The Role of Astragaloside IV against Cerebral Ischemia/Reperfusion Injury: Suppression of Apoptosis via Promotion of P62-LC3-Autophagy

Abstract
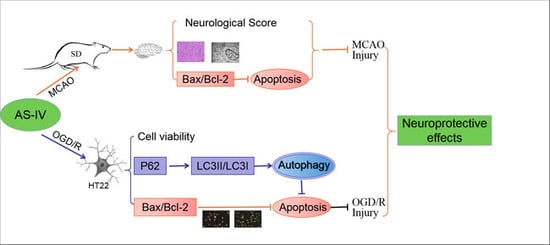
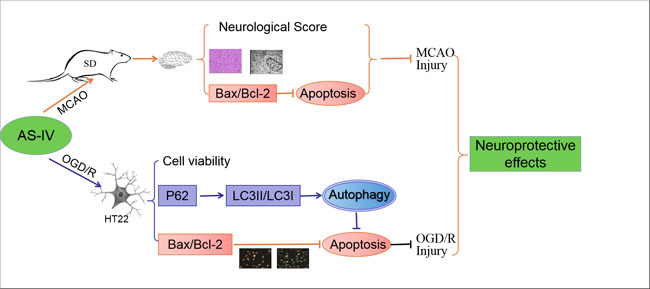{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
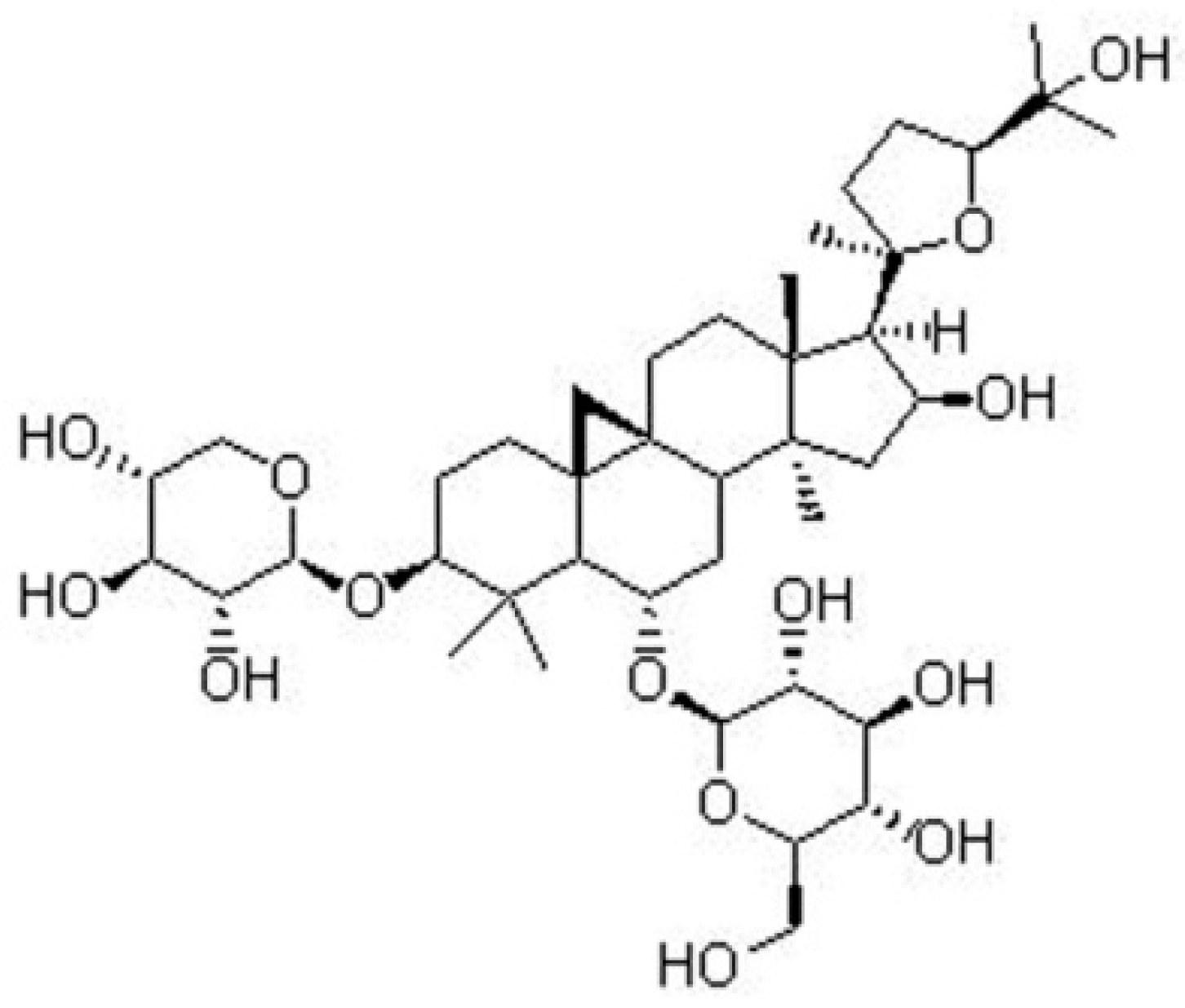
1. Introduction
2. Results
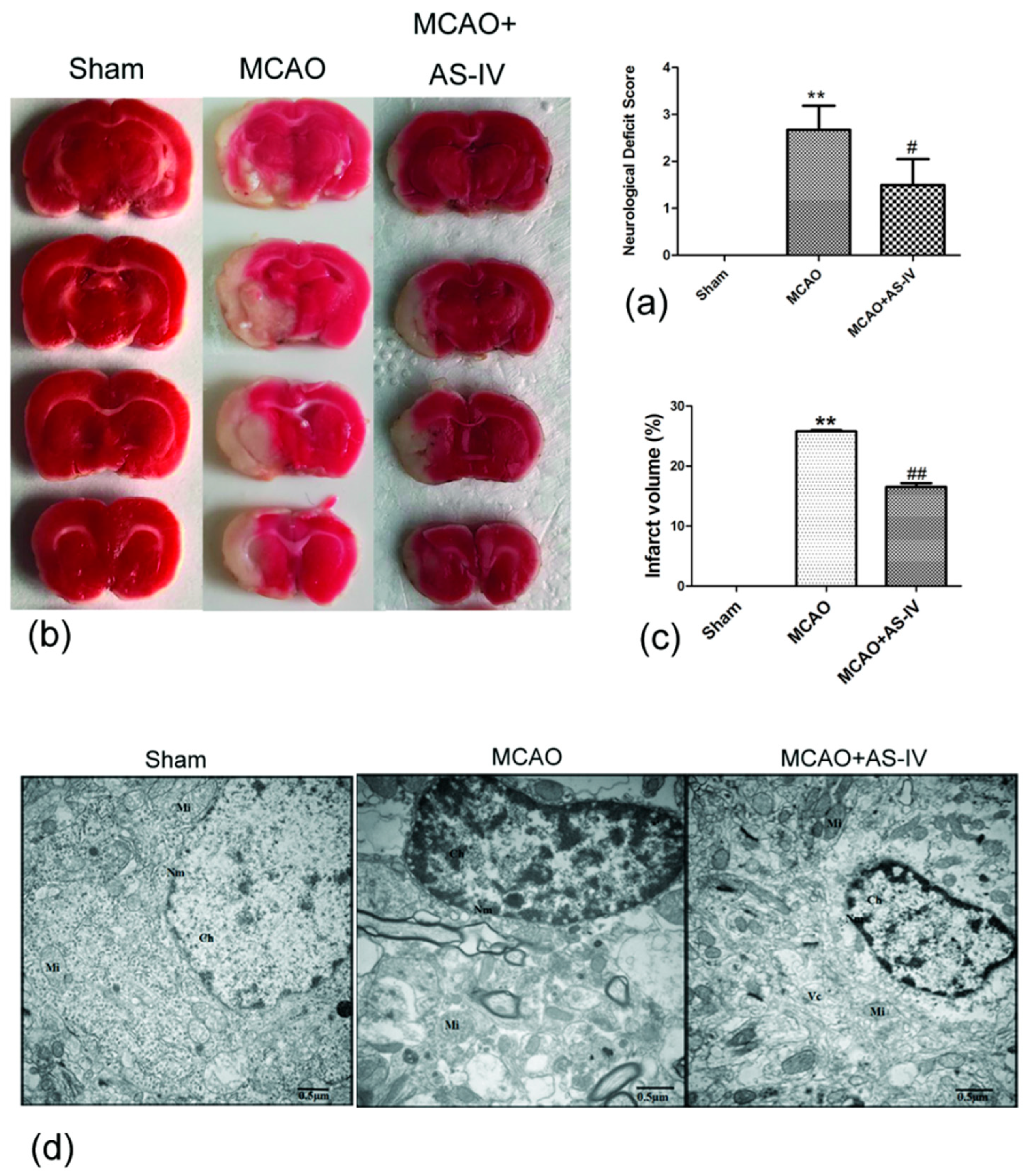
2.1. Astragaloside IV Inhibits Cerebral Damage following Ischemia/Reperfusion Injury
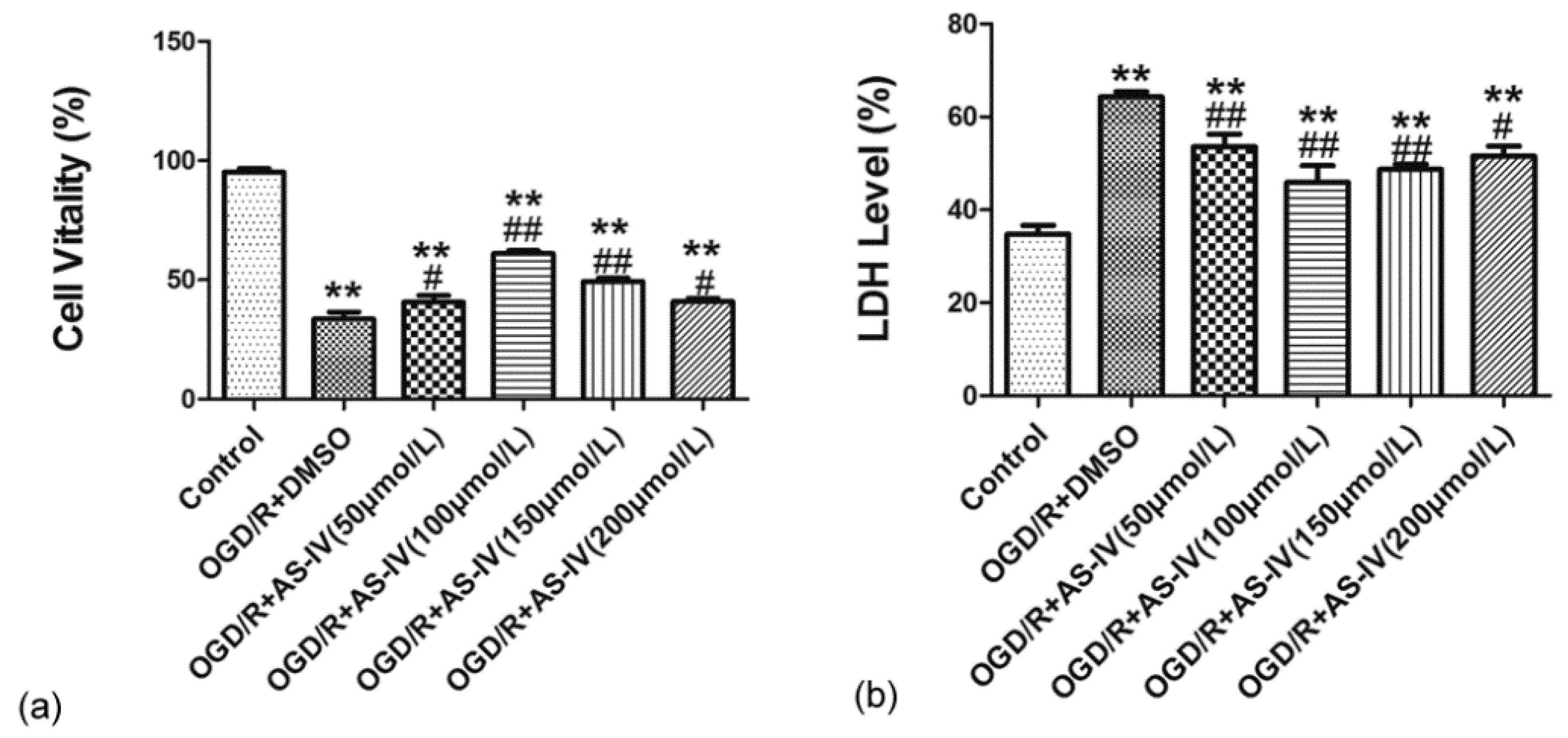
2.2. Astragaloside IV Increased the Viability of HT22 Cells after OGD/R
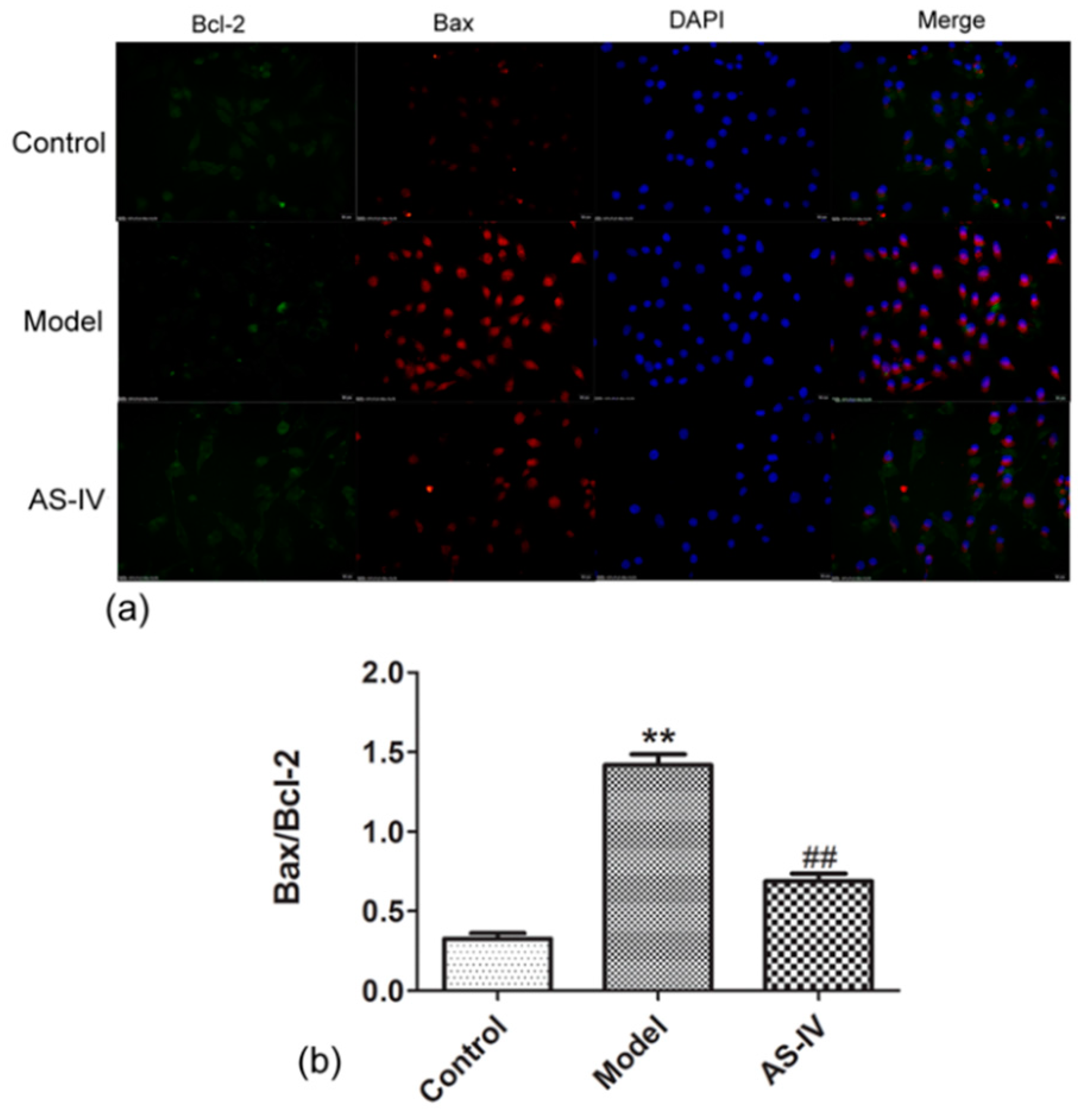
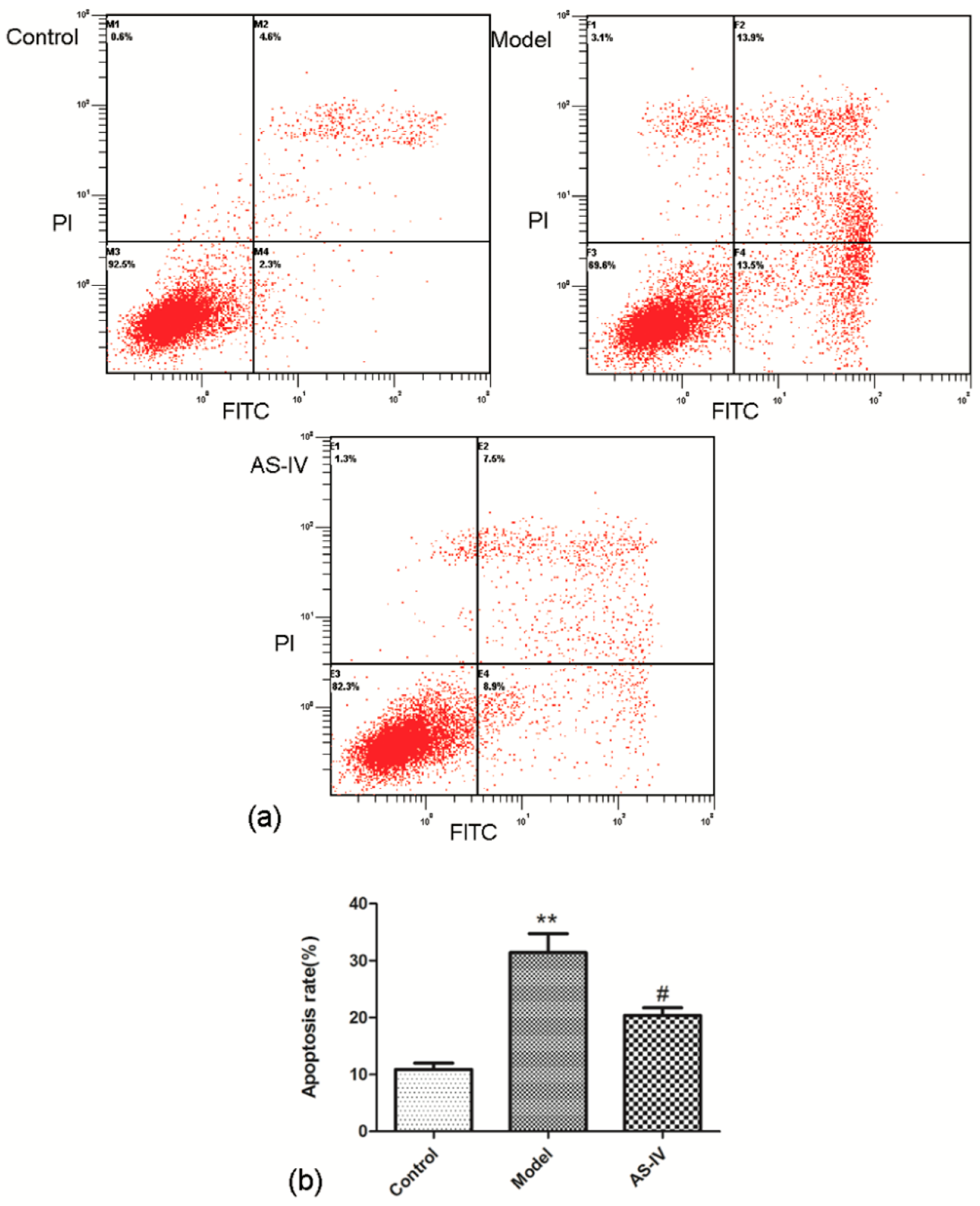
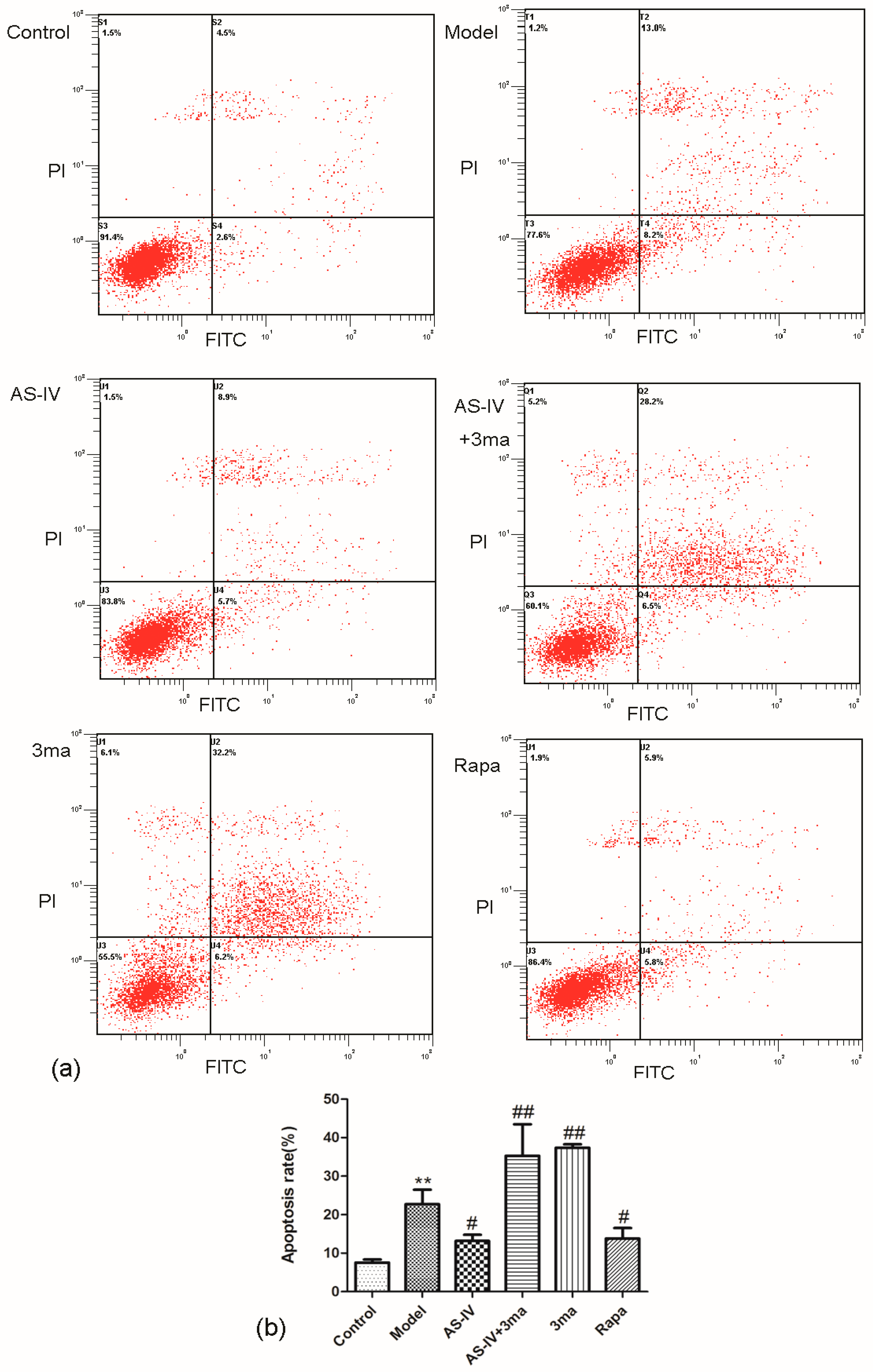
2.3. Astragaloside IV Reduced Apoptosis in OGD/R HT22 Cells
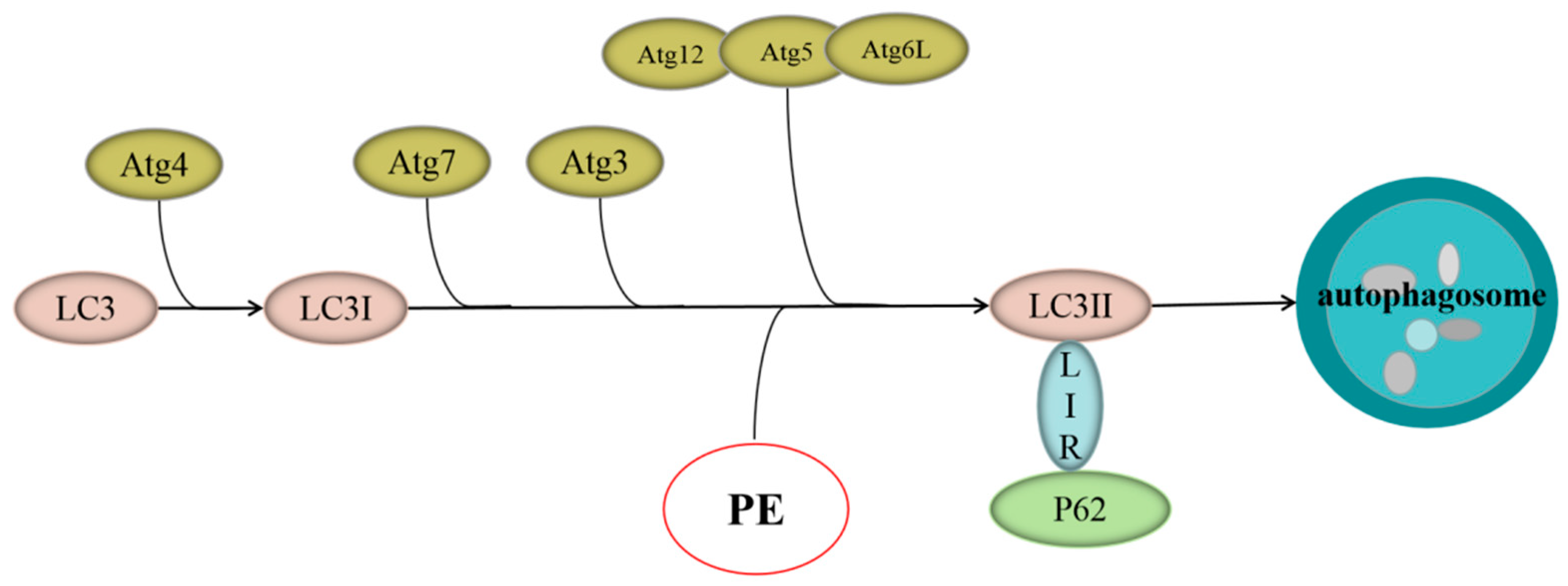
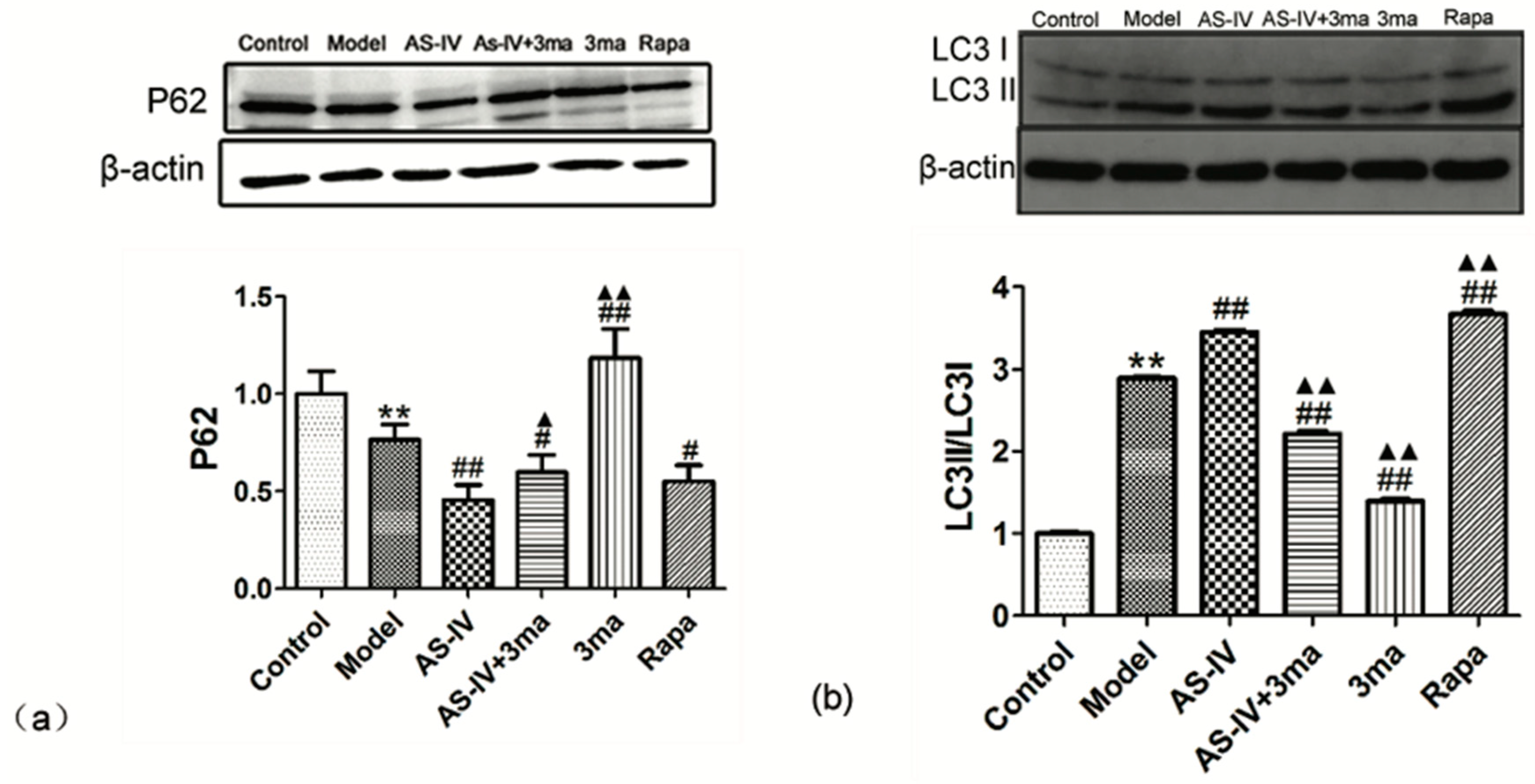
2.4. Astragaloside IV Enhanced Autophagy on HT22 Cells after OGD/R
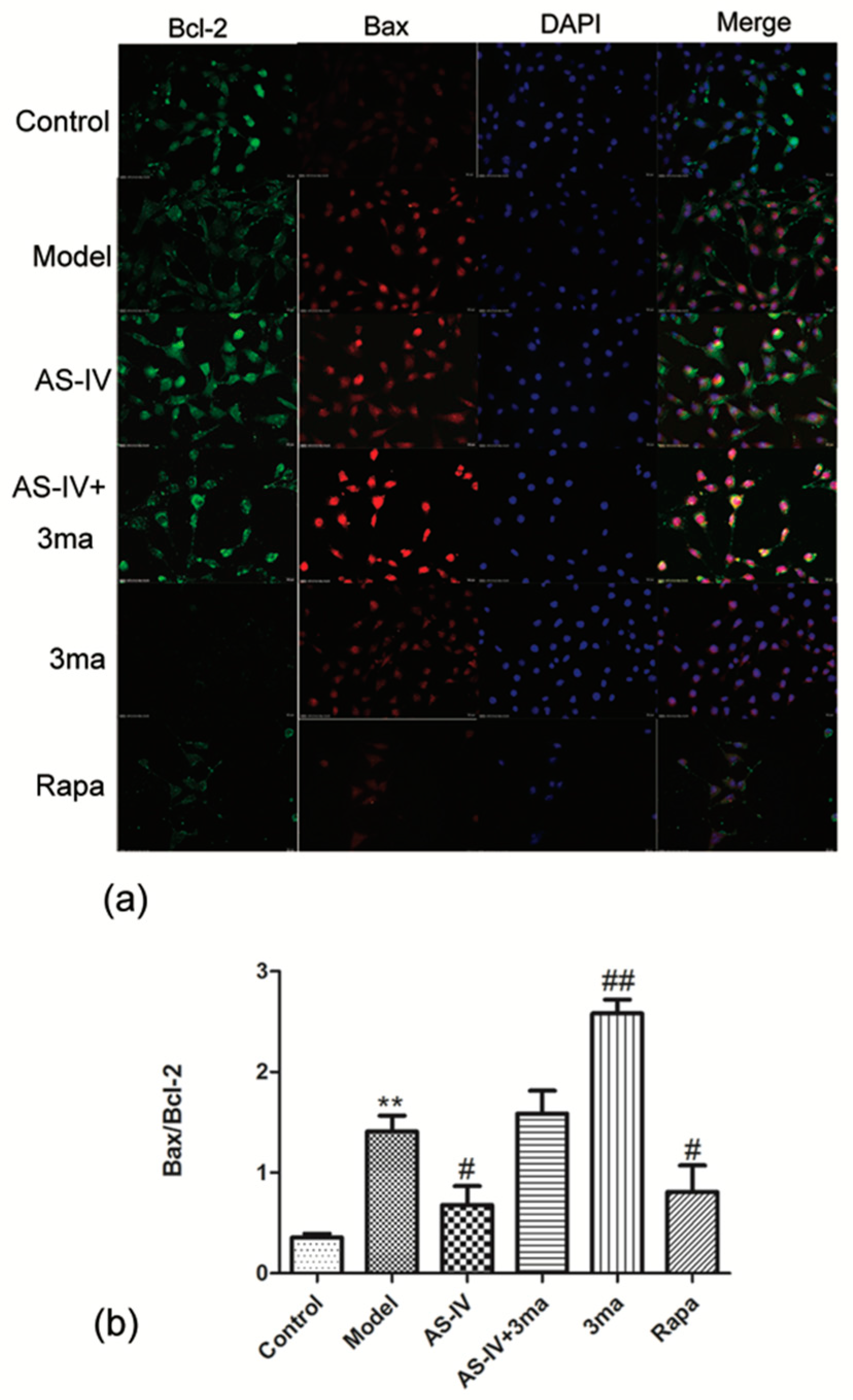
2.5. Astragaloside IV Inhibited Apoptosis on HT22 Cells after OGD/R by Activating Autophagy
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Middle Cerebral Artery Occlusion (MCAO)/Reperfusion (I/R) and Drug Treatment
4.3. Neurological Score Measurement
4.4. TTC Staining
4.5. Cell Culture
4.6. In Vitro Model of OGD/R
4.7. Cell Viability Analysis and Cytotoxicity Assay
4.8. Western Blot
4.9. Bax andBcl-2 Immunofluorescence Staining
4.10. Flow Cytometry
4.11. Data Analysis
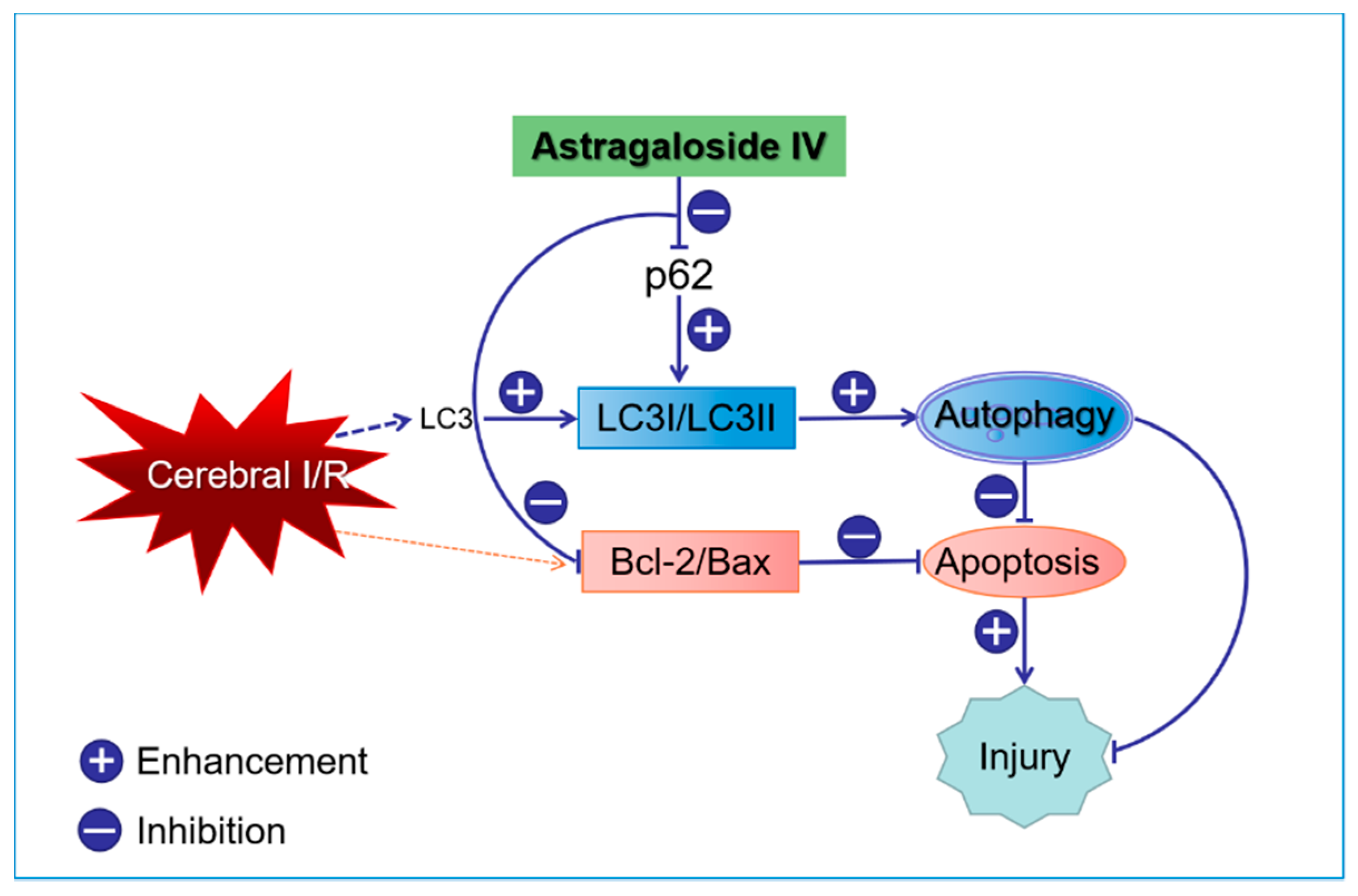
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Min, J.W.; Lu, L.; Freeling, J.L.; Martin, D.S.; Wang, H. USP14 inhibitor attenuates cerebral ischemia/reperfusion-induced neuronal injury in mice. J. Neurochem. 2017, 140, 826–833. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, H.; Kai, J.; Zhu, F.; Dong, J.; Xu, Z.; Wong, M.; Zeng, L.H. Rapamycin prevents cerebral stroke by modulating apoptosis and autophagy in penumbra in rats. Ann. Clin. Transl. Neurol. 2018, 5, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kong, X.; Xiu, H.; Dou, Y.; Wu, Z.; Sun, P. Combination of curcumin and vagus nerve stimulation attenuates cerebral ischemia/reperfusion injury-induced behavioral deficits. Biomed. Pharmacother. 2018, 103, 614–620. [Google Scholar] [CrossRef]
- Khandelwal, P.; Yavagal, D.R.; Sacco, R.L. Acute ischemic stroke intervention. J. Am. Coll. Cardiol. 2016, 67, 2631–2644. [Google Scholar]
- Liu, Y.; Yang, H.; Jia, G.; Li, L.; Chen, H.; Bi, J.; Wang, C. The synergistic neuroprotective effects of combined rosuvastatin and resveratrol pretreatment against cerebral ischemia/reperfusion injury. J. Stroke Cerebrovasc. Dis. 2018, 27, 1697–1704. [Google Scholar] [CrossRef]
- Sheng, R.; Liu, X.Q.; Zhang, L.S.; Gao, B.; Han, R.; Wu, Y.Q.; Zhang, X.Y.; Qin, Z.H. Autophagy regulates endoplasmic reticulum stress in ischemic preconditioning. Autophagy 2012, 8, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.G.; Tao, W.; Yang, S.B.; Wang, J.F.; Mei, Z.G.; Feng, Z.T. Autophagy: Novel insights into therapeutic target of electro acupuncture against cerebral ischemia/ reperfusion injury. Neural Regen Res. 2019, 14, 954–961. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, T.; Zhang, Y.; Li, J.; Jin, L.; Sun, Y.; Shi, N.; Liu, K.; Sun, X. Ischemic post conditioning alleviates cerebral ischemia-reperfusion injury through activating autophagy during early reperfusion in rats. Neurochem. Res. 2018, 43, 1826–1840. [Google Scholar] [CrossRef]
- Wang, M.; Li, Y.J.; Ding, Y.; Zhang, H.N.; Sun, T.; Zhang, K.; Yang, L.; Guo, Y.Y.; Liu, S.B.; Zhao, M.G.; et al. Silibinin prevents autophagic cell death upon oxidative stress in cortical neurons and cerebral ischemia-reperfusion injury. Mol. Neurobiol. 2016, 53, 932–943. [Google Scholar] [CrossRef]
- Oh, S.H.; Choi, Y.B.; Kim, J.H.; Weihl, C.C.; Ju, J.S. Quantification of autophagy flux using LC3 ELISA. Anal. Biochem. 2017, 530, 57–67. [Google Scholar] [CrossRef]
- Wang, J.; Garbutt, C.; Ma, H.; Gao, P.; Hornicek, F.J.; Kan, Q.; Shi, H.; Duan, Z. Expression and role of autophagy-associated p62 (SQSTM1) in multidrug resistant ovarian cancer. Gynecol. Oncol. 2018, 150, 143–150. [Google Scholar] [CrossRef]
- Li, H.; Wu, J.; Shen, H.; Yao, X.; Liu, C.; Pianta, S.; Han, J.; Borlongan, C.V.; Chen, G. Autophagy in hemorrhagic stroke: Mechanisms and clinical implications. Prog. Neurobiol. 2018, 163–164, 79–97. [Google Scholar] [CrossRef]
- Wong, W. Focus Issue: Autophagy as hero and villain. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef]
- Goodall, M.L.; Fitzwalter, B.E.; Zahedi, S.; Wu, M.; Rodriguez, D.; Mulcahy-Levy, J.M.; Green, D.R.; Morgan, M.; Cramer, S.D.; Thorburn, A. The autophagy machinery controls cell death switching between apoptosis and necroptosis. Dev. Cell 2016, 37, 337–349. [Google Scholar] [CrossRef]
- Qin, L.; Wang, Z.; Tao, L.; Wang, Y. ER stress negatively regulates AKT/TSC/mTOR pathway to enhance autophagy. Autophagy 2010, 6, 239–247. [Google Scholar] [CrossRef]
- Kalinichenko, S.G.; Matveeva, N. Morphological characteristic of apoptosis and its significance in neurogenesis. Morfologiia 2007, 131, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Chen, R.; Chen, X.; Xie, Z.; Huo, F.; Wu, Z. Che-1 inhibits oxygen-glucose deprivation/reoxygenation-induced neuronal apoptosis associated with inhibition of the p53-mediated proapoptotic signaling pathway. Neuroreport 2018, 29, 1193–1200. [Google Scholar] [CrossRef]
- Hu, G.Q.; Du, X.; Li, Y.J.; Gao, X.Q.; Chen, B.Q.; Yu, L. Inhibition of cerebral ischemia/reperfusion injury-induced apoptosis: Nicotiflorin and JAK2/STAT3 pathway. Neural Regen Res. 2017, 12, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Song, F.; Lu, D.Y.; Dong, L.; Wu, Q.; Liu, Z.Y.; Li, Y.J.; Wang, A.M. Anti-oxidation and anti-apoptosis mechanism of Xinshao formula on cerebral ischemia reperfusion injury. ZhongguoZhong Yao ZaZhi 2018, 43, 3519–3524. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Xue, L.; Hao, C.; Liao, W.; Wan, Q. Treatment with enriched environment reduces neuronal apoptosis in the periinfarct cortex after cerebral ischemia/reperfusion injury. Cell. Physiol. Biochem. 2017, 41, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Tao, T.; Li, C.L.; Yang, W.C.; Zeng, X.Z.; Song, C.Y.; Yue, Z.Y.; Dong, H.; Qian, H. Protective effects of propofol against whole cerebral ischemia/reperfusion injury in rats through the inhibition of the apoptosis-inducing factor pathway. Brain Res. 2016, 1644, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef]
- Liu, Q.S.; Deng, R.; Li, S.; Li, X.; Li, K.; Kebaituli, G.; Li, X.; Liu, R. Ellagic acid protects against neuron damage in ischemic stroke through regulating the ratio of Bcl-2/Bax expression. Appl. Physiol. Nutr. Metab. 2017, 42, 855–860. [Google Scholar] [CrossRef]
- Zhang, W.W.; Xu, F.; Wang, D.; Ye, J.; Cai, S.Q. BuyangHuanwu decoction ameliorates ischemic stroke by modulating multiple targets with multiple components: In vitro evidences. Chin. J. Nat. Med. 2018, 16, 194–202. [Google Scholar] [CrossRef]
- Zheng, X.W.; Shan, C.S.; Xu, Q.Q.; Wang, Y.; Shi, Y.H.; Wang, Y.; Zheng, G.Q. BuyangHuanwudecoction targets SIRT1/VEGF pathway to promote angiogenesis after cerebral ischemia/reperfusion injury. Front. Neurosci. 2018, 12, 911. [Google Scholar] [CrossRef]
- Yang, J.; Gao, F.; Zhang, Y.; Liu, Y.; Zhang, D. BuyangHuanwudecoction (BYHWD) enhances angiogenic effect of mesenchymal stem cell by upregulating VEGF expression after focal cerebral ischemia. J. Mol. Neurosci. 2015, 56, 898–906. [Google Scholar] [CrossRef]
- Jinglong, T.; Weijuan, G.; Jun, L.; Tao, Q.; Hongbo, Z.; Shasha, L. The molecular and electrophysiological mechanism of BuyangHuanwudecoction in learning and memory ability of vascular dementia rats. Brain Res. Bull. 2013, 99, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Auyeung, K.K.; Han, Q.B.; Ko, J.K. Astragalus membranaceus: A review of its protection against inflammation and gastrointestinal cancers. Am. J. Chin. Med. 2016, 44, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.Z.; Wu, F.; Shen, J.; Lu, L.; Fu, D.L.; Liao, W.J.; Zheng, G.Q. Clinical efficacy and safety of buyanghuanwu decoction for acute ischemic stroke: A systematic review and meta-analysis of 19 randomized controlled trials. Evid.-Based Complement. Altern. Med. 2012, 2012, 630124. [Google Scholar] [CrossRef]
- Liu, S.S.; Gao, W.J.; Qian, T.; Zhang, X. Effect of astragalus injection on expression of JNK3 in hippocampus of cerebral ischemia reperfusion rats. Chin. Pharmacol. Bull. 2012, 5, 78–83. [Google Scholar]
- Ip, F.C.; Zhao, Y.M.; Chan, K.W.; Cheng, E.Y.; Tong, E.P.; Chandrashekar, O.; Fu, G.M.; Zhao, Z.Z.; Ip, N.Y. Neuroprotective effect of a novel Chinese herbal decoction on cultured neurons and cerebral ischemic rats. BMC Complement. Altern. Med. 2016, 16, 437. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Guo, R.J.; Wang, J.L.; He, L.J.; Liu, X.Z.; Wang, J.H. Literature research of medication rule in treatment of apoplexy with qi deficiency and blood stasis syndrome. J. Trad. Chin. Med. 2018, 59, 69–73. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhou, Q.H.; Xu, M.B.; Zhou, X.L.; Zheng, G.Q. Astragaloside IV for experimental focal cerebral ischemia: Preclinical evidence and possible mechanisms. Oxid. Med. Cell. Longev. 2017, 2017, 8424326. [Google Scholar] [CrossRef]
- Huang, X.P.; Tan, H.; Chen, B.Y.; Deng, C.Q. Astragalus extract alleviates nerve injury after cerebral ischemia by improving energy metabolism and inhibiting apoptosis. Biol. Pharm. Bull. 2012, 35, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebralartery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xue, X.; Xian, L.; Guo, Z.; Ito, Y.; Sun, W. Potential neuroprotection of protodioscin against cerebral ischemia-reperfusion injury in rats through intervening inflammation and apoptosis. Steroids 2016, 113, 52–63. [Google Scholar] [CrossRef]
- Chen, C.M.; Wu, C.T.; Yang, T.H.; Liu, S.H.; Yang, F.Y. Preventive effect of low intensity pulsed ultrasound against experimental cerebral ischemia/reperfusion injury via apoptosis reduction and brain-derived neurotrophic factor induction. Sci. Rep. 2018, 8, 5568. [Google Scholar] [CrossRef]
- Kalogeris, T.; Baines, C.P.; Krenz, M.; Korthuis, R.J. Cell biology of ischemia/reperfusion injury. Int. Rev. Cell Mol. Biol. 2012, 298, 229–317. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Zhang, H.; Mu, Y. Pharmacological effects of Astragaloside IV: A literature review. J. Trad. Chin. Med 2013, 33, 413–416. [Google Scholar] [CrossRef]
- Li, L.; Hou, X.; Xu, R.; Liu, C.; Tu, M. Research review on the pharmacological effects of astragaloside IV. Fundam. Clin. Pharmacol. 2017, 31, 17–36. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.F.; Zhang, Y.; Zhou, X.H.; Wu, M.S.; Zhao, Y.M.; Gao, W.J. Effects of astragaloside IV on apoptosis of PC12 cells induced by hypoxia/hypoglycemia and reoxygenation. Chin. Pharmacol. Bull. 2016, 32, 1411–1415. [Google Scholar]
- Ji, H.J.; Hu, J.F.; Wang, Y.H.; Chen, X.Y.; Zhou, R.; Chen, N.H. Osthole improves chronic cerebral hypoperfusion induced cognitive deficits and neuronal damage in hippocampus. Eur. J. Pharmacol 2010, 636, 96–101. [Google Scholar] [CrossRef]
- Hatok, J.; Racay, P. Bcl-2 family proteins: Master regulators of cell survival. Biomol. Concepts 2016, 7, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.M.; Kim, Y.R.; Kim, H.N.; Choi, Y.W.; Lee, J.W.; Kim, C.M.; Baek, J.U.; Shin, H.K.; Choi, B.T. Erratum to: Neuroprotection and spatial memory enhancement of four herbal mixture extract in HT22 hippocampal cells and a mouse model of focal cerebral ischemia. BMC Complement. Altern. Med. 2015, 15, 276. [Google Scholar] [CrossRef]
- Shimada, S.; Hirabayashi, M.; Ishige, K.; Kosuge, Y.; Kihara, T.; Ito, Y. Activation of dopamine D4 receptors is protective against hypoxia/reoxygenation-induced cell death in HT22 cells. J. Pharmacol. Sci. 2010, 114, 217–224. [Google Scholar] [CrossRef]
- Chumboatong, W.; Thummayot, S.; Govitrapong, P.; Tocharus, C.; Jittiwat, J.; Tocharus, J. Neuroprotection of agomelatine against cerebral ischemia/reperfusion injury through an antiapoptotic pathway in rat. Neurochem. Int. 2017, 102, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Wicha, P.; Tocharus, J.; Janyou, A. Hexahydrocurcumin protects against cerebral ischemia/reperfusion injury, attenuates inflammation, and improves antioxidant defenses in a rat stroke model. PLoS ONE 2017, 12, e0189211. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.T.; Warr, M.R.; Adelman, E.R.; Lansinger, O.M.; Flach, J.; Verovskaya, E.V.; Figueroa, M.E.; Passegue, E. Autophagy maintains the metabolism and function of young and old stem cells. Nature 2017, 543, 205–210. [Google Scholar] [CrossRef]
- Carloni, S.; Buonocore, G.; Balduini, W. Protective role of autophagy in neonatal hypoxia-ischemia induced brain injury. Neurobiol. Dis. 2008, 32, 329–339. [Google Scholar] [CrossRef]
- Gump, J.M.; Staskiewicz, L.; Morgan, M.J.; Bamberg, A.; Riches, D.W.; Thorburn, A. Autophagy variation within a cell population determines cell fate through selective degradation of Fap-1. Nat. Cell Biol. 2014, 16, 47–54. [Google Scholar] [CrossRef]
- Takahashi, A.; Takabatake, Y.; Kimura, T.; Maejima, I.; Namba, T.; Yamamoto, T.; Matsuda, J.; Minami, S.; Kaimori, J.Y.; Matsui, I.; et al. Autophagy Inhibits the Accumulation of Advanced Glycation End Products by Promoting Lysosomal Biogenesis and Function in the Kidney Proximal Tubules. Diabetes 2017, 66, 1359–1372. [Google Scholar] [CrossRef]
- Sheng, R.; Zhang, L.S.; Han, R.; Liu, X.Q.; Gao, B.; Qin, Z.H. Autophagy activation is associated with neuroprotection in a rat model of focal cerebral ischemic preconditioning. Autophagy 2010, 6, 482–494. [Google Scholar] [CrossRef]
- Song, H.; Yan, C.; Tian, X.; Zhu, N.; Li, Y.; Liu, D.; Liu, Y.; Liu, M.; Peng, C.; Zhang, Q.; et al. CREG protects from myocardial ischemia/reperfusion injury by regulating myocardial autophagy and apoptosis. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1893–1903. [Google Scholar] [CrossRef]
- Kim, J.H.; Hong, S.K.; Wu, P.K.; Richards, A.L.; Jackson, W.T.; Park, J.I. Raf/MEK/ERK can regulate cellular levels of LC3B and SQSTM1/p62 at expression levels. Exp. Cell Res. 2014, 327, 340–352. [Google Scholar] [CrossRef]
- Liu, J.; Meng, Q.; Jing, H.; Zhou, S. Astragaloside IV protects against apoptosis in human degenerative chondrocytes through autophagy activation. Mol. Med. Rep. 2017, 16, 3269–3275. [Google Scholar] [CrossRef]
- Wang, X.; Gao, Y.; Tian, N.; Zhu, Z.; Wang, T.; Xu, J.; Wu, B.; Zhang, N. Astragaloside IV represses high glucose-induced mesangial cells activation by enhancing autophagy via SIRT1 deacetylation of NF-kappaB p65 subunit. Drug Des. Dev. Ther. 2018, 12, 2971–2980. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Gao, H.; Tao, L.; Zhang, Y.; Zhai, J.; Sun, J.; Song, Y.; Zhang, S. Astragaloside IV protects against cisplatin-induced liver and kidney injury via autophagy-mediated inhibition of NLRP3 in rats. J. Toxicol. Sci. 2019, 44, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Song, Q.; Li, X.; Li, D.; Zhang, Q.; Meng, W.; Zhao, Q. Neuroprotective effects of Kukoamine A on neurotoxin-induced Parkinson’s model through apoptosis inhibition and autophagy enhancement. Neuropharmacology 2017, 117, 352–363. [Google Scholar] [CrossRef] [PubMed]
- Carloni, S.; Girelli, S.; Scopa, C.; Buonocore, G.; Longini, M.; Balduini, W. Activation of autophagy and Akt/CREB signaling play an equivalent role in the neuroprotective effect of rapamycin in neonatal hypoxia-ischemia. Autophagy 2010, 6, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Qu, Y.Z.; Zhao, Z.W.; Wu, S.X.; Liu, Y.Y.; Wei, X.Y.; Gao, L.; Gao, G.D. Astragaloside IV protects against focal cerebral ischemia/reperfusion injury correlating to suppression of neutrophils adhesion-related molecules. Neurochem. Int. 2012, 60, 458–465. [Google Scholar] [CrossRef]
- Xing, Y.; Wang, M.M.; Feng, Y.S.; Dong, F.; Zhang, F. Possible involvement of PTEN signaling pathway in the anti-apoptotic effect of electroacupuncture following ischemic stroke in rats. Cell. Mol. Neurobiol. 2018, 38, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Li, M.; Huang, Q.; Han, F.; Gu, J.H.; Xie, J.; Han, R.; Qin, Z.H.; Zhou, Z. Ischemia/reperfusion-induced upregulation of TIGAR in brain is mediated by SP1 and modulated by ROS and hormones involved in glucose metabolism. Neurochem. Int. 2015, 80, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Moran, J.; Perez-Basterrechea, M.; Garrido, P.; Diaz, E.; Alonso, A.; Otero, J.; Colado, E.; Gonzalez, C. Effects of estrogen and phytoestrogen treatment on an in vitro model of recurrent stroke on HT22 neuronal cell line. Cell. Mol. Neurobiol. 2017, 37, 405–416. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors.. |










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, Y.; Jin, X.-f.; Zhou, X.-h.; Dong, X.-h.; Yu, W.-t.; Gao, W.-j. The Role of Astragaloside IV against Cerebral Ischemia/Reperfusion Injury: Suppression of Apoptosis via Promotion of P62-LC3-Autophagy. Molecules 2019, 24, 1838. https://doi.org/10.3390/molecules24091838
Zhang Y, Zhang Y, Jin X-f, Zhou X-h, Dong X-h, Yu W-t, Gao W-j. The Role of Astragaloside IV against Cerebral Ischemia/Reperfusion Injury: Suppression of Apoptosis via Promotion of P62-LC3-Autophagy. Molecules. 2019; 24(9):1838. https://doi.org/10.3390/molecules24091838
Chicago/Turabian StyleZhang, Yi, Ying Zhang, Xiao-fei Jin, Xiao-hong Zhou, Xian-hui Dong, Wen-tao Yu, and Wei-juan Gao. 2019. "The Role of Astragaloside IV against Cerebral Ischemia/Reperfusion Injury: Suppression of Apoptosis via Promotion of P62-LC3-Autophagy" Molecules 24, no. 9: 1838. https://doi.org/10.3390/molecules24091838
APA StyleZhang, Y., Zhang, Y., Jin, X.-f., Zhou, X.-h., Dong, X.-h., Yu, W.-t., & Gao, W.-j. (2019). The Role of Astragaloside IV against Cerebral Ischemia/Reperfusion Injury: Suppression of Apoptosis via Promotion of P62-LC3-Autophagy. Molecules, 24(9), 1838. https://doi.org/10.3390/molecules24091838