
DNA Origami “Quick” Refolding inside of a Micron-Sized Compartment

 , ,
, ,
Abstract
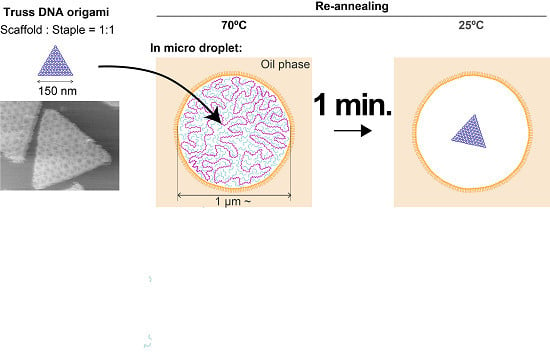
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. DNA Origami Design and Preparation
4.3. Preparation of Droplets Containing DNA Origami
4.4. DNA Sample Extraction from Droplets
4.5. Agarose Gel Electrophoresis
4.6. AFM Observation
4.7. DLS Measurement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anfinsen, C.B. Principles that govern the folding of protein chains. Science 1973, 181, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Hendrick, J.P.; Hartl, F.U. Molecular chaperone functions of heat-shock proteins. Ann. Rev. Biochem. 1993, 62, 349–384. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R.D. Structure of chromatin. Ann. Rev. Biochem. 1977, 46, 931–954. [Google Scholar] [CrossRef] [PubMed]
- Noll, M. DNA folding in the nucleosome. J. Mol. Biol. 1977, 116, 49–71. [Google Scholar] [CrossRef]
- Seeman, N.C. DNA in a material world. Nature 2003, 421, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Nangreave, J.; Liu, Y.; Yan, H. Structural DNA nanotechnology: State of the art and future perspective. J. Am. Chem. Soc. 2014, 136, 11198–11211. [Google Scholar] [CrossRef]
- Endo, M.; Yang, Y.; Sugiyama, H. DNA origami technology for biomaterials applications. Biomat. Sci. 2013, 1, 347–360. [Google Scholar] [CrossRef]
- Linko, V.; Dietz, H. The enabled state of DNA nanotechnology. Curr. Opin. Biotech. 2013, 24, 555–561. [Google Scholar] [CrossRef]
- Rothemund, P.W. Folding DNA to create nanoscale shapes and patterns. Nature 2006, 440, 297–302. [Google Scholar] [CrossRef]
- Saccà, B.; Niemeyer, C.M. DNA origami: The art of folding DNA. Angew. Chem. Int. Ed. 2012, 51, 58–66. [Google Scholar] [CrossRef]
- Gerling, T.; Kube, M.; Kick, B.; Dietz, H. Sequence-Programmable covalent bonding of designed DNA assemblies. Sci. Adv. 2018, 4, eaau1157. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, M.; Nigorikawa, S.; Sakaue, T.; Fujiwara, K.; Tokita, M. Multiple patterns of polymer gels in microspheres due to the interplay among phase separation, wetting, and gelation. Proc. Nat. Acad. Sci. USA 2014, 111, 15894–15899. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Yanagisawa, M.; Sato, Y.T.; Fujiwara, K.; Yoshikawa, K. Cell-Sized confinement in microspheres accelerates the reaction of gene expression. Sci. Rep. 2012, 2, 283. [Google Scholar] [CrossRef] [PubMed]
- Fallah-Araghi, A.; Meguellati, K.; Baret, J.C.; El Harrak, A.; Mangeat, T.; Karplus, M.; Ladame, S.; Marques, C.M.; Griffiths, A.D. Enhanced chemical synthesis at soft interfaces: A universal reaction-adsorption mechanism in microcompartments. Phy. Rev. Lett. 2014, 112, 028301. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.-I.M.; Tsumoto, K.; Hamada, T.; Akiyoshi, K.; Nakatani, Y.; Yoshikawa, K. Gene expression within cell-sized lipid vesicles. ChemBioChem 2003, 4, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Stano, P.; Luisi, P.L. Protein synthesis in sub-micrometer water-In-oil droplets. Chembiochem 2015, 16, 2073–2079. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Murayama, Y.; Fujiwara, K.; Fujisawa, T.; Sasaki, S.; Kidoaki, S.; Yanagisawa, M. Increasing elasticity through changes in the secondary structure of gelatin by gelation in a microsized lipid space. ACS Cent. Sci. 2018, 4, 477–483. [Google Scholar] [CrossRef]
- Matthies, M.; Nayan, P.A.; Thorsten, L.S. Design and synthesis of triangulated DNA origami trusses. Nano. Lett. 2016, 16, 2108–2113. [Google Scholar] [CrossRef]
- Benson, E.; Mohammed, A.; Gardell, J.; Masich, S.; Czeizler, E.; Orponen, P.; Högberg, B. DNA rendering of polyhedral meshes at the nanoscale. Nature 2015, 523, 441–444. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Krainer, G.; Grundmeier, G.; Schlierf, M.; Keller, A. Structural stability of DNA origami nanostructures in the presence of chaotropic agents. Nanoscale 2016, 8, 10398–10405. [Google Scholar] [CrossRef]
- Dunn, K.E.; Dannenberg, F.; Ouldridge, T.E.; Kwiatkowska, M.; Turberfield, A.J.; Bath, J. Guiding the folding pathway of DNA origami. Nature 2015, 525, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Lee Tin Wah, J.; David, C.; Rudiuk, S.; Baigl, D.; Estévez-Torres, A. Observing and controlling the folding pathway of DNA origami at the nanoscale. ACS Nano. 2016, 10, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.; Möritz, N.; Dietz, H. The sequence of events during folding of a DNA origami. Sci. Adv. 2019, 5, eaaw1412. [Google Scholar] [CrossRef] [PubMed]
- Porras, M.; Solans, C.; González, C.; Martínez, A.; Guinart, A.; Gutiérrez, J.M. Studies of formation of W/O nano-emulsions. Coll. Surf. A: Physicochem. Eng. Asp. 2004, 249, 115–118. [Google Scholar]
- Weitz, M.; Kim, J.; Kapsner, K.; Winfree, E.; Franco, E.; Simmel, F.C. Diversity in the dynamical behaviour of a compartmentalized programmable biochemical oscillator. Nat. Chem. 2014, 6, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Komiya, K.; Kawamata, I.; Murata, S.; Shin-Ichiro, M.N. Isothermal amplification of specific DNA molecules inside giant unilamellar vesicles. Chem. Comm. 2019, 55, 9084–9087. [Google Scholar] [CrossRef] [PubMed]
- Douglas, S.M.; Marblestone, A.H.; Teerapittayanon, S.; Vazquez, A.; Church, G.M.; Shih, W.M. Rapid prototyping of 3D DNA-origami shapes with caDNAno. Nuc. Acids Res. 2009, 37, 5001–5006. [Google Scholar] [CrossRef]
- vHelix-Free-form DNA-nanostructure Design. Available online: http://www.vhelix.net/ (accessed on 19 August 2019).
- Stahl, E.; Martin, T.G.; Praetorius, F.; Dietz, H. Facile and scalable preparation of pure and dense DNA origami solutions. Angew. Chem. Int. Ed. 2014, 53, 12735–12740. [Google Scholar] [CrossRef]
- Muscat, R.A.; Bath, J.; Turberfield, A.J. A programmable molecular robot. Nano Lett. 2011, 11, 982–987. [Google Scholar] [CrossRef]
- Murata, S.; Konagaya, A.; Kobayashi, S.; Saito, H.; Hagiya, M. Molecular robotics: A new paradigm for artifacts. New Gen. Comp. 2013, 31, 27–45. [Google Scholar] [CrossRef]
- Sato, Y.; Hiratsuka, Y.; Kawamata, I.; Murata, S.; Nomura, S.-I.M. Micrometer-Sized molecular robot changes its shape in response to signal molecules. Sci. Robot. 2017, 2, eaal3735. [Google Scholar] [CrossRef]
Sample Availability: Samples of the DNA sequences are available online. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, T.; Sato, Y.; Otaka, H.; Kawamata, I.; Murata, S.; Nomura, S.-I.M. DNA Origami “Quick” Refolding inside of a Micron-Sized Compartment. Molecules 2020, 25, 8. https://doi.org/10.3390/molecules25010008
Watanabe T, Sato Y, Otaka H, Kawamata I, Murata S, Nomura S-IM. DNA Origami “Quick” Refolding inside of a Micron-Sized Compartment. Molecules. 2020; 25(1):8. https://doi.org/10.3390/molecules25010008
Chicago/Turabian StyleWatanabe, Taiki, Yusuke Sato, Hayato Otaka, Ibuki Kawamata, Satoshi Murata, and Shin-Ichiro M. Nomura. 2020. "DNA Origami “Quick” Refolding inside of a Micron-Sized Compartment" Molecules 25, no. 1: 8. https://doi.org/10.3390/molecules25010008
APA StyleWatanabe, T., Sato, Y., Otaka, H., Kawamata, I., Murata, S., & Nomura, S.-I. M. (2020). DNA Origami “Quick” Refolding inside of a Micron-Sized Compartment. Molecules, 25(1), 8. https://doi.org/10.3390/molecules25010008