
- Yang, Y.-L.; Chang, F.-R.; Wu, Y.-C. Annosqualine: A novel alkaloid from the stems of Annona squamosa. Helv. Chim. Acta 2004, 87, 1392–1399. [Google Scholar] [CrossRef]
- Lajide, L.; Escoubas, P.; Mizutani, J. Termite antifeedant activity in Xylopia aethiopica. Phytochemistry 1995, 40, 1105–1112. [Google Scholar] [CrossRef]
- Lin, I.J.; Yeh, H.C.; Cham, T.M.; Chen, C.Y. A new butanolide from the leaves of Cinnamomum reticulatum. Chem. Nat. Compd. 2011, 47, 43. [Google Scholar] [CrossRef]
- Tanaka, H.; Nakamura, T.; Ichino, K.; Ito, K. A phenolic amide from Actinodaphne longifolia. Phytochemistry 1989, 28, 2516–2517. [Google Scholar] [CrossRef]
- Hussain, S.F.; Gözler, B.; Shamma, M.; Gözler, T. Feruloyltyramine from Hypecoum. Phytochemistry 1980, 21, 2979–2980. [Google Scholar] [CrossRef]
- Sakakibara, I.; Katsuhara, T.; Ikeya, Y.; Hayashi, K.; Mitsuhashi, H. Cannabisin A, an arylnaphthalene lignanamide from fruits of Cannabis sativa. Phytochemistry 1991, 30, 3013–3016. [Google Scholar] [CrossRef]
- Munoz, O.; Piovano, M.; Garbarino, J.; Hellwing, V.; Breitmaier, E. Tropane alkaloids from Schizanthus litoralis. Phytochemistry 1996, 43, 709–713. [Google Scholar] [CrossRef]
- Tian, J.-L.; Liang, X.; Gao, P.-Y.; Li, D.-Q.; Sun, Q.; Li, L.-Z.; Song, S.-J. Two new alkaloids from Portulaca oleracea and their cytotoxic activities. J. Asian Nat. Prod. Res. 2014, 16, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, W.; Yang, X.; Xiu, F.; Xu, H.; Ying, X.; Stien, D. An isoindole alkaloid from Portulaca oleracea L. Nat. Prod. Res. 2018, 32, 2431–2436. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.-S.; Shi, L.-S.; Kuo, S.-C. Alkaloids and other constituents from Tribulus terrestris. Phytochemistry 1999, 50, 1411–1415. [Google Scholar] [CrossRef]
- Raj, M.K.; Balachandran, C.; Duraipandiyan, V.; Agastian, P.; Ignacimuthu, S.; Vijayakumar, A. Isolation of terrestribisamide from Peltophorum pterocarpum (DC.) Baker ex. K. Heyne and its antimicrobial, antioxidant, and cytotoxic activities. Med. Chem. Res. 2013, 22, 3823–3830. [Google Scholar]
- Choi, S.W.; Lee, S.K.; Kim, E.O.; Oh, J.H.; Yoon, K.S.; Parris, N.; Hicks, K.B.; Moreau, R.A. Antioxidant and antimelanogenic activities of polyamine conjugates from corn bran and related hydroxycinnamic acids. J. Agric. Food Chem. 2007, 55, 3920–3925. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-W.; Nakanishi, K. The structures of balanitins, potent molluscicides isolated from Balanites aegyptiaca. Tetrahedron 1982, 38, 513–519. [Google Scholar] [CrossRef]
- Rizk, M.Z.; Metwally, N.S.; Hamed, M.A.; Mohamed, A.M. Correlation between steroid sex hormones, egg laying capacity and cercarial shedding in Biomphalaria alexandrina snails after treatment with Haplophyllum tuberculatum. Exp. Parasitol. 2012, 132, 171–179. [Google Scholar] [CrossRef]
- da Silva Filho, A.A.; Costa, E.S.; Cunha, W.R.; e Silva, M.L.A.; Nanayakkara, N.P.D.; Bastos, J.K. In vitro antileishmanial and antimalarial activities of tetrahydrofuran lignans isolated from Nectandra megapotamica (Lauraceae). Phytother. Res. PTR 2008, 22, 1307–1310. [Google Scholar] [CrossRef]
- de Oliveira, R.B.; Vaz, A.; Alves, R.O.; Liarte, D.B.; Donnici, C.L.; Romanha, A.J.; Zani, C.L. Arylfurans as potential Trypanosoma cruzi trypanothione reductase inhibitors. Memórias Inst. Oswaldo Cruz 2006, 101, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Tovar, J.; Wilkinson, S.; Mottram, J.C.; Fairlamb, A.H. Evidence that trypanothione reductase is an essential enzyme in Leishmania by targeted replacement of the tryA gene locus. Mol. Microbiol. 1998, 29, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.O.F. Trypanothione reductase: A viable chemotherapeutic target for antitrypanosomal and antileishmanial drug design. Drug Target Insights 2007, 2, 117739280700200000. [Google Scholar] [CrossRef]
- Zhang, L.P.; Ji, Z.Z. Synthesis, antiinflammatory and anticancer activity of cinnamic acids, their derivatives and analogues. Yao Xue Xue Bao 1992, 27, 817–823. [Google Scholar] [PubMed]
- Ramos-Nino, M.E.; Clifford, M.N.; Adams, M.R. Quantitative structure activity relationship for the effect of benzoic acids, cinnamic acids and benzaldehydes on Listeria monocytogenes. J. Appl. Bacteriol. 1996, 80, 303–310. [Google Scholar] [CrossRef]
- Pedersen, H.A.; Steffensen, S.K.; Christophersen, C.; Mortensen, A.G.; Jørgensen, L.N.; Niveyro, S.; De Troiani, R.M.; Rodríguez-Enríquez, R.J.; Barba-de la Rosa, A.P.; Fomsgaard, I.S. Synthesis and quantitation of six phenolic amides in Amaranthus spp. J. Agric. Food Chem. 2010, 58, 6306–6311. [Google Scholar] [CrossRef]
- Natella, F.; Nardini, M.; Di Felice, M.; Scaccini, C. Benzoic and cinnamic acid derivatives as antioxidants: Structure—Activity relation. J. Agric. Food Chem. 1999, 47, 1453–1459. [Google Scholar] [CrossRef]
- Potterat, O.; Hamburger, M. Concepts and technologies for tracking bioactive compounds in natural product extracts: Generation of libraries, and hyphenation of analytical processes with bioassays. Nat. Prod. Rep. 2013, 30, 546–564. [Google Scholar] [CrossRef]
- Cunningham, I. New culture medium for maintenance of tsetse tissues and growth of trypanosomatids. J. Protozool. 1977, 24, 325–329. [Google Scholar] [CrossRef]
- Desjardins, R.E.; Canfield, C.J.; Haynes, J.D.; Chulay, J.D. Quantitative assessment of antimalarial activity in vitro by a semiautomated microdilution technique. Antimicrob. Agents Chemother. 1979, 16, 710–718. [Google Scholar] [CrossRef]
- Ponnudurai, T.; Leeuwenberg, A.D.; Meuwissen, J.H. Chloroquine sensitivity of isolates of Plasmodium falciparum adapted to in vitro culture. Trop. Geogr. Med. 1981, 33, 50–54. [Google Scholar] [PubMed]
- Baltz, T.; Baltz, D.; Giroud, C.; Crockett, J. Cultivation in a semi-defined medium of animal infective forms of Trypanosoma brucei, T. equiperdum, T. evansi, T. rhodesiense and T. gambiense. EMBO J. 1985, 4, 1273–1277. [Google Scholar] [CrossRef] [PubMed]
- Räz, B.; Iten, M.; Grether-Bühler, Y.; Kaminsky, R.; Brun, R. The Alamar Blue assay to determine drug sensitivity of African trypanosomes (T.b. rhodesiense and T.b. gambiense) in vitro. Acta Trop. 1997, 68, 139–147. [Google Scholar] [CrossRef]
- Huber, W.; Koella, J.C. A comparison of three methods of estimating EC50 in studies of drug resistance of malaria parasites. Acta Trop. 1993, 55, 257–261. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Gogal, R.M.; Walsh, J.E. A new rapid and simple non-radioactive assay to monitor and determine the proliferation of lymphocytes: An alternative to [3H]thymidine incorporation assay. J. Immunol. Methods 1994, 170, 211–224. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | L. donovani | P. falciparum | T. b. rhodesiense | L6 Cells | |||
|---|---|---|---|---|---|---|---|
| No. | IC50 a (µM) | SI b | IC50 a (µM) | SI b | IC50 a (µM) | SI b | |
| 1 | 22.6 ± 6.5 | 4.2 | 53.1 ± 2.0 | 1.8 | 59.4 ± 2.5 | 1.6 | 95.2 ± 9.3 |
| 2 | 4.5 ± 1.0 | 25.5 | 9.5 ± 0.1 | 12.1 | 48.3 ± 1.3 | 2.4 | 115.0 ± 2.6 |
| 3 | 36.0 ± 7.3 | 4.7 | 36.1 ± 1.0 | 4.7 | 47.5 ± 4.5 | 3.6 | 2001 ± 47.0 |
| 4 | 29.2 ± 6.0 | 4.8 | 17.4 ± 2.1 | 8.0 | 40.7 ± 0.4 | 3.4 | 163.0 ± 4.5 |
| 5 | 141.3 ± 1.9 | n.d | 158.7 | n.d | 206.0 ± 66.7 | n.d | ˃317.5 |
| 6 | 99.4 ± 9.3 | 2.5 | 68.4 ± 3.8 | 3.6 | 120.9 ± 36.9 | 2.0 | 246.8 ± 3.7 |
| 7 | 136.5 ± 1.5 | n.d | 48.1 ± 10.9 | n.d | 114.0 ± 0.0 | n.d | ˃280.9 |
| 8 | 97.0 ± 0.9 | 1.5 | 30.6 ± 8.3 | 4.8 | 18.9 ± 8.4 | 7.8 | 146.5 ± 18.8 |
| 9 | 69.6 ± 4.6 | 3.0 | 30.4 ± 8.0 | 6.8 | 72.7 ± 30.4 | 2.9 | 207.8 ± 21.6 |
| 10 | 55.3 ± 3.0 | 2.3 | 9.3 ± 1.3 | 13.7 | 27.5 ± 8.4 | 4.6 | 127.6 ± 3.9 |
| Positive control | 0.5 d | 0.01 e | 0.01 c | 0.03 f | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud, A.B.; Danton, O.; Kaiser, M.; Han, S.; Moreno, A.; Abd Algaffar, S.; Khalid, S.; Oh, W.K.; Hamburger, M.; Mäser, P. Lignans, Amides, and Saponins from Haplophyllum tuberculatum and Their Antiprotozoal Activity. Molecules 2020, 25, 2825. https://doi.org/10.3390/molecules25122825
Mahmoud AB, Danton O, Kaiser M, Han S, Moreno A, Abd Algaffar S, Khalid S, Oh WK, Hamburger M, Mäser P. Lignans, Amides, and Saponins from Haplophyllum tuberculatum and Their Antiprotozoal Activity. Molecules. 2020; 25(12):2825. https://doi.org/10.3390/molecules25122825
Chicago/Turabian StyleMahmoud, Abdelhalim Babiker, Ombeline Danton, Marcel Kaiser, Sohee Han, Aitor Moreno, Shereen Abd Algaffar, Sami Khalid, Won Keun Oh, Matthias Hamburger, and Pascal Mäser. 2020. "Lignans, Amides, and Saponins from Haplophyllum tuberculatum and Their Antiprotozoal Activity" Molecules 25, no. 12: 2825. https://doi.org/10.3390/molecules25122825
APA StyleMahmoud, A. B., Danton, O., Kaiser, M., Han, S., Moreno, A., Abd Algaffar, S., Khalid, S., Oh, W. K., Hamburger, M., & Mäser, P. (2020). Lignans, Amides, and Saponins from Haplophyllum tuberculatum and Their Antiprotozoal Activity. Molecules, 25(12), 2825. https://doi.org/10.3390/molecules25122825