
Nutrient and Sensory Metabolites Profiling of Averrhoa Carambola L. (Starfruit) in the Context of Its Origin and Ripening Stage by GC/MS and Chemometric Analysis

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Primary Metabolites Profiling Viz. Sugars, Amino and Organic Acids via GC-MS Post Silylation
2.1.1. Sugars
2.1.2. Polyols
2.1.3. Organic Acids
2.1.4. Amino Acids
2.1.5. Alcohols
2.1.6. Fatty Acids/Hydrocarbons and Sterols
2.1.7. Nitrogenous Compounds
2.1.8. Aromatics
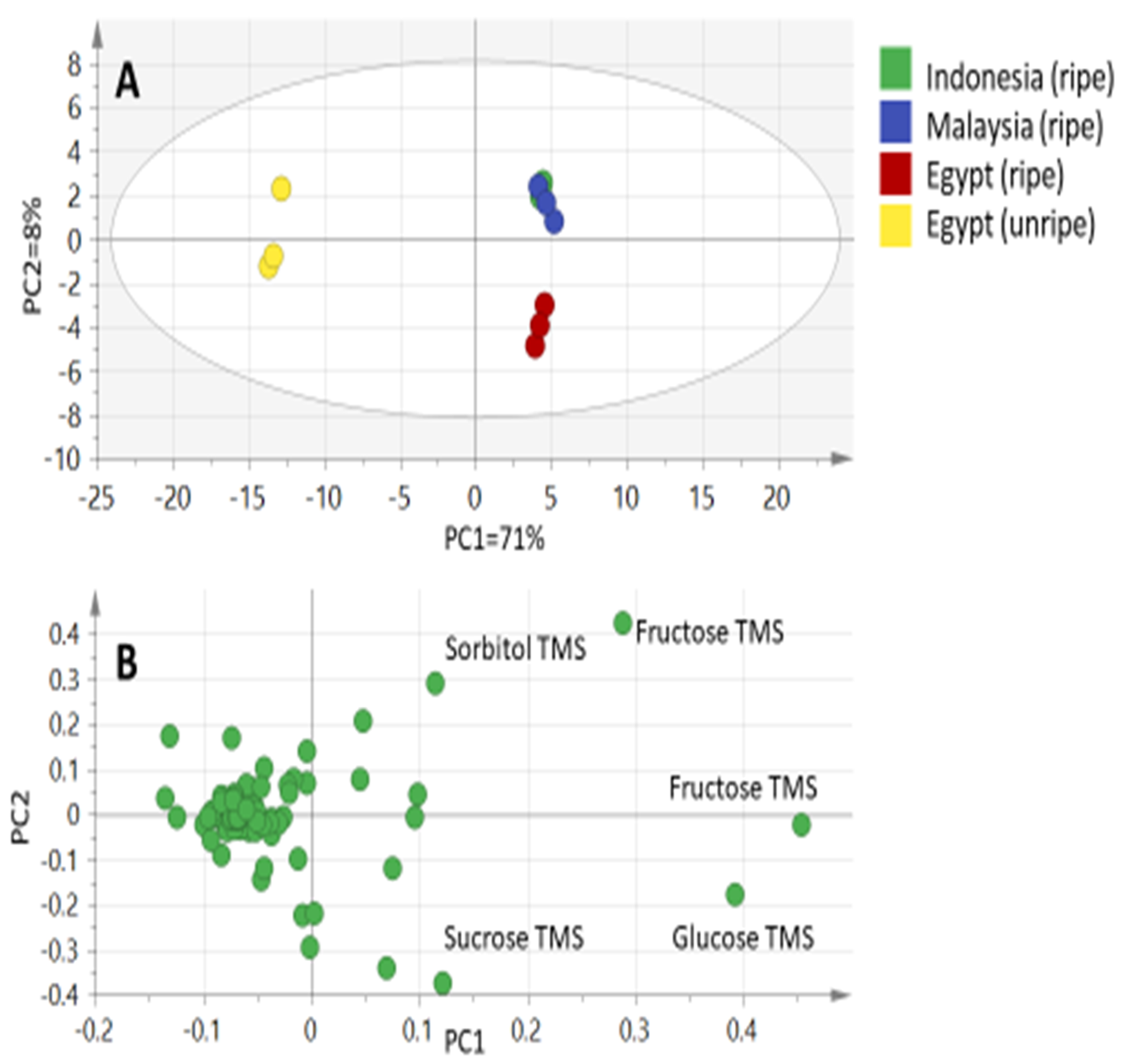
2.2. Mutlivariate Data Analyses of Primary Fruit Metabolites by GC-MS
2.3. Volatiles Profiling via Headspace SPME Coupled to GC-MS
2.3.1. Esters
2.3.2. Aldehydes
2.3.3. Ethers/oxides
2.3.4. Ketones
2.4. Quantitative Determination of Major Silylated Primary Metabolites
2.4.1. Quantitative Determination of Major Detected Sugars Viz. Fructose and Glucose
2.4.2. Quantitative Determination of Oxalic Acid
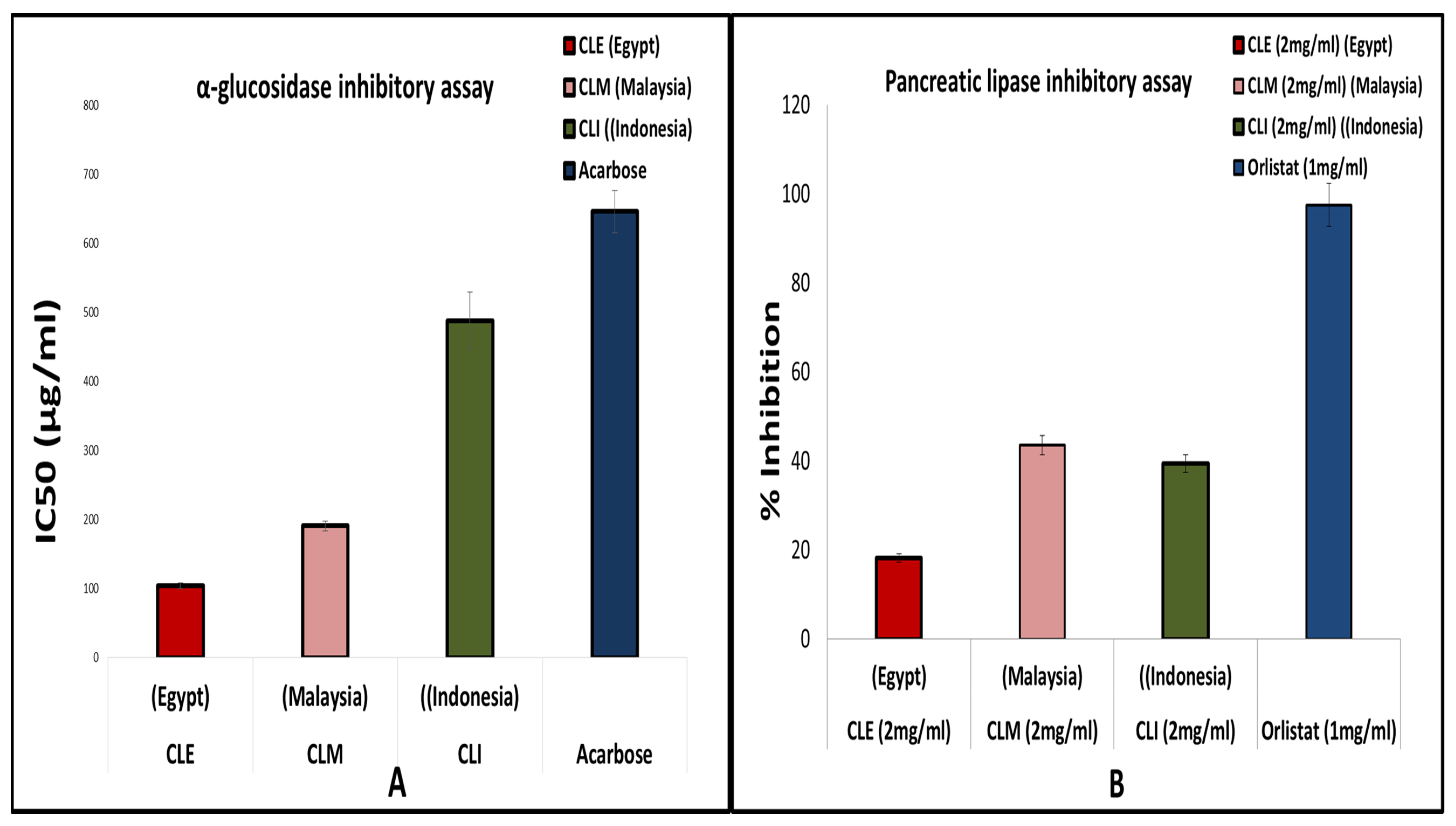
2.5. Enzyme Inhibition Assays
2.5.1. α-Glucosidase Inhibitory Assay
2.5.2. Pancreatic Lipase Inhibitory Assay
3. Materials and Methods
3.1. Plant, SPME, and Chemicals
3.2. SPME Volatiles Isolation from Whole Fruit
3.3. GC-MS Volatile Analysis
3.4. GC-MS Analysis of Silylated Primary Metabolites
3.5. Quantitative Determination of Major Silylated Primary Metabolites Detected by GC-MS Analysis
3.5.1. Quantitative Determination of Oxalic Acid
3.5.2. Quantitative Determination of Major Detected SugarsGlucose and Fructose
3.6. GC-MS Multivariate Data Analyses
3.7. Enzyme Inhibition Assays
3.7.1. α-Glucosidase Inhibitory Assay
3.7.2. Pancreatic Lipase Inhibitory Assay
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Farag, M.A.; El Senousy, A.S.; El-Ahmady, S.H.; Porzel, A.; Wessjohann, L.A. Comparative metabolome-based classification of Senna drugs: A prospect for phyto-equivalency of its different commercial products. Metabolomics 2019, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ribes-Moya, A.M.; Raigón, M.D.; Moreno-Peris, E.; Fita, A.; Rodríguez-Burruezo, A. Response to organic cultivation of heirloom Capsicum peppers: Variation in the level of bioactive compounds and effect of ripening. PLoS ONE 2018, 13, e0207888. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Xie, H.; Jia, X.; Wei, X. Flavonoid C-glycosides from star fruit and their antioxidant activity. J. Funct. Foods 2015, 16, 204–210. [Google Scholar] [CrossRef]
- Verma, S.; Dhaneshwar, S.; Ramana, M.V.; Rawat, A.K.S. Gas chromatography–mass spectrometry and high-performance thin-layer chromatography quantifications of some physiologically active secondary metabolites in Averrhoa carambola L. fruits. JPC J. Planar Chromatogr. Mod. TLC 2018, 31, 207–212. [Google Scholar] [CrossRef]
- Jia, X.; Yang, D.; Yang, Y.; Xie, H. Carotenoid-Derived flavor precursors from Averrhoa carambola fresh fruit. Molecules 2019, 24, 256. [Google Scholar] [CrossRef]
- Guevara, M.; Tejera, E.; Granda-Albuja, M.G.; Iturralde, G.; Chisaguano-Tonato, M.; Granda-Albuja, S.; Jaramillo-Vivanco, T.; Giampieri, F.; Battino, M.; Alvarez-Suarez, J.M. Chemical composition and antioxidant activity of the main fruits consumed in the western coastal region of Ecuador as a source of health-promoting compounds. Antioxidants 2019, 8, 387. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Yang, D.; Xie, H.; Jiang, Y.; Wei, X. Non-flavonoid phenolics from Averrhoa carambola fresh fruit. J. Funct. Foods 2017, 32, 419–425. [Google Scholar] [CrossRef]
- Yan, S.W.; Ramasamy, R.; Alitheen, N.B.M.; Rahmat, A. A comparative assessment of nutritional composition, total phenolic, total flavonoid, antioxidant capacity, and antioxidant vitamins of two types of malaysian underutilized fruits (Averrhoa bilimbi and Averrhoa carambola). Int. J. Food Prop. 2013, 16, 1231–1244. [Google Scholar] [CrossRef]
- Yang, D.; Jia, X.; Xie, H. Heptyl vicianoside and methyl caramboside from sour star fruit. Nat. Prod. Res. 2018, 33, 1233–1236. [Google Scholar] [CrossRef]
- Bhat, R.; Ameran, S.B.; Voon, H.C.; Karim, A.A.; Tze, L.M. Quality attributes of starfruit (Averrhoa carambola L.) juice treated with ultraviolet radiation. Food Chem. 2011, 127, 641–644. [Google Scholar] [CrossRef]
- Vicentini, V.E.P.; Camparoto, M.L.; Teixeira, R.O.; Mantovani, M.S. Averrhoa carambola L., Syzygium cumini (L.) Skeels and Cissus sicyoides L.: Medicinal herbal tea effects on vegetal and animal test systems. Acta Sci. Biol. Sci. 2001, 23, 593–598. [Google Scholar]
- Zheng, N.; Lin, X.; Wen, Q.; Zhang, S.; Huang, J.; Xu, X.; Huang, R. Averrhoa carambola L. (Oxalidaceae) roots, on advanced glycation end-product-mediated renal injury in type 2 diabetic KKAy mice. Toxicol. Lett. 2013, 219, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Saghir, S.A.M.; Sadikun, A.; Khaw, K.Y.; Murugaiyah, V. Star fruit (Averrhoa carambola L.): From traditional uses to pharmacological activities Fruta de la estrella (Averrhoa carambola L.): Desde los usos tradicionales a las actividades farmacologicas]. Bol. Latinoam. Y del Caribe Plantas Med. Y Aromat. 2013, 12, 209–219. [Google Scholar]
- Yang, D.; Jia, X.; Xie, H.; Wei, X. LWT—Food science and technology further dihydrochalcone C -glycosides from the fruit of Averrhoa carambola. LWT Food Sci. Technol. 2016, 65, 604–609. [Google Scholar] [CrossRef]
- Jia, X.; Xie, H.; Jiang, Y.; Wei, X. Flavonoids isolated from the fresh sweet fruit of Averrhoa carambola, commonly known as star fruit. Phytochemistry 2018, 153, 156–162. [Google Scholar] [CrossRef]
- Chau, C.F.; Chen, C.H.; Lin, C.Y. Insoluble fiber-rich fractions derived from Averrhoa carambola: Hypoglycemic effects determined by In Vitro methods. LWT Food Sci. Technol. 2004, 37, 331–335. [Google Scholar] [CrossRef]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol. 2012, 49, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Shui, G.; Leong, L.P. Analysis of polyphenolic antioxidants in star fruit using liquid chromatography and mass spectrometry. J. Chromatogr. A 2004, 1022, 67–75. [Google Scholar] [CrossRef]
- Yang, D.; Xie, H.; Yang, B.; Wei, X. Two tetrahydroisoquinoline alkaloids from the fruit of Averrhoa carambola. Phytochem. Lett. 2014, 7, 217–220. [Google Scholar] [CrossRef]
- Pino, J.A.; Marbot, R.; Aguero, J. Volatile components of starfruit (Averrhoa carambola L.). J. Essent. Oil Res. 2000, 12, 429–430. [Google Scholar] [CrossRef]
- Forrey, R.R.; Regional, W.; Service, R. Volatile components of starfruit. Food Chem. 1974, 22, 496–498. [Google Scholar] [CrossRef]
- Frohlich, O.; Schreier, P. Additional volatile constituents of carambola (Averrhoa carambola L.) fruit. Flavour Fragr. J. 1989, 4, 177–184. [Google Scholar] [CrossRef]
- Salamon, A.; Baca, E.; Baranowski, K.; Michałowska, D. The evaluation of anti-nutritive components in beer on the example of oxalic acid. Rocz. Państwowego Zakładu Hig. 2012, 63, 37–42. [Google Scholar]
- Farag, M.A.; Fahmy, S.; Choucry, M.A.; Wahdan, M.O.; Elsebai, M.F. Metabolites profiling reveals for antimicrobial compositional differences and action mechanism in the toothbrushing stick “miswak” Salvadora persica. J. Pharm. Biomed. Anal. 2017, 133, 32–40. [Google Scholar] [CrossRef]
- Farag, M. Comparative mass spectrometry & nuclear magnetic resonance metabolomic approaches for nutraceuticals quality control analysis: A brief review. Recent Pat. Biotechnol. 2013, 8, 17–24. [Google Scholar] [CrossRef]
- Farag, M.A.; El-Kersh, D.M.; Rasheed, D.M.; Heiss, A.G. Volatiles distribution in Nigella species (black cumin seeds) and in response to roasting as analyzed via solid-phase microextraction (SPME) coupled to chemometrics. Ind. Crops Prod. 2017, 108, 564–571. [Google Scholar] [CrossRef]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef]
- Barrière, D.A.; Noll, C.; Roussy, G.; Lizotte, F.; Kessai, A.; Kirby, K.; Belleville, K.; Beaudet, N.; Longpré, J.M.; Carpentier, A.C.; et al. Combination of high-fat/high-fructose diet and low-dose streptozotocin to model long-term type-2 diabetes complications. Sci. Rep. 2018, 8, 1–17. [Google Scholar] [CrossRef]
- Mohamed Rashid, A.; Lu, K.; Yip, Y.M.; Zhang, D. Averrhoa carambola L. peel extract suppresses adipocyte differentiation in 3T3-L1 cells. Food Funct. 2016, 7, 881–892. [Google Scholar] [CrossRef]
- Ferrara, L. Averrhoa carambola Linn: Is it really a toxic fruit? Int. J. Med. Rev. 2018, 5, 2–5. [Google Scholar] [CrossRef]
- Featherstone, S. Ingredients Used in the Preparation of Canned Foods; Woodhead: Springfield, IL, USA, 2015; ISBN 9780857096784. [Google Scholar]
- Kijsamanmith, K.; Surarit, R.; Vongsavan, N. Effect of tropical fruit juices on dentine permeability and erosive ability in removing the smear layer: An In Vitro study. J. Dent. Sci. 2016, 11, 130–135. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pintaudi, B.; Di Vieste, G.; Bonomo, M. The effectiveness of Myo-Inositol and D-Chiro inositol treatment in type 2 diabetes. Int. J. Endocrinol. 2016, 2016, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.T.T.; Huang, W.; Han, C.; Li, J.; Xie, Q.; Wei, J.; Xu, X.; Lai, Z.; Huang, X.; Huang, R.B.; et al. Effects of Averrhoa carambola L. (Oxalidaceae) juice mediated on hyperglycemia, hyperlipidemia, and its influence on regulatory protein expression in the injured kidneys of streptozotocin-induced diabetic mice. Am. J. Transl. Res. 2017, 9, 36. [Google Scholar] [PubMed]
- Dotaniya, M.L.; Datta, S.C.; Biswas, D.R.; Meena, H.M.; Kumar, K. Production of oxalic acid as influenced by the application of organic residue and its effect on phosphorus uptake by wheat (Triticum aestivum L.) in an inceptisol of North India. Natl. Acad. Sci. Lett. 2014, 37, 401–405. [Google Scholar] [CrossRef][Green Version]
- Chakraborty, N.; Ghosh, R.; Ghosh, S.; Narula, K.; Tayal, R.; Datta, A.; Chakraborty, S. Reduction of oxalate levels in tomato fruit and consequent metabolic remodeling following overexpression of a fungal oxalate decarboxylase. Plant Physiol. 2013, 162, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Nguefack, N.; Borel, M.; Foba-tendo, J.; Yufanyi, D.M.; Etape, E.P.; Eko, J.N.; Ngolui, L.J. Averrhoa carambola: A renewable source of oxalic acid for the facile and green synthesis of divalent metal (Fe, Co, Ni, Zn, and Cu) oxalates and oxide nanoparticles. J. Appl. Chem. 2014, 2014, 767695. [Google Scholar] [CrossRef]
- Lombardo, V.A.; Osorio, S.; Borsani, J.; Lauxmann, M.A.; Bustamante, C.A.; Budde, C.O.; Andreo, C.S.; Lara, M.V.; Fernie, A.R.; Drincovich, M.F. Metabolic profiling during peach fruit development and ripening reveals the metabolic networks that underpin each developmental stage. Plant Physiol. 2011, 157, 1696–1710. [Google Scholar] [CrossRef]
- Kumar, A.; Bachhawat, A.K. Pyroglutamic acid: Throwing light on a lightly studied metabolite. Curr. Sci. 2012, 102, 288–297. [Google Scholar]
- Bere, E.; WiBere, E. Wild berries: A good source of omega-3. Eur. J. Clin. Nutr. 2007, 61, 431–433. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-6/Omega-3 essential fatty acid ratio and chronic diseases. Food Rev. Int. 2004, 20, 77–90. [Google Scholar] [CrossRef]
- Oliveira, I.; Baptista, P.; Malheiro, R.; Casal, S.; Bento, A.; Pereira, J.A. Influence of strawberry tree (Arbutus unedo L.) fruit ripening stage on chemical composition and antioxidant activity. Food Res. Int. 2011, 44, 1401–1407. [Google Scholar] [CrossRef]
- Soncini, R.; Santiago, M.B.; Orlandi, L.; Moraes, G.O.I.; Peloso, A.L.M.; Dos Santos, M.H.; Alves-Da-Silva, G.; Paffaro, V.A.; Bento, A.C.; Giusti-Paiva, A. Hypotensive effect of aqueous extract of Averrhoa carambola L. (Oxalidaceae) in rats: An In Vivo and In Vitro approach. J. Ethnopharmacol. 2011, 133, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.K.; Shen, C.R.; Liu, C.L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef] [PubMed]
- Määttä-Riihinen, K.R.; Kähkönen, M.P.; Törrönen, A.R.; Heinonen, I.M. Catechins and procyanidins in berries of Vaccinium species and their antioxidant activity. J. Agric. Food Chem. 2005, 53, 8485–8491. [Google Scholar] [CrossRef] [PubMed]
- Nikolaos, G.; Alexandros, P.; Evangelia, L.; Vassilik, T.; Maria-Nektaria, N. Effect of ripening stage on the total phenolics content, lycopene and antioxidant activity of tomato fruits grown to a geothermal greenhouse. Ann. Univ. Craiova 2018, 23, 115–120. [Google Scholar]
- Del Olmo, A.; Calzada, J.; Nuñez, M. Benzoic acid and its derivatives as naturally occurring compounds in foods and as additives: Uses, exposure, and controversy. Crit. Rev. Food Sci. Nutr. 2017, 57, 3084–3103. [Google Scholar] [CrossRef]
- Zainudin, M.A.M.; Hamid, A.A.; Anwar, F.; Osman, A.; Saari, N. Variation of bioactive compounds and antioxidant activity of carambola (Averrhoa carambola L.) fruit at different ripening stages. Sci. Hortic. 2014, 172, 325–331. [Google Scholar] [CrossRef]
- Khalil, M.N.A.; Fekry, M.I.; Farag, M.A. Metabolome based volatiles profiling in 13 date palm fruit varieties from Egypt via SPME GC—MS and chemometrics. Food Chem. 2017, 217, 171–181. [Google Scholar] [CrossRef]
- Beekwilder, J. Functional characterization of enzymes forming volatile esters from strawberry and banana. Plant Physiol. 2004, 135, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Tsuzuki, S. Higher straight-chain aliphatic aldehydes: Importance as odor-active volatiles in human foods and issues for future research. J. Agric. Food Chem. 2019, 67, 4720–4725. [Google Scholar] [CrossRef] [PubMed]
- Ozkara, K.T.; Amanpour, A.; Guclu, G.; Kelebek, H.; Selli, S. GC-MS-Olfactometric differentiation of aroma-active compounds in turkish heat-treated sausages by application of aroma extract dilution analysis. Food Anal. Methods 2019, 12, 729–741. [Google Scholar] [CrossRef]
- McCrummen, S.T.; Wang, Y.; Hanson, T.R.; Bott, L.; Liu, S. Culture environment and the odorous volatile compounds present in pond-raised channel catfish (Ictalurus punctatus). Aquac. Int. 2018, 26, 685–694. [Google Scholar] [CrossRef]
- Winterhalter, P.; Rouseff, R. Carotenoid-derived aroma compounds: An introduction. ACS Symp. Ser. 2001, 802, 1–17. [Google Scholar] [CrossRef]
- Lopez, P.; Van Sisseren, M.; De Marco, S.; Jekel, A.; De Nijs, M.; Mol, H.G.J. A straightforward method to determine flavouring substances in food by GC-MS. Food Chem. 2015, 174, 407–416. [Google Scholar] [CrossRef]
- Hausner, E.A.; Robert, H. Poppenga Hazards Associated with the Use of Herbal and Other Natural Products. In Small Animal Toxicology, 3rd ed.; Peterson, M.E., Talcott, P.A., Eds.; Saunders: St. Louis, MO, USA, 2013; pp. 335–356. ISBN 9781455707171. [Google Scholar]
- Kongor, J.E.; Hinneh, M.; de Walle, D.V.; Afoakwa, E.O.; Boeckx, P.; Dewettinck, K. Factors influencing quality variation in cocoa (Theobroma cacao) bean flavour profile—A review. Food Res. Int. 2016, 82, 44–52. [Google Scholar] [CrossRef]
- Kashima, Y.; Nakaya, S.; Miyazawa, M. Volatile composition and sensory properties of indian herbal medicine-pavonia odorata-used in ayurveda. J. Oleo Sci. 2014, 63, 149–158. [Google Scholar] [CrossRef]
- Les, F.; Arbonés-mainar, J.M.; Sofía, M.; López, V. Pomegranate polyphenols and urolithin A inhibit α -glucosidase, dipeptidyl peptidase-4, lipase, triglyceride accumulation and adipogenesis related genes in 3T3-L1 adipocyte-like cells. J. Ethnopharmacol. 2018, 220, 67–74. [Google Scholar] [CrossRef]
- Wan-Nadilah, W.A.; Akhtar, M.T.; Shaari, K.; Khatib, A.; Hamid, A.A.; Hamid, M. Variation in the metabolites and α-glucosidase inhibitory activity of Cosmos caudatus at different growth stages. BMC Complement. Altern. Med. 2019, 19, 1–15. [Google Scholar] [CrossRef]
- Yang, C.; Yen, Y.; Hung, K.; Hsu, S.; Lan, S.; Lin, H. Inhibitory effects of pu-erh tea on alpha glucosidase and alpha amylase: A systemic review. Nutr. Diabetes 2019, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kang, M.-J.; Kim, M.-J.; Kim, M.-E.; Song, J.-H.; Lee, Y.-M.; Kim, J.-I. Pancreatic lipase inhibitory activity of taraxacum officinale In Vitro and In Vivo. Nutr. Res. Pract. 2008, 2, 200. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Rasheed, D.M.; Kamal, I.M. Volatiles and primary metabolites profiling in two Hibiscus sabdariffa (roselle) cultivars via headspace SPME-GC-MS and chemometrics. Food Res. Int. 2015, 78, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Porzel, A.; Wessjohann, L.A. Comparative metabolite profiling and fingerprinting of medicinal licorice roots using a multiplex approach of GC–MS, LC–MS and 1D NMR techniques. Phytochemistry 2012, 76, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Gad, H.A.; Heiss, A.G.; Wessjohann, L.A. Metabolomics driven analysis of six Nigella species seeds via UPLC-qTOF-MS and GC–MS coupled to chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef]
- Elmassry, M.M.; Kormod, L.; Labib, R.M.; Farag, M.A. Metabolome based volatiles mapping of roasted umbelliferous fruits aroma via HS-SPME GC/MS and peroxide levels analyses. J. Chromatogr. B 2018, 1099, 117–126. [Google Scholar] [CrossRef]
- Farag, M.A.; Wessjohann, L.A. Volatiles profiling in medicinal licorice roots using steam distillation and solid-phase microextraction (SPME) coupled to chemometrics. J. Food Sci. 2012, 77, C1179–C1184. [Google Scholar] [CrossRef]
- Farag, M.A.; Khattab, A.R.; Ehrlich, A.; Kropf, M.; Heiss, A.G.; Wessjohann, L.A. Gas chromatography/mass spectrometry-based metabolite profiling of nutrients and antinutrients in eight lens and lupinus seeds (Fabaceae). J. Agric. Food Chem. 2018, 66, 4267–4280. [Google Scholar] [CrossRef]
- Farag, M.A.; Huhman, D.V.; Lei, Z.; Sumner, L.W. Metabolic profiling and systematic identification of flavonoids and isoflavonoids in roots and cell suspension cultures of Medicago truncatula using HPLC-UV-ESI-MS and GC-MS. Phytochemistry 2007, 68, 342–354. [Google Scholar] [CrossRef]
- Farag, M.A.; Mohsen, M.; Heinke, R.; Wessjohann, L.A. Metabolomic fingerprints of 21 date palm fruit varieties from Egypt using UPLC/PDA/ESI-qTOF-MS and GC-MS analyzed by chemometrics. Food Res. Int. 2014, 64, 218–226. [Google Scholar] [CrossRef]
- Fatoki, O.S. Determination of oxalic acid in vegetables. Veg. Veg. Prod. 1994, 16, 161–167. [Google Scholar] [CrossRef]
- Chemists, A.; Helrich, K. Official Methods of Analysis of the Association of Official Analytical Chemists; The Association: Arlington, VA, USA, 1990; Volume 2, ISBN 0935584420 9780935584424. [Google Scholar]
- Lateef, S.S. Determining total sugar content in maple syrup to meet FDA nutrition labeling requirements application note. Agil. Technol. 2011, 1–8, Publication Number 5990-7935EN. [Google Scholar]
- Sumner, L.W.; Broeckling, C.D.; Reddy, I.R.; Duran, A.L.; Zhao, X.C. MET-IDEA: Data extraction tool for mass spectrometry-based metabolomics. Anal. Chem. 2006, 78, 4334–4341. [Google Scholar] [CrossRef]
- Les, F.; Venditti, A.; Cásedas, G.; Frezza, C.; Guiso, M. Everlasting flower (Helichrysum stoechas Moench) as a potential source of bioactive molecules with antiproliferative, antioxidant, antidiabetic and neuroprotective properties. Ind. Crop. Prod. 2017, 108, 295–302. [Google Scholar] [CrossRef]
Sample Availability: Samples of the plants are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak # | RT (min) | KI | Name | Class | CLI | CLM | CLRE | CLURE |
|---|---|---|---|---|---|---|---|---|
| 1 | 7.34 | 1079 | Lactic acid 2TMS* | Acid | 0.39 ± 0.22 | 0.41 ± 0.05 | 0.17 ± 0.02 | 0.97 ± 0.69 |
| 2 | 7.69 | 1096 | Glycolic acid 2TMS | 0.01 ± 0.00 | - | - | 0.30 ± 0.39 | |
| 3 | 8.58 | 1133 | Oxalic acid 2TMS | 1.97 ± 2.00 | 0.07 ± 0.00 | 2.71 ± 2.24 | 1.67 ± 0.33 | |
| 4 | 10.45 | 1210 | Malonic acid 2TMS | 0.06 ± 0.00 | 0.02 ± 0.00 | 0.03 ± 0.01 | 0.82 ± 0.86 | |
| 5 | 11.29 | 1242 | Methylmalonate 2TMS | 0.02 ± 0.01 | 0.02 ± 0.00 | 0.29 ± 0.05 | 1.29 ± 0.87 | |
| 6 | 12.91 | 1305 | Maleic acid 2TMS | 0.26 ± 0.07 | 0.20 ± 0.04 | 0.07 ± 0.01 | 1.37 ± 0.81 | |
| 7 | 13.21 | 1316 | Succinic acid 2TMS | 0.01 ± 0.00 | - | - | 0.74 ± 0.14 | |
| 8 | 13.56 | 1330 | Glyceric acid3TMS | 0.02 ± 0.01 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.74 ± 0.14 | |
| 9 | 13.85 | 1342 | 2,3-Dihydroxybutanoic acid 3TMS | 0.01 ± 0.00 | 0.01 ± 0.00 | - | 0.66 ± 0.11 | |
| 10 | 14.12 | 1352 | Fumaric acid 2TMS | 0.15 ± 0.02 | 0.11 ± 0.00 | 0.04 ± 0.00 | 1.12 ± 0.64 | |
| 11 | 14.39 | 1362 | Nonanoic acid, TMS ester | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.02 ± 0.00 | 1.51 ± 0.61 | |
| 12 | 14.52 | 1367 | Citramalic acid 2TMS ester | 0.09 ± 0.02 | 0.11 ± 0.00 | 0.13 ± 0.01 | 1.07 ± 0.86 | |
| 13 | 15.19 | 1393 | Pentonic acid, 5-deoxy-3 TMS, γ-lactone | - | - | - | 0.33 ± 0.44 | |
| 14 | 15.54 | 1407 | Glutaric acid 2TMS | - | - | - | 0.68 ± 0.10 | |
| 15 | 15.78 | 1417 | 2-Methylglutaric acid, 2TMS- ester | - | - | - | 0.78 ± 0.10 | |
| 16 | 16.03 | 1427 | Erythronic acid γ-lactone, 2 TMS-ether | 0.01 ± 0.01 | - | - | 0.77 ± 0.13 | |
| 17 | 17.62 | 1490 | Malic acid 3TMS* | 1.89 ± 0.02 | −1.41 ± 0.05 | 0.56 ± 0.05 | 0.65 ± 0.09 | |
| 18 | 18.00 | 1506 | Adipic acid 2TMS | - | - | - | 1.34 ± 0.67 | |
| 19 | 20.16 | 1595 | Tartaric acid 2TMS-ether, 2TMS ester | 0.06 ± 0.03 | 0.04 ± 0.01 | - | 0.63 ± 0.11 | |
| 20 | 24.61 | 1794 | Azelaic acid 2TMS | 0.04 ± 0.00 | 0.01 ± 0.00 | 0.04 ± 0.00 | 0.92 ± 0.11 | |
| 21 | 25.03 | 1814 | Citric acid 4TMS* | 0.14 ± 0.04 | 0.11 ± 0.04 | 0.12 ± 0.03 | 0.54 ± 0.32 | |
| 22 | 27.63 | 1940 | Ascorbic acid 4TMS-ether | - | - | - | 0.65 ± 0.09 | |
| Total acids | 5.16 | 2.55 | 4.21 | 19.54 | ||||
| 23 | 11.42 | 1247 | Diethylene glycol, 2TMS | Alcohol | - | - | - | 0.57±0.44 |
| 24 | 15.03 | 1387 | Ethylene glycol 2TMS* | 0.03 ± 0.01 | 0.02 ± 0.00 | 0.03 ± 0.00 | 1.17 ± 0.11 | |
| 25 | 45.42 | 3047 | Nonaethylene glycol 2TMS | 0.05 ± 0.05 | 0.02 ± 0.02 | 0.03 ± 0.02 | 2.24 ± 0.40 | |
| 26 | 46.61 | 3136 | Octacosyl TMS | - | - | - | 0.69 ± 0.10 | |
| 27 | 49.24 | 3305 | Decaethylene glycol 2TMS | - | - | - | 0.68 ± 0.11 | |
| 28 | 49.75 | 3332 | 1-Triacontanol TMS | 0.02 ± 0.00 | - | - | 0.56 ± 0.25 | |
| Total alcohols | 0.10 | 0.06 | 0.07 | 5.91 | ||||
| 29 | 14.68 | 1373 | 3,4-[(2)Trimethylsiloxy]dihydro-2(3H)-furanone | Aldehyde/furan | - | - | 0.01 ± 0.00 | 0.64 ± 0.32 |
| Total furan | 0.00 | 0.00 | 0.01 | 0.64 | ||||
| 30 | 7.82 | 1102 | Alanine 2TMS | Amino acid | 0.06 ± 0.02 | 0.02 ± 0.00 | 0.02 ± 0.01 | 0.70 ± 0.14 |
| 31 | 9.83 | 1185 | Proline TMS* | 0.27 ± 0.05 | 0.07 ± 0.01 | 0.07 ± 0.01 | 0.16 ± 0.10 | |
| 32 | 10.64 | 1217 | Valine 2TMS* | 0.05 ± 0.00 | 0.03 ± 0.00 | 0.02 ± 0.00 | 0.69 ± 0.16 | |
| 33 | 11.75 | 1260 | Serine 2TMS- | 0.57 ± 0.00 | 0.26 ± 0.02 | 0.34 ± 0.05 | 0.59 ± 0.38 | |
| 34 | 12.65 | 1295 | Threonine, O-TMS, TMS ester | 0.10 ± 0.03 | 0.04 ± 0.01 | 0.06 ± 0.01 | 1.19 ± 0.96 | |
| 35 | 14.30 | 1359 | Serine 3TMS | 0.08 ± 0.07 | - | - | 0.67 ± 0.11 | |
| 36 | 14.94 | 1384 | Threonine 3TMS | 0.01 ± 0.01 | - | 0.01 ± 0.00 | 0.66 ± 0.10 | |
| 37 | 15.94 | 1423 | Aspartic acid 2 TMS ester | 0.35 ± 0.05 | 0.10 ± 0.03 | 0.04 ± 0.01 | 0.95 ± 0.18 | |
| 38 | 17.16 | 1472 | Proline TMS* | 0.12 ± 0.00 | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.78 ± 0.11 | |
| 39 | 18.29 | 1518 | Pyroglutamic acid 2TMS | 1.51 ± 0.18 | 0.35 ± 0.02 | 0.57 ± 0.20 | 1.01 ± 0.41 | |
| Total amino acids | 3.12 | 0.89 | 1.15 | 7.40 | ||||
| 40 | 11.60 | 1254 | Benzoic acid TMS | Aromatic | 0.04 ± 0.03 | 0.01 ± 0.00 | 0.01 ± 0.00 | 1.03 ± 0.51 |
| 41 | 42.74 | 2849 | Catechin, 5TMS | 0.04 ± 0.04 | 0.74 ± 0.20 | 1.42 ± 0.02 | 0.69 ± 0.11 | |
| 42 | 43.34 | 2892 | Thymol-β-glucopyranoside-O-TMS | 0.08 ± 0.02 | 0.02 ± 0.01 | 0.05 ± 0.01 | 0.79 ± 0.14 | |
| 43 | 46.49 | 3127 | α-Tocopherol, O-TMS | - | 0.01 ± 0.01 | - | 0.77 ± 0.12 | |
| Total aromatics | 0.17 | 0.79 | 1.47 | 3.28 | ||||
| 44 | 25.72 | 1847 | Myristic acid TMS | Free fatty acid | 0.11 ± 0.00 | 0.11 ± 0.01 | 0.14 ± 0.01 | 0.41 ± 0.35 |
| 45 | 29.70 | 2047 | Palmitic acid TMS* | 0.02 ± 0.01 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.81 ± 0.09 | |
| 46 | 30.74 | 2102 | Methyl 8-octadecenoate | - | - | - | 0.90 ± 0.76 | |
| 47 | 31.23 | 2129 | Methyl stearate | - | - | - | 0.51 ± 0.53 | |
| 48 | 32.64 | 2206 | Linoleic acid TMS* | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.66 ± 0.11 | |
| 49 | 32.75 | 2213 | α-Linolenic acid TMS | 0.03 ± 0.02 | 0.03 ± 0.00 | 0.03 ± 0.00 | 0.87 ± 0.12 | |
| 50 | 32.76 | 2214 | Octadecenoic acid TMS | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.01 ± 0.00 | 0.73 ± 0.10 | |
| 51 | 32.88 | 2221 | Oleic acid TMS | 0.01 ± 0.01 | 0.01 ± 0.00 | 0.01 ± 0.00 | 1.17 ± 0.28 | |
| 52 | 33.23 | 2241 | Stearic acid TMS* | 0.06 ± 0.02 | 0.05 ± 0.00 | 0.06 ± 0.00 | 0.88 ± 0.51 | |
| 53 | 38.84 | 2583 | 1-Monopalmitin TMS | 0.09 ± 0.05 | 0.08 ± 0.03 | 0.06 ± 0.00 | 0.47 ± 0.41 | |
| 54 | 41.70 | 2775 | Stearic acid, 2,3-[2trimethylsiloxy]propyl ester | 0.06 ± 0.03 | 0.06 ± 0.01 | 0.05 ± 0.01 | 0.65 ± 0.82 | |
| Total free fatty acids | 0.42 | 0.40 | 0.39 | 8.06 | ||||
| 55 | 34.29 | 2301 | Tricosane | Hydrocarbon | 0.01 ± 0.01 | - | - | 0.82 ± 0.02 |
| 56 | 37.56 | 2500 | Pentacosane | - | - | - | 0.33 ± 0.45 | |
| 57 | 39.09 | 2599 | Hexacosane | 0.04 ± 0.01 | 0.02±0.00 | 0.02 ± 0.01 | 0.70 ± 0.10 | |
| 58 | 40.60 | 2699 | Heptacosane | 0.02 ± 0.00 | - | 0.04 ± 0.00 | 0.68 ± 0.09 | |
| 59 | 43.43 | 2898 | Nonacosane | 0.01 ± 0.00 | - | 0.01 ± 0.00 | 0.66 ± 0.10 | |
| Total hydrocarbons | 0.08 | 0.03 | 0.06 | 3.20 | ||||
| 60 | 12.11 | 1274 | Phosphoric acid 3TMS | Inorganic | 2.31 ± 0.44 | 1.85 ± 0.46 | 2.05 ± 0.20 | 0.64 ± 0.10 |
| Total inorganic | 2.31 | 1.85 | 2.05 | 0.64 | ||||
| 61 | 9.14 | 1156 | 2-Pyrrolidinone TMS | Nitrogenous | 0.02 ± 0.01 | - | 0.01 ± 0.00 | 0.64 ± 0.44 |
| 62 | 11.57 | 1253 | Urea 2TMS | 0.01 ± 0.01 | - | 0.02 ± 0.00 | 0.66 ± 0.11 | |
| 63 | 11.85 | 1264 | Aminoethanol, O,N,N-3 TMS | 0.13 ± 0.00 | 0.04 ± 0.00 | 0.10 ± 0.02 | 0.58 ± 0.39 | |
| 64 | 18.50 | 1526 | GABA 3TMS | 1.58 ± 0.51 | 0.36 ± 0.05 | 0.34 ± 0.07 | 0.80 ± 0.08 | |
| 65 | 21.85 | 1669 | unknown nitrogenous | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.68 ± 0.09 | |
| 66 | 30.18 | 2072 | N-Acetyl-d-glucosamine 4 TMS | 0.35 ± 0.44 | 0.65 ± 0.01 | 0.08 ± 0.03 | 2.09 ± 3.21 | |
| 67 | 35.28 | 2360 | Oleic acid amide | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 1.02 ± 0.29 | |
| 68 | 36.10 | 2409 | Oleamide, N-TMS | 0.33 ± 0.09 | 0.23 ± 0.02 | 0.25 ± 0.07 | 1.47 ± 0.23 | |
| 69 | 37.41 | 2490 | (E)-13-Docosenamide | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.27 ± 0.42 | |
| Total nitrogenous | 2.45 | 1.32 | 0.84 | 8.22 | ||||
| 70 | 48.12 | 3237 | Campesterol TMS | Sterol | 0.07 ± 0.00 | 0.06 ± 0.03 | 0.05 ± 0.01 | 1.18 ± 0.52 |
| 71 | 49.55 | 3322 | β-Sitosterol TMS* | 0.18 ± 0.02 | 0.14 ± 0.02 | 0.16 ± 0.01 | 1.13 ± 0.84 | |
| Total sterols | 0.25 | 0.19 | 0.21 | 2.31 | ||||
| 72 | 16.67 | 1452 | Threose 3TMS | Sugar | - | - | - | 0.80 ± 0.13 |
| 73 | 18.83 | 1540 | Threonic acid 3TMS | - | - | - | 0.63 ± 0.11 | |
| 74 | 19.25 | 1558 | Erythronic acid 4TMS | 0.06 ± 0.04 | 0.04 ± 0.01 | 0.11 ± 0.00 | 0.63 ± 0.10 | |
| 75 | 20.54 | 1611 | Ribofuranose 4TMS (isomer 1) | 0.03 ± 0.00 | 0.03 ± 0.00 | 0.09 ± 0.01 | 0.76 ± 0.13 | |
| 76 | 20.79 | 1622 | Arabinonic acid, 3TMS γ-lactone | - | - | 0.01 ± 0.00 | 0.63 ± 0.11 | |
| 77 | 21.47 | 1652 | Lyxose 4TMS | 0.12 ± 0.00 | 0.14 ± 0.01 | 0.08 ± 0.01 | 0.76 ± 0.13 | |
| 78 | 21.63 | 1659 | Arabinose 4TMS | 0.06 ± 0.01 | 0.03 ± 0.01 | 0.06 ± 0.00 | 0.63 ± 0.10 | |
| 79 | 22.07 | 1679 | Xylose 4TMS | 0.03 ± 0.00 | 0.02 ± 0.00 | 0.06 ± 0.01 | 0.77 ± 0.12 | |
| 80 | 22.25 | 1686 | Mannose, 6-deoxy-2,3,4,5-4-O-TMS | 0.02 ± 0.02 | - | 0.01 ± 0.00 | 0.64 ± 0.10 | |
| 81 | 22.49 | 1697 | Levoglucosan 3TMS | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.02 ± 0.00 | 0.79 ± 0.12 | |
| 82 | 22.81 | 1711 | Rhamnose 4TMS | 0.11 ± 0.00 | 0.06 ± 0.01 | 0.07 ± 0.01 | 2.84 ± 0.93 | |
| 83 | 22.97 | 1718 | Fucose 4TMS | 0.03 ± 0.00 | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.64 ± 0.10 | |
| 84 | 23.94 | 1763 | Sorbofuranose 5TMS | - | - | 0.10 ± 0.02 | 0.65 ± 0.11 | |
| 85 | 24.22 | 1776 | Tagatofuranose 5TMS | 1.21 ± 0.13 | 1.18 ± 0.01 | 3.69 ± 0.40 | 0.59 ± 0.47 | |
| 86 | 24.69 | 1797 | Fructofuranose 5TMS | 0.17 ± 0.15 | 0.38 ± 0.34 | 2.16 ± 1.98 | 0.86 ± 0.05 | |
| 87 | 26.13 | 1866 | Fructose 5TMS* | 31.45 ± 1.01 | 28.68 ± 3.44 | 29.96 ± 1.35 | 0.65 ± 0.09 | |
| 88 | 26.64 | 1891 | Glucose 5 TMS* | 23.08 ± 1.02 | 20.73 ± 4.26 | 24.93 ± 0.29 | 0.43 ± 0.36 | |
| 89 | 26.77 | 1897 | Fructose 5TMS* | 16.47 ± 0.05 | 15.95 ± 0.00 | 8.48 ± 0.54 | 0.32 ± 0.35 | |
| 90 | 26.98 | 1908 | Sorbose 5TMS | 1.14 ± 0.01 | 1.08 ± 0.08 | 0.85 ± 0.26 | 0.64 ± 0.10 | |
| 91 | 26.99 | 1908 | Glucose 5 TMS* | 0.03 ± 0.01 | 0.02 ± 0.01 | 0.05 ± 0.01 | 0.67 ± 0.10 | |
| 92 | 28.35 | 1977 | Gulose, 5TMS | 0.63 ± 0.12 | 0.61 ± 0.10 | 0.62 ± 0.04 | 0.87 ± 0.76 | |
| 93 | 28.41 | 1980 | Mannose 5TMS | 0.96 ± 0.65 | 1.46 ± 0.83 | 2.05 ± 0.08 | 0.27 ± 0.24 | |
| 94 | 28.57 | 1988 | Galactose 5TMS | 0.52 ± 0.14 | 0.24 ± 0.04 | 0.02 ± 0.00 | 0.66 ± 0.08 | |
| 95 | 28.62 | 1990 | Gluconic acid 6TMS | 0.59 ± 0.08 | 0.31 ± 0.06 | 0.25 ± 0.04 | 0.54 ± 0.30 | |
| 96 | 29.25 | 2023 | Allose 5TMS-ether TMS | 2.44 ± 0.11 | 2.36 ± 0.06 | 2.12 ± 0.30 | 0.87 ± 0.11 | |
| 97 | 30.71 | 2100 | Sedoheptulose 6TMS | 0.04 ± 0.00 | 0.03 ± 0.00 | 0.13 ± 0.01 | 1.01 ± 0.50 | |
| 98 | 31.13 | 2123 | Sedoheptulose 6TMS | 0.10 ± 0.01 | 0.03 ± 0.01 | 0.04 ± 0.00 | 1.58 ± 1.34 | |
| 99 | 31.79 | 2159 | Fructopyranose, 4TMS | 0.05 ± 0.01 | 0.01 ± 0.00 | - | 0.73 ± 0.10 | |
| 100 | 32.20 | 2182 | Glucopyranose, 5TMS | 0.11 ± 0.03 | 0.03 ± 0.01 | 0.05 ± 0.01 | 0.86 ± 0.12 | |
| 101 | 37.50 | 2496 | O-β-Galactopyranosyl-d-mannopyranose 8TMS | 0.31 ± 0.04 | 0.21 ± 0.04 | 0.13 ± 0.02 | 0.62 ± 0.31 | |
| 102 | 38.98 | 2591 | Lactose 8TMS | 0.01 ± 0.00 | 0.02 ± 0.01 | 0.01 ± 0.00 | 0.66 ± 0.10 | |
| 103 | 39.50 | 2626 | Sucrose 8TMS* | 0.64 ± 0.18 | 5.79 ± 0.35 | 7.28 ± 0.39 | 0.77 ± 0.13 | |
| 104 | 40.24 | 2675 | Mannobiose 8TMS | 0.30 ± 0.01 | 0.22 ± 0.01 | 0.76 ± 0.07 | 0.87 ± 0.24 | |
| 105 | 40.59 | 2698 | Cellobiose 8TMS | 0.09 ± 0.05 | 0.05 ± 0.00 | 0.06 ± 0.00 | 0.86 ± 0.13 | |
| 106 | 40.95 | 2723 | Sucrose 8TMS* | 0.35 ± 0.06 | 0.24 ± 0.02 | 0.65 ± 0.09 | 0.79 ± 0.14 | |
| 107 | 41.05 | 2731 | Trehalose 8TMS | 0.15 ± 0.04 | 0.07 ± 0.01 | 0.05 ± 0.00 | 0.97 ± 0.22 | |
| 108 | 41.25 | 2744 | Maltose 8TMS | 0.31 ± 0.06 | 0.38 ± 0.01 | 0.69 ± 0.07 | 0.56 ± 0.29 | |
| 109 | 42.25 | 2814 | Gentiobiose 8TMS | 0.12 ± 0.01 | 0.08 ± 0.00 | 0.10 ± 0.01 | 0.86 ± 0.15 | |
| 110 | 42.27 | 2816 | Isomaltulose 7TMS | 0.14 ± 0.01 | 0.06 ± 0.01 | 0.07 ± 0.00 | 0.80 ± 0.15 | |
| 111 | 42.91 | 2861 | Gentiobiose 8TMS | 0.19 ± 0.01 | 0.10 ± 0.01 | 0.16 ± 0.01 | 0.82 ± 0.13 | |
| 112 | 45.03 | 3018 | unidentified disaccharide | 0.01 ± 0.00 | - | - | 1.32 ± 0.20 | |
| Total sugars | 82.07 | 80.68 | 85.96 | 32.68 | ||||
| 113 | 17.81 | 1498 | Threitol 4TMS | Polyol | 0.01 ± 0.00 | - | - | 0.47 ± 0.39 |
| 114 | 23.32 | 1735 | Fucitol 5TMS | 0.03 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.54 ± 0.50 | |
| 115 | 27.40 | 1929 | Sorbitol 6TMS | 2.88 ± 0.20 | 10.35 ± 7.10 | 2.43 ± 0.07 | 0.66 ± 0.08 | |
| 116 | 30.39 | 2083 | Myo-Inositol 6TMS | 0.80 ± 0.21 | 0.82± 0.07 | 1.02 ± 0.08 | 5.57 ± 7.45 | |
| 117 | 43.89 | 2932 | Galactinol 9TMS | 0.14 ± 0.00 | 0.03 ± 0.00 | 0.11 ± 0.01 | 0.90 ± 0.15 | |
| Total polyols | 3.87 | 11.23 | 3.58 | 8.13 | ||||
| Total 100% | 100.00 | 100.00 | 100.00 | 100.00 |
| Peak # | RT (min) | RI | Name | Class | CLRE | CLI | CLM |
|---|---|---|---|---|---|---|---|
| 1 | 6.41 | 873 | 2-Heptanone | Ketone | 5.69 | - | - |
| 2 | 9.52 | 1051 | 2-Hydroxy-2-methylhept-6-en-3-one | 1.61 | 6.17 | 3.69 | |
| 3 | 9.78 | 1068 | 2-Nonanone | 2.00 | - | - | |
| 4 | 10.70 | 1129 | 4-Ketoisophorone | - | 2.55 | - | |
| 5 | 11.24 | 1167 | 3-Hexanone, 2,4-dimethyl- | 1.25 | 2.73 | 9.02 | |
| 6 | 14.43 | 1415 | Nerylacetone | - | 12.13 | 12.72 | |
| Total ketones | 10.55 | 23.58 | 25.44 | ||||
| 7 | 7.00 | 902 | Methyl caproate | Ester | 49.82 | - | - |
| 8 | 8.29 | 976 | Ethyl caproate | 10.32 | - | - | |
| 9 | 10.11 | 1089 | 3-Hexen-1-ol, propanoate, (Z)- | 0.22 | - | - | |
| 10 | 10.23 | 1097 | Methyl caprylate | 4.23 | - | - | |
| 11 | 11.96 | 1219 | Linalyl acetate | - | 0.69 | - | |
| 12 | 12.54 | 1263 | Oxalic acid, butyl propyl ester | 1.20 | 1.30 | 2.91 | |
| 13 | 16.17 | 1548 | Oxalic acid, heptyl propyl ester | - | 3.84 | 6.49 | |
| Total esters | 65.79 | 5.83 | 9.40 | ||||
| 14 | 9.99 | 1081 | Nonanal | Aldehyde | 16.49 | 13.82 | 33.46 |
| 15 | 11.43 | 1181 | Decanal | 0.10 | 6.16 | - | |
| 16 | 11.73 | 1202 | β-Cyclocitral | - | 4.89 | - | |
| 17 | 15.45 | 1499 | Myristicin | - | 16.82 | 4.61 | |
| Total aldehydes | 16.59 | 41.69 | 38.07 | ||||
| 18 | 8.11 | 965 | Myrcene | Monoterpene hydrocarbon | 0.46 | - | - |
| 19 | 8.82 | 1006 | Limonene | 1.40 | - | - | |
| Total monoterpene hydrocarbons | 1.86 | - | - | ||||
| 20 | 8.62 | 994 | Hexanoic acid | Acid | 3.85 | - | - |
| Total acids | 3.85 | - | - | ||||
| 21 | 10.18 | 1094 | 2,5-dimethyl cyclohexanol | Alcohol | - | 9.41 | - |
| Total alcohols | - | 9.41 | - | ||||
| 22 | 12.63 | 1270 | (E)-Anethole | Oxide | 0.45 | - | - |
| 23 | 12.68 | 1274 | Safrole | - | 8.53 | - | |
| Total oxides | 0.45 | 8.53 | - | ||||
| 24 | 13.75 | 1360 | Tridecane, 4-methyl- | Aliphatic hydrocarbon | 0.92 | 10.97 | 18.07 |
| Total aliphatic hydrocarbons | 0.92 | 10.97 | 18.07 | ||||
| Total | 99.56 | 100.00 | 90.98 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramadan, N.S.; Wessjohann, L.A.; Mocan, A.; C Vodnar, D.; H. El-Sayed, N.; A. El-Toumy, S.; Abdou Mohamed, D.; Abdel Aziz, Z.; Ehrlich, A.; A. Farag, M. Nutrient and Sensory Metabolites Profiling of Averrhoa Carambola L. (Starfruit) in the Context of Its Origin and Ripening Stage by GC/MS and Chemometric Analysis. Molecules 2020, 25, 2423. https://doi.org/10.3390/molecules25102423
Ramadan NS, Wessjohann LA, Mocan A, C Vodnar D, H. El-Sayed N, A. El-Toumy S, Abdou Mohamed D, Abdel Aziz Z, Ehrlich A, A. Farag M. Nutrient and Sensory Metabolites Profiling of Averrhoa Carambola L. (Starfruit) in the Context of Its Origin and Ripening Stage by GC/MS and Chemometric Analysis. Molecules. 2020; 25(10):2423. https://doi.org/10.3390/molecules25102423
Chicago/Turabian StyleRamadan, Nehal S., Ludger A. Wessjohann, Andrei Mocan, Dan C Vodnar, Nabil H. El-Sayed, Sayed A. El-Toumy, Doha Abdou Mohamed, Zeinab Abdel Aziz, Anja Ehrlich, and Mohamed A. Farag. 2020. "Nutrient and Sensory Metabolites Profiling of Averrhoa Carambola L. (Starfruit) in the Context of Its Origin and Ripening Stage by GC/MS and Chemometric Analysis" Molecules 25, no. 10: 2423. https://doi.org/10.3390/molecules25102423
APA StyleRamadan, N. S., Wessjohann, L. A., Mocan, A., C Vodnar, D., H. El-Sayed, N., A. El-Toumy, S., Abdou Mohamed, D., Abdel Aziz, Z., Ehrlich, A., & A. Farag, M. (2020). Nutrient and Sensory Metabolites Profiling of Averrhoa Carambola L. (Starfruit) in the Context of Its Origin and Ripening Stage by GC/MS and Chemometric Analysis. Molecules, 25(10), 2423. https://doi.org/10.3390/molecules25102423