White Matter Brain Network Research in Alzheimer’s Disease Using Persistent Features


Abstract
1. Introduction
2. Results
2.1. Demographic Information
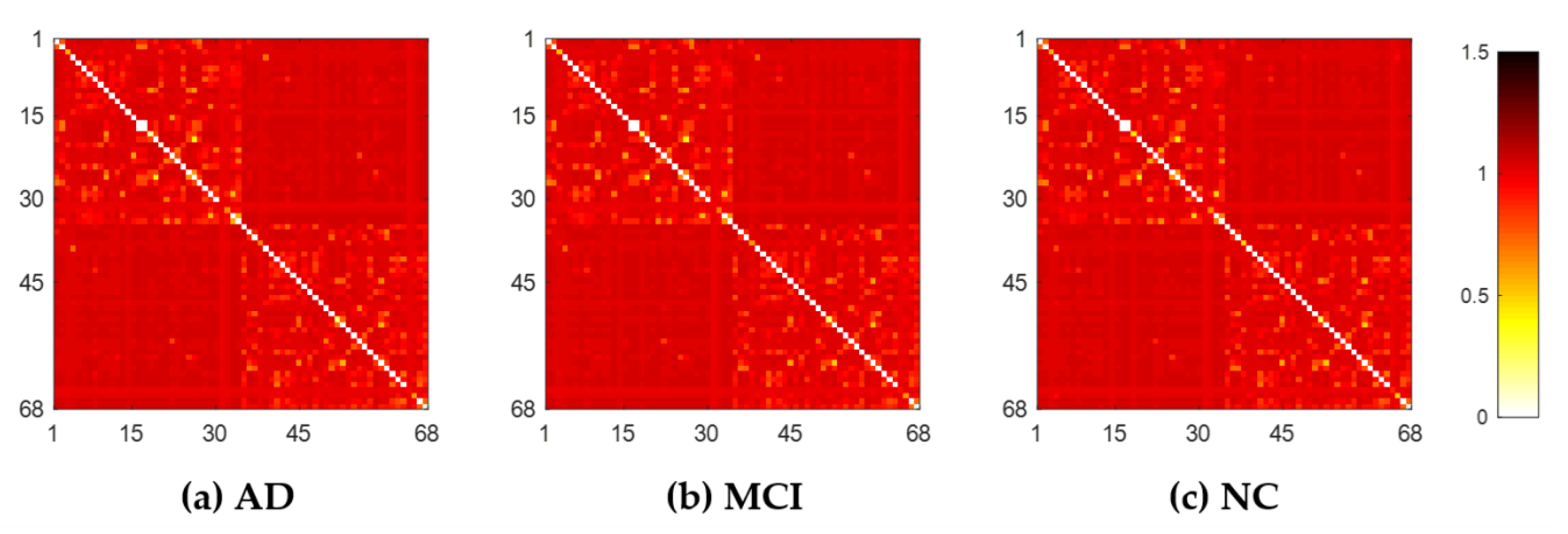
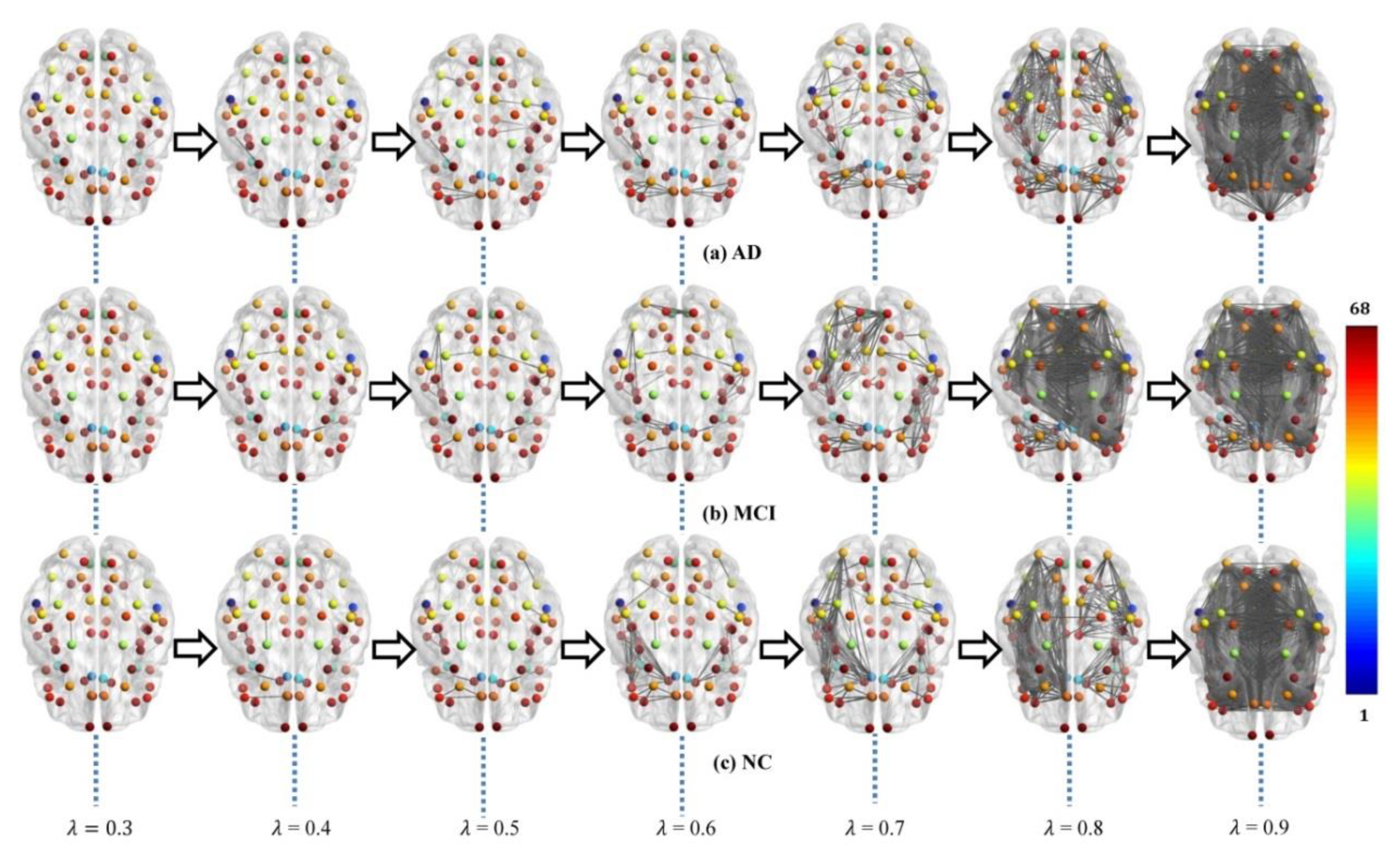
2.2. WM Brain Network
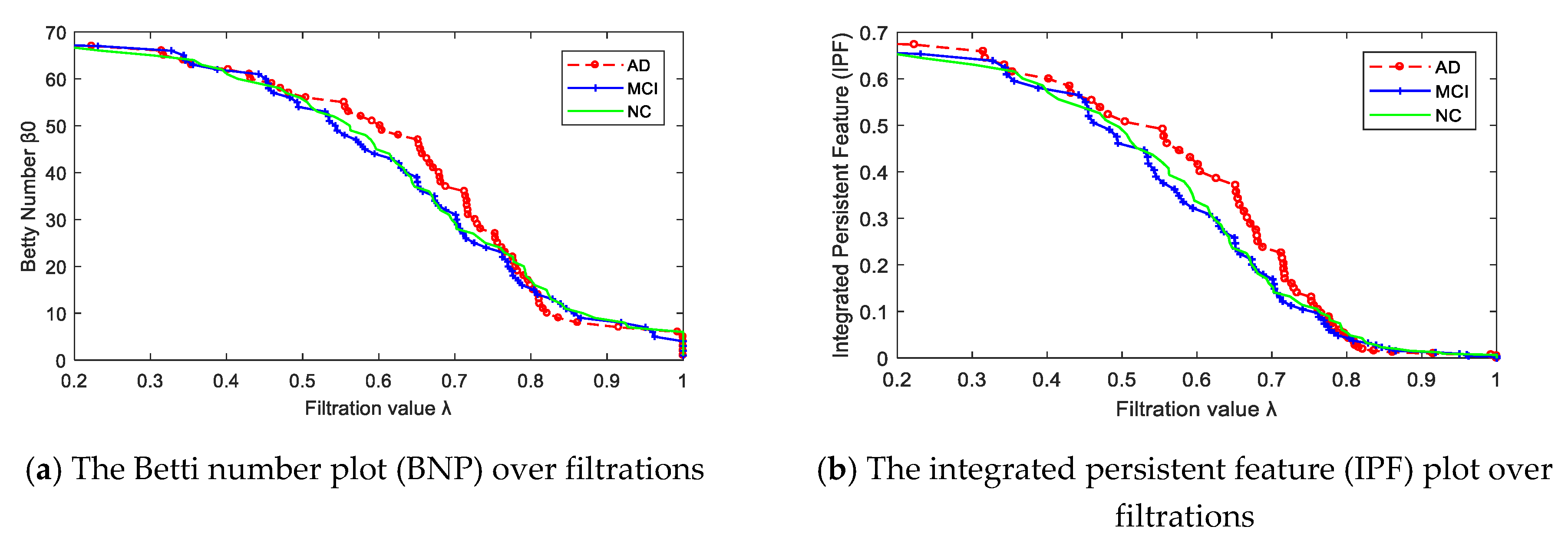
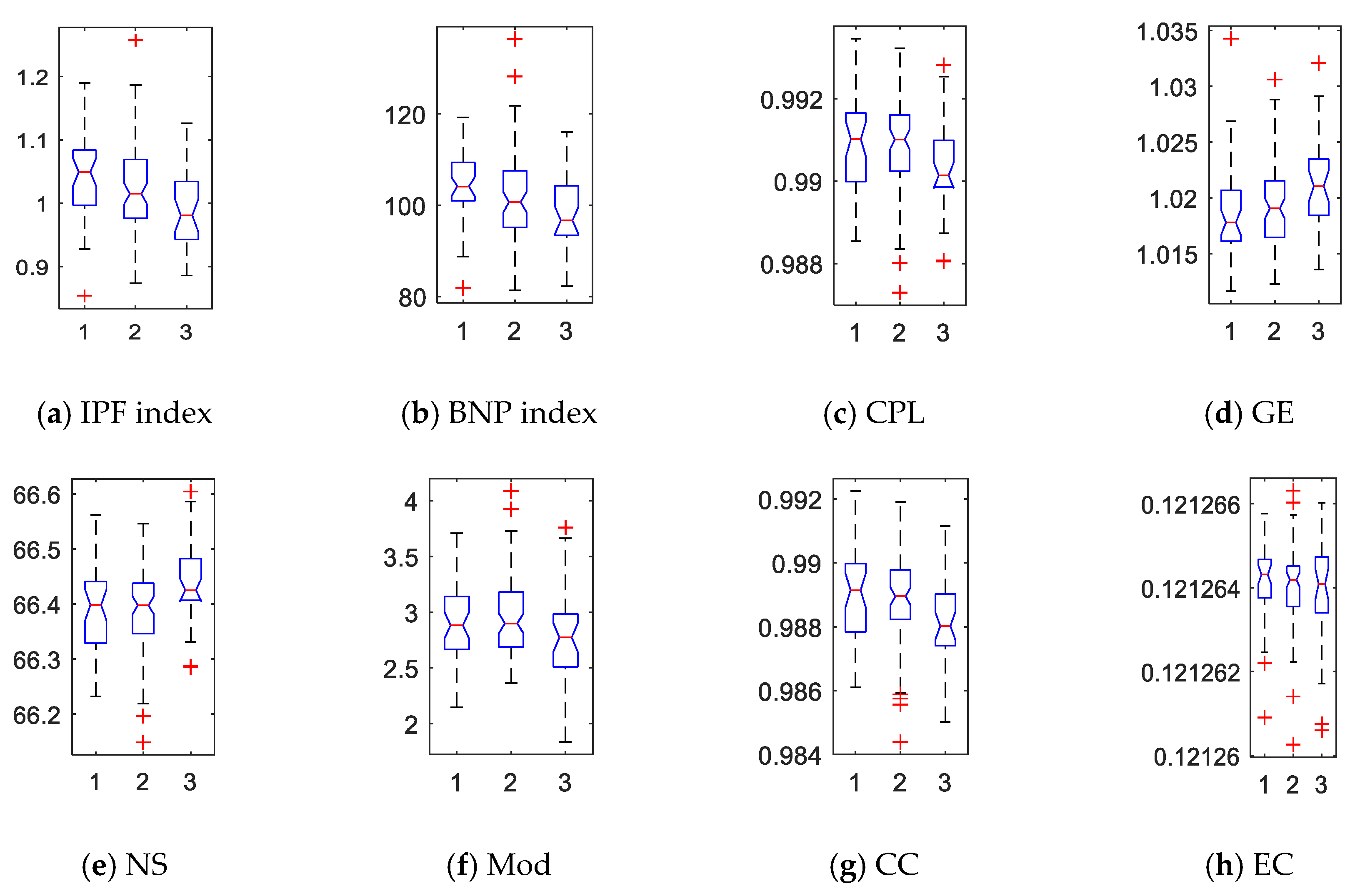
2.3. Network Properties
2.4. Statistical Group Difference Performance
2.5. Main Findings
3. Discussion
3.1. Validation on Various Parcellations
3.2. Exploring Other Connectivity Definitions
3.3. Limitations and Future Work
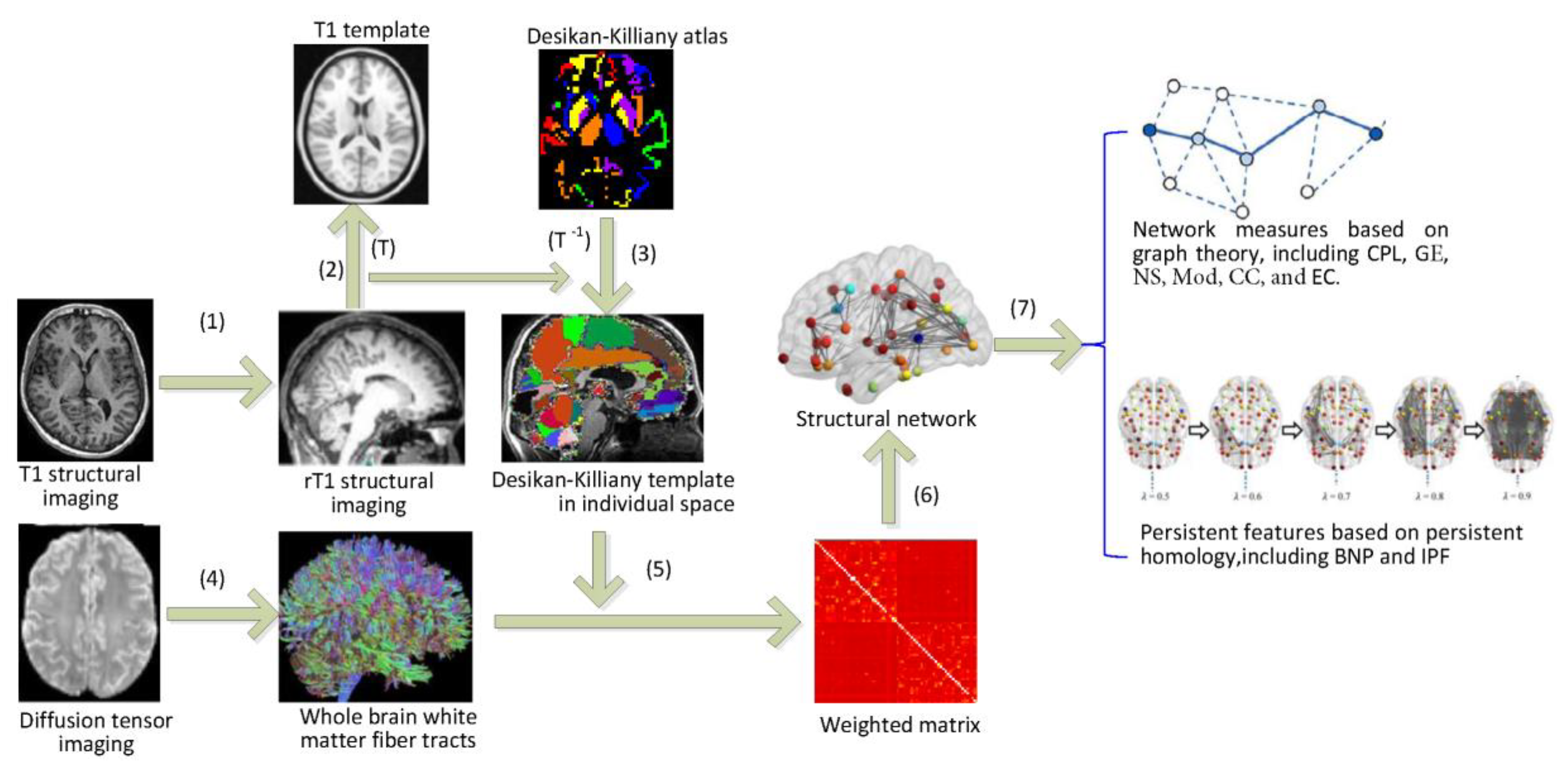
4. Materials and Methods
4.1. Subjects and Data Preprocessing
4.2. Network Construction
- (1)
- Firstly, the T1 image of each subject is registered to its b0 image to obtain the rT1 image in each subject space.
- (2)
- Secondly, the rT1 image in the individual space is registered to the T1 template of ICBM-DTI-152 in the MNI space, and the spatial transformation parameter T is obtained.
- (3)
- The Desikan–Killiany template in the MNI space is converted into the individual subject space using inverse transform parameter T−1.
- (4)
- Probabilistic fiber tracking [45] was performed to obtain the white matter fiber tracts in the whole brain tissue of each subject. Each pair of brain regions is assessed using 5000 times of probability tractography and the number of traces that reach both source and target regions is regarded as the connection between them.
- (5)
- We then calculate the weighted matrix W (68 × 68) where each element measures the similarity of the probability fiber connection patterns between each pair of the brain regions [43]. The edge weight is defined as 1 minus Pearson correlation of fiber connections between them, i.e.,where are the fiber connections of the i-th and j-th brain region to other regions respectively, cov is the covariance, is the standard deviation, and is the coefficient of the Pearson correlation.
- (6)
- For each individual, a weighted matrix W is treated as a WM brain structural network. The connectivity ranges from 0 to 2 whose value closer to 0 means stronger relationship between a pair of brain regions.
4.3. Network Indices
4.3.1. Graph Theoretical Measures
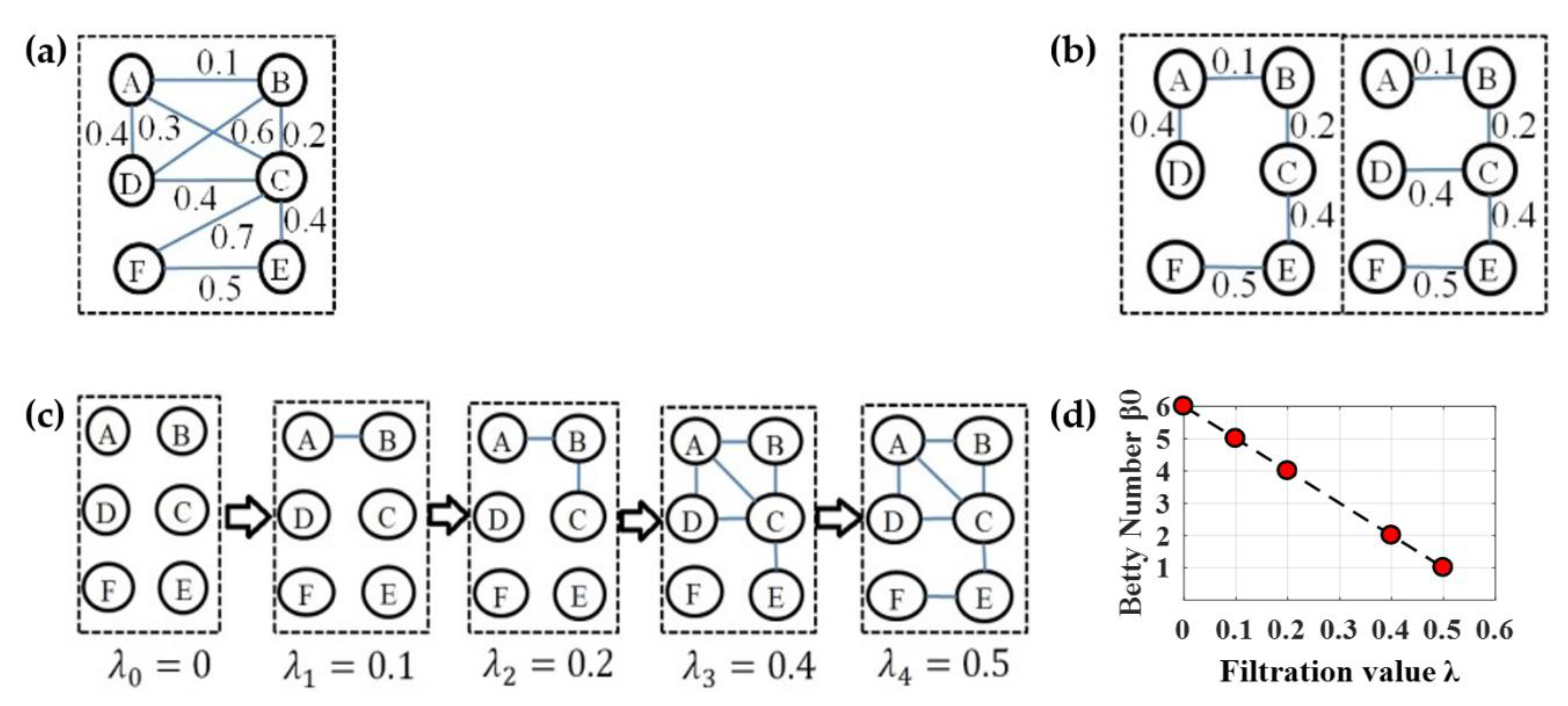
4.3.2. Persistent Features
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scheltens, P.; Blennow, K.; Breteler, M.M.B.; De Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. Lancet 2016, 388, 505–517. [Google Scholar] [CrossRef]
- Wu, Z.; Xu, D.; Potter, T.; Zhang, Y.; Alzheimers Dis Neuroimaging, I. Effects of brain parcellation on the characterization of topological deterioration in alzheimer’s disease. Front. Aging Neurosci. 2019, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Aisen, P.S.; DuBois, B.; Froelich, L.; Jack, C.R., Jr.; Jones, R.W.; Morris, J.C.; Raskin, J.; Dowsett, S.A.; Scheltens, P. Drug development in alzheimer’s disease: The path to 2025. Alzheimers Res. Ther. 2016, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Sporns, O.; Tononi, G.; Kötter, R. The human connectome: A structural description of the human brain. PLoS Comput. Biol. 2005, 1, e42. [Google Scholar] [CrossRef]
- Canter, R.G.; Penney, J.; Tsai, L.-H. The road to restoring neural circuits for the treatment of Alzheimer’s disease. Nature 2016, 539, 187–196. [Google Scholar] [CrossRef]
- Frere, S.; Slutsky, I. Alzheimer’s disease: From firing instability to homeostasis network collapse. Neuron 2018, 97, 32–58. [Google Scholar] [CrossRef]
- Palop, J.J.; Mucke, L. Network abnormalities and interneuron dysfunction in Alzheimer disease. Nat. Rev. Neurosci. 2016, 17, 777–792. [Google Scholar] [CrossRef]
- Tohda, C. New age therapy for Alzheimer’s disease by neuronal network reconstruction. Biol. Pharm. Bull. 2016, 39, 1569–1575. [Google Scholar] [CrossRef]
- Nir, T.M.; Villalon-Reina, J.E.; Prasad, G.; Jahanshad, N.; Joshi, S.H.; Toga, A.W.; Bernstein, M.A.; Jack, C.R.; Weiner, M.W.; Thompson, P.M.; et al. Diffusion weighted imaging-based maximum density path analysis and classification of Alzheimer’s disease. Neurobiol. Aging 2015, 36, S132–S140. [Google Scholar] [CrossRef]
- Rose, S.E.; Chen, F.; Chalk, J.B.; Zelaya, F.O.; Strugnell, W.E.; Benson, M.; Semple, J.; Doddrell, D.M. Loss of connectivity in Alzheimer’s disease: An evaluation of white matter tract integrity with colour coded mr diffusion tensor imaging. J. Neurol. Neurosurg. Psychiatry 2000, 69, 528–530. [Google Scholar] [CrossRef]
- Sporns, O. Graph theory methods: Applications in brain networks. Dialogues Clin. Neurosci. 2018, 20, 111–121. [Google Scholar]
- Pievani, M.; De Haan, W.; Wu, T.; Seeley, W.W.; Frisoni, G.B. Functional network disruption in the degenerative dementias. Lancet Neurol. 2011, 10, 829–843. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. NeuroImage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Qiu, T.; Luo, X.; Shen, Z.; Huang, P.; Xu, X.; Zhou, J.; Zhang, M. Disrupted brain network in progressive mild cognitive impairment measured by eigenvector centrality mapping is linked to cognition and cerebrospinal fluid biomarkers. J. Alzheimers Dis. 2016, 54, 1483–1493. [Google Scholar] [CrossRef] [PubMed]
- Daianu, M.; Jahanshad, N.; Nir, T.M.; Jack, C.R.; Weiner, M.W.; Bernstein, M.A.; Thompson, P.M.; Alzheimer’s Dis, N. Rich club analysis in the alzheimer’s disease connectome reveals a relatively undisturbed structural core network. Hum. Brain Mapp. 2015, 36, 3087–3103. [Google Scholar] [CrossRef]
- Van Wijk, B.C.M.; Stam, C.J.; Daffertshofer, A. Comparing brain networks of different size and connectivity density using graph theory. PLoS ONE 2010, 5, e13701. [Google Scholar] [CrossRef]
- Drakesmith, M.; Caeyenberghs, K.; Dutt, A.; Lewis, G.; David, A.S.; Jones, D.K. Overcoming the effects of false positives and threshold bias in graph theoretical analyses of neuroimaging data. NeuroImage 2015, 118, 313–333. [Google Scholar] [CrossRef]
- Edelsbrunner, H.; Harer, J. Computational Topology: An Introduction; American Mathematical Society: Providence, RI, USA, 2010. [Google Scholar]
- Giusti, C.; Ghrist, R.; Bassett, D.S. Two’s company, three (or more) is a simplex: Algebraic-topological tools for understanding higher-order structure in neural data. J. Comput. Neurosci. 2016, 41, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Leon, P.; Knock, S.A.; Spiegler, A.; Jirsa, V.K. Mathematical framework for large-scale brain network modeling in the virtual brain. NeuroImage 2015, 111, 385–430. [Google Scholar] [CrossRef]
- Deco, G.; Jirsa, V.K.; McIntosh, A.R. Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat. Rev. Neurosci. 2011, 12, 43–56. [Google Scholar] [CrossRef]
- Lee, H.; Kang, H.; Chung, M.K.; Kim, B.-N.; Lee, D.S. Persistent brain network homology from the perspective of dendrogram. IEEE Trans. Med. Imaging 2012, 31, 2267–2277. [Google Scholar]
- Choi, H.; Kim, Y.K.; Kang, H.; Lee, H.; Im, H.-J.; Kim, E.E.; Chung, J.-K.; Lee, D.S. Abnormal metabolic connectivity in the pilocarpine-induced epilepsy rat model: A multiscale network analysis based on persistent homology. NeuroImage 2014, 99, 226–236. [Google Scholar] [CrossRef]
- Lee, H.; Kang, H.; Chung, M.K.; Lim, S.; Kim, B.N.; Lee, D.S. Integrated multimodal network approach to pet and mri based on multidimensional persistent homology. Hum. Brain Mapp. 2017, 38, 1387–1402. [Google Scholar] [CrossRef]
- Chung, M.K.; Hanson, J.L.; Ye, J.; Davidson, R.J.; Pollak, S.D. Persistent homology in sparse regression and its application to brain morphometry. IEEE Trans. Med. Imaging 2015, 34, 1928–1939. [Google Scholar] [CrossRef]
- Kuang, L.; Han, X.; Chen, K.; Caselli, R.J.; Reiman, E.M.; Wang, Y.; Weiner, M.W.; Aisen, P.; Weiner, M.; Petersen, R.; et al. A concise and persistent feature to study brain resting-state network dynamics: Findings from the Alzheimer’s disease neuroimaging initiative. Hum. Brain Mapp. 2019, 40, 1062–1081. [Google Scholar] [CrossRef]
- Jack, C.R.; Bernstein, M.A.; Fox, N.C.; Thompson, P.; Alexander, G.; Harvey, D.; Borowski, B.; Britson, P.J.; Whitwell, J.L.; Ward, C. The Alzheimer’s disease neuroimaging initiative (ADNI): MRI methods. J. Magn. Reson. Imaging 2008, 27, 685–691. [Google Scholar] [CrossRef]
- Hollander, M.; Wolfe, D.A.; Chicken, E. Nonparametric Statistical Methods; John Wiley & Sons: New York, NY, USA, 1973. [Google Scholar]
- Morris, J.C. The clinical dementia rating (CDR): Current version and scoring rules. Neurology 1993, 43, 2412–2414. [Google Scholar] [CrossRef] [PubMed]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the national institute on aging-Alzheimer’s association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimer’s Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.S.; Segonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on mri scans into gyral based regions of interest. NeuroImage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- Dai, Z.; He, Y. Disrupted structural and functional brain connectomes in mild cognitive impairment and alzheimer’s disease. Neurosci. Bull. 2014, 30, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.B.; Van Westen, D.; Stomrud, E.; Strandberg, T.O.; Volpe, G.; Westman, E.; Hansson, O. Abnormal structural brain connectome in individuals with preclinical Alzheimer’s disease. Cereb. Cortex 2018, 28, 3638–3649. [Google Scholar] [CrossRef] [PubMed]
- Kuang, L.; Zhao, D.; Xing, J.; Chen, Z.; Xiong, F.; Han, X. Metabolic brain network analysis of FDG-PET in Alzheimer’s disease using kernel-based persistent features. Molecules 2019, 24, 2301. [Google Scholar] [CrossRef] [PubMed]
- Martensson, G.; Pereira, J.B.; Mecocci, P.; Vellas, B.; Tsolaki, M.; Kloszewska, I.; Soininen, H.; Lovestone, S.; Simmons, A.; Volpe, G.; et al. Stability of graph theoretical measures in structural brain networks in Alzheimer’s disease. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fischl, B.; Salat, D.H.; Busa, E.; Albert, M.; Dieterich, M.; Haselgrove, C.; Van der Kouwe, A.; Killiany, R.; Kennedy, D.; Klaveness, S.; et al. Whole brain segmentation: Automated labeling of neuroanatomical structures in the human brain. Neuron 2002, 33, 341–355. [Google Scholar] [CrossRef]
- Korgaonkar, M.S.; Fornito, A.; Williams, L.M.; Grieve, S.M. Abnormal structural networks characterize major depressive disorder: A connectome analysis. Biol. Psychiatry 2014, 76, 567–574. [Google Scholar] [CrossRef]
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. NeuroImage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Proix, T.; Spiegler, A.; Schirner, M.; Rothmeier, S.; Ritter, P.; Jirsa, V.K. How do parcellation size and short-range connectivity affect dynamics in large-scale brain network models? NeuroImage 2016, 142, 135–149. [Google Scholar] [CrossRef]
- Zalesky, A.; Fornito, A.; Harding, I.H.; Cocchi, L.; Yucel, M.; Pantelis, C.; Bullmore, E.T. Whole-brain anatomical networks: Does the choice of nodes matter? NeuroImage 2010, 50, 970–983. [Google Scholar] [CrossRef]
- Schirner, M.; Rothmeier, S.; Jirsa, V.K.; McIntosh, A.R.; Ritter, P. An automated pipeline for constructing personalized virtual brains from multimodal neuroimaging data. NeuroImage 2015, 117, 343–357. [Google Scholar] [CrossRef]
- Wee, C.Y.; Yap, P.T.; Li, W.B.; Denny, K.; Browndyke, J.N.; Potter, G.G.; Welsh-Bohmer, K.A.; Wang, L.H.; Shen, D.G. Enriched white matter connectivity networks for accurate identification of MCI patients. NeuroImage 2011, 54, 1812–1822. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, J.; Yu, J.; Zhan, L.; Thompson, P.M.; Wang, Y. Enhancing diffusion MRI measures by integrating grey and white matter morphometry with hyperbolic wasserstein distance. Proc. IEEE Int. Symp. Biomed. Imaging 2017, 2017, 520–524. [Google Scholar] [PubMed]
- Petkoski, S.; Palva, J.M.; Jirsa, V.K. Phase-lags in large scale brain synchronization: Methodological considerations and in-silico analysis. PLoS Comput. Biol. 2018, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Shu, N.; An, L.; Wang, P.; Sun, L.; Xia, M.-R.; Wang, J.-H.; Gong, G.-L.; Zang, Y.-F.; Wang, Y.-F.; et al. Probabilistic diffusion tractography and graph theory analysis reveal abnormal white matter structural connectivity networks in drug-naive boys with attention deficit/hyperactivity disorder. J. Neurosci. 2013, 33, 10676–10687. [Google Scholar] [CrossRef] [PubMed]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Wang, W.; Yang, L.; Chen, K.; Chen, R.; Han, Y. Rich club disturbances of the human connectome from subjective cognitive decline to Alzheimer’s disease. Theranostics 2018, 8, 3237–3255. [Google Scholar] [CrossRef]
- De Pasquale, F.; Della Penna, S.; Sporns, O.; Romani, G.; Corbetta, M. A dynamic core network and global efficiency in the resting human brain. Cereb. Cortex 2016, 26, 4015–4033. [Google Scholar] [CrossRef]
- Watts, D.J.; Strogatz, S.H. Collective dynamics of ‘small-world’ networks. Nature 1998, 393, 440. [Google Scholar] [CrossRef]
- Wang, J.; Zuo, X.; Dai, Z.; Xia, M.; Zhao, Z.; Zhao, X.; Jia, J.; Han, Y.; He, Y. Disrupted functional brain connectome in individuals at risk for Alzheimer’s disease. Biol. Psychiatry 2013, 73, 472–481. [Google Scholar] [CrossRef]
- Maria, C.; Boissonnat, J.-D.; Glisse, M.; Yvinec, M. The gudhi library: Simplicial complexes and persistent homology. In Proceedings of the 4th International Congress on Mathematical Software (ICMS 2014), Seoul, Korea, 5–9 August 2014; pp. 167–174. [Google Scholar]
- Zomorodian, A.; Carlsson, G. Computing persistent homology. Discret. Comput. Geom. 2005, 33, 249–274. [Google Scholar] [CrossRef]
- Carlsson, G. Topology and data. Bull. Am. Math. Soc. 2009, 46, 255–308. [Google Scholar] [CrossRef]
- Lee, H.; Chung, M.K.; Kang, H.; Kim, B.-N.; Lee, D.S. Computing the shape of brain networks using graph filtration and gromov-hausdorff metric. In Proceedings of the 14th International Conference on Medical Image Computing and Computer Assisted Intervention (MICCAI 2011), Toronto, ON, Canada, 18–22 September 2011; pp. 302–309. [Google Scholar]
Sample Availability: Samples are available from the ADNI (adni.loni.usc.edu). |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AD (n = 40) | MCI (n = 77) | NC (n = 33) | p-Value | |
|---|---|---|---|---|
| Male/Female | 22/18 | 42/35 | 18/15 | 0.999 |
| Age | 72.45 ± 5.60 | 74.22 ± 6.73 | 74.18 ± 8.28 | 0.388 |
| Education | 16.1 ± 2.72 | 15.57 ± 2.67 | 15.45 ± 2.81 | 0.443 |
| CDR | ≥1 | 0.5 | 0 | 0 |
| IPF | BNP | CPL | GE | NS | Mod | CC | EC | |
|---|---|---|---|---|---|---|---|---|
| AD vs. MCI | 0.084 | 0.095 | 0.355 | 0.235 | 0.372 | 0.322 | 0.308 | 0.234 |
| AD vs. NC | 0.0003 | 0.001 | 0.011 | 0.007 | 0.011 | 0.016 | 0.007 | 0.049 |
| MCI vs. NC | 0.007 | 0.015 | 0.014 | 0.021 | 0.012 | 0.024 | 0.011 | 0.113 |
| AD vs. MCI vs. NC | 0.002 | 0.003 | 0.020 | 0.016 | 0.020 | 0.046 | 0.014 | 0.470 |
| Parcellation | IPF | BNP | CPL | GE | NS | Mod | CC | EC |
|---|---|---|---|---|---|---|---|---|
| Differences on DK84 Atlas | ||||||||
| AD vs. MCI | ↑c | ↑c | ns | ns | ns | ns | ns | ns |
| AD vs. NC | ↑a | ↑a | ↑a | ↓a | ↓a | ↑b | ↑a | ↑b |
| MCI vs. NC | ↑a | ↑a | ↑a | ↓a | ↓a | ↑b | ↑a | ↑b |
| Differences on AAL90 Atlas | ||||||||
| AD vs. MCI | ↑b | ↑c | ns | ns | ↓c | ↑c | ns | ns |
| AD vs. NC | ↑a | ↑a | ↑b | ↓b | ↓a | ↑b | ↓b | ↑b |
| MCI vs. NC | ↑a | ↑a | ↑b | ↓b | ↓b | ↑b | ↓c | ↑b |
| Differences on DK272 Atlas | ||||||||
| AD vs. MCI | ↑b | ↑c | ↑c | ↓c | ns | ns | ns | ns |
| AD vs. NC | ↑a | ↑a | ↑b | ↓b | ↓a | ↑b | ↑c | ↑b |
| MCI vs. NC | ↑b | ↑b | ↑b | ↓b | ↓b | ↑c | ↑b | ↑c |
| Between-Group | IPF | BNP | CPL | GE | NS | Mod | CC | EC |
|---|---|---|---|---|---|---|---|---|
| FA-based connectivity | ||||||||
| AD vs. MCI | 0.056 | 0.073 | 0.121 | 0.336 | 0.231 | 0.255 | 0.457 | 0.436 |
| AD vs. NC | 0.009 | 0.013 | 0.032 | 0.016 | 0.028 | 0.021 | 0.047 | 0.058 |
| MCI vs. NC | 0.013 | 0.022 | 0.044 | 0.042 | 0.025 | 0.022 | 0.037 | 0.086 |
| Spearman Correlation | ||||||||
| AD vs. MCI | 0.382 | 0.246 | 0.038 | 0.291 | 0.036 | 0.163 | 0.291 | 0.046 |
| AD vs. NC | 0.048 | 0.460 | 0.005 | 0.225 | 0.004 | 0.054 | 0.307 | 0.009 |
| MCI vs. NC | 0.026 | 0.302 | 0.114 | 0.225 | 0.111 | 0.184 | 0.138 | 0.147 |
| Partial Correlation | ||||||||
| AD vs. MCI | 0.320 | 0.483 | 0.406 | 0.255 | 0.394 | 0.433 | 0.023 | 0.247 |
| AD vs. NC | 0.022 | 0.045 | 0.061 | 0.065 | 0.063 | 0.373 | 0.141 | 0.064 |
| MCI vs. NC | 0.030 | 0.022 | 0.068 | 0.143 | 0.069 | 0.421 | 0.306 | 0.139 |
| Absolute value of Pearson correlation | ||||||||
| AD vs. MCI | 0.072 | 0.083 | 0.376 | 0.287 | 0.387 | 0.058 | 0.321 | 0.090 |
| AD vs. NC | 0.008 | 0.015 | 0.012 | 0.015 | 0.013 | 0.020 | 0.045 | 0.120 |
| MCI vs. NC | 0.012 | 0.024 | 0.013 | 0.043 | 0.012 | 0.388 | 0.071 | 0.235 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuang, L.; Gao, Y.; Chen, Z.; Xing, J.; Xiong, F.; Han, X. White Matter Brain Network Research in Alzheimer’s Disease Using Persistent Features. Molecules 2020, 25, 2472. https://doi.org/10.3390/molecules25112472
Kuang L, Gao Y, Chen Z, Xing J, Xiong F, Han X. White Matter Brain Network Research in Alzheimer’s Disease Using Persistent Features. Molecules. 2020; 25(11):2472. https://doi.org/10.3390/molecules25112472
Chicago/Turabian StyleKuang, Liqun, Yan Gao, Zhongyu Chen, Jiacheng Xing, Fengguang Xiong, and Xie Han. 2020. "White Matter Brain Network Research in Alzheimer’s Disease Using Persistent Features" Molecules 25, no. 11: 2472. https://doi.org/10.3390/molecules25112472
APA StyleKuang, L., Gao, Y., Chen, Z., Xing, J., Xiong, F., & Han, X. (2020). White Matter Brain Network Research in Alzheimer’s Disease Using Persistent Features. Molecules, 25(11), 2472. https://doi.org/10.3390/molecules25112472