
Evaluation of Biological Response of Lettuce (Lactuca sativa L.) and Weeds to Safranal Allelochemical of Saffron (Crocus sativus) by Using Static Exposure Method


 and
and 
Abstract
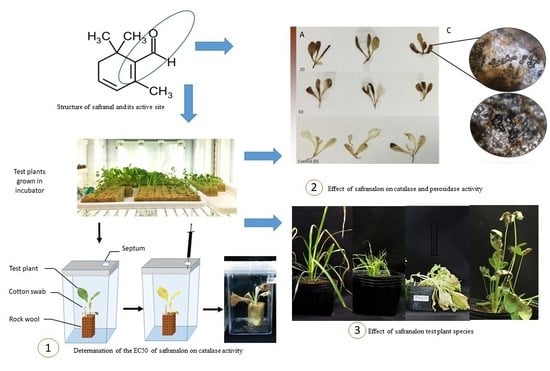
1. Introduction
2. Results and Discussion
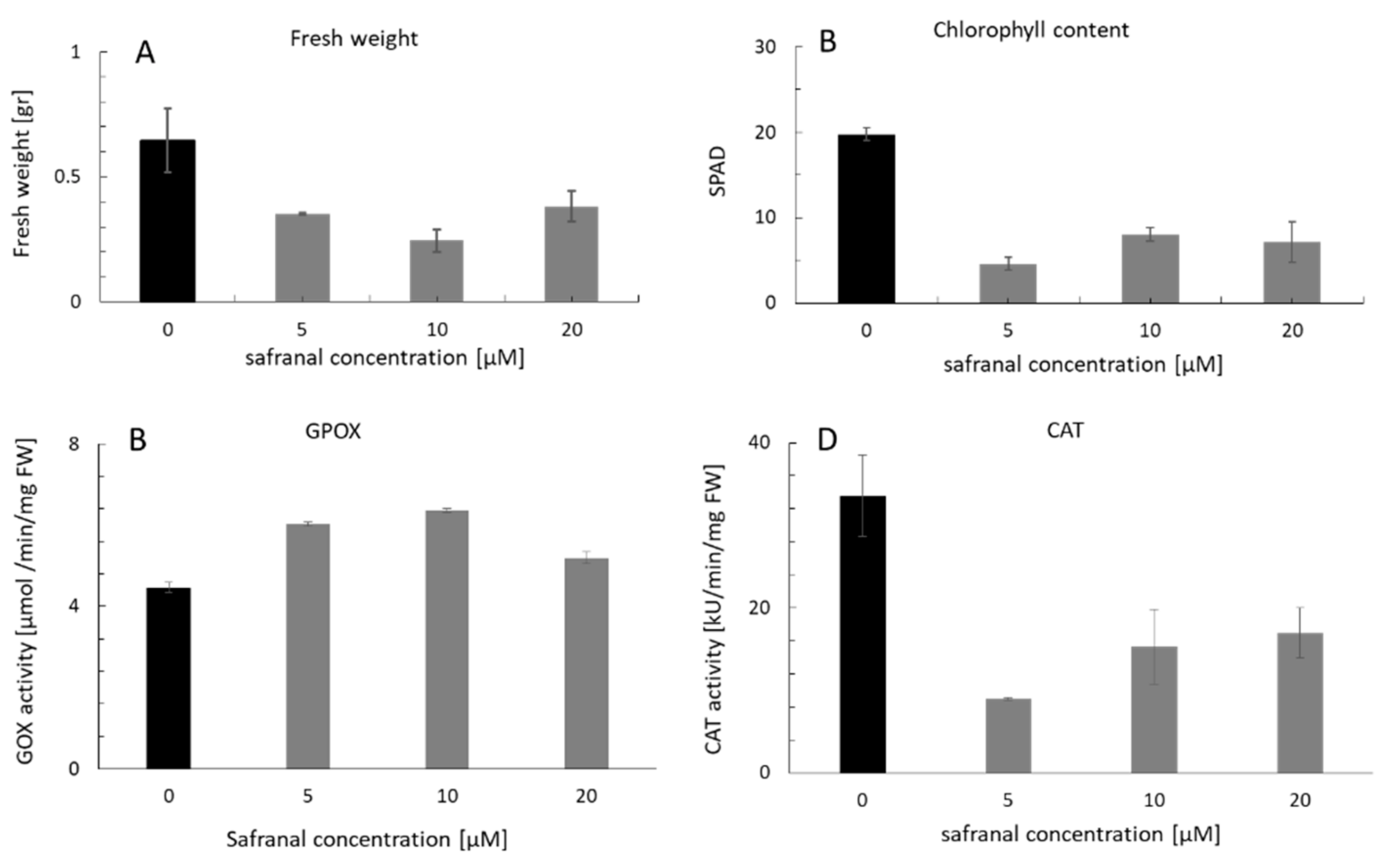
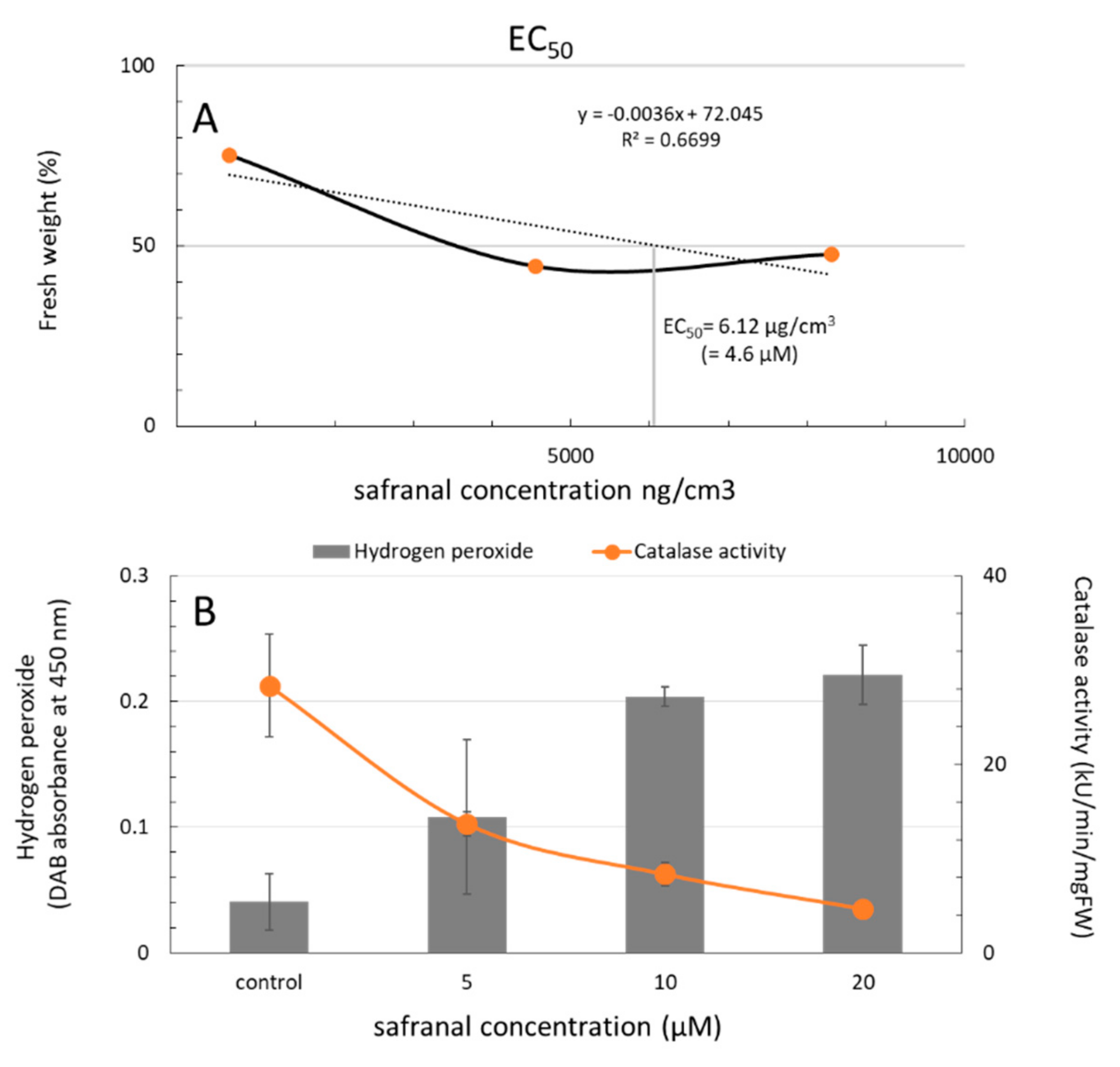
2.1. Physiological Response of L. sativa to the Volatile Safranal
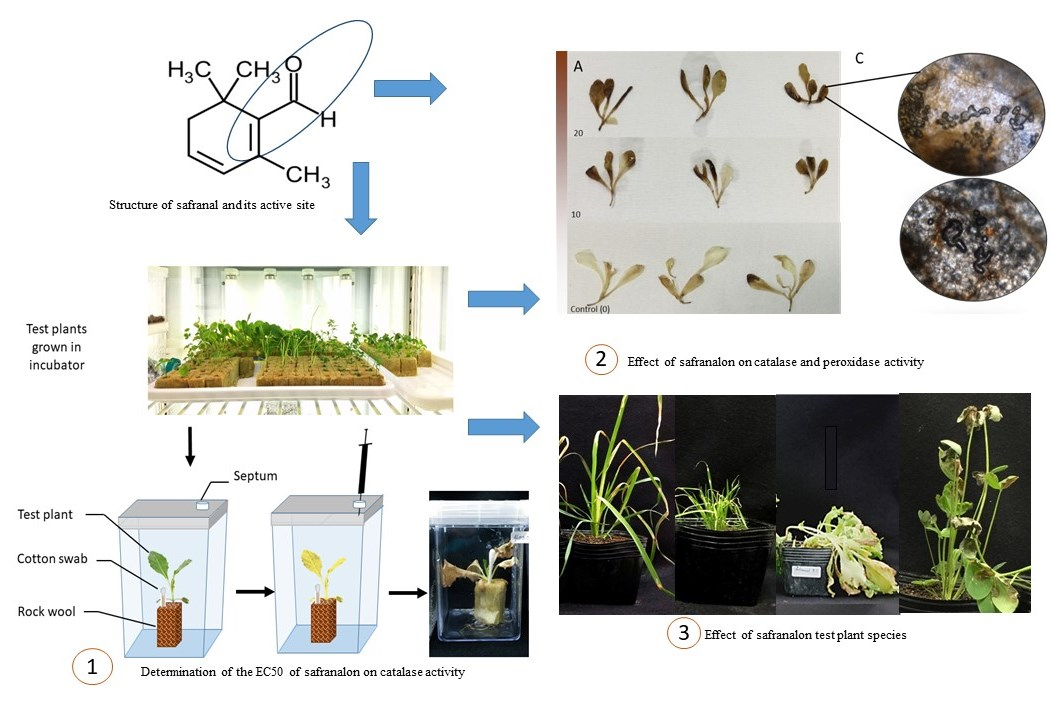
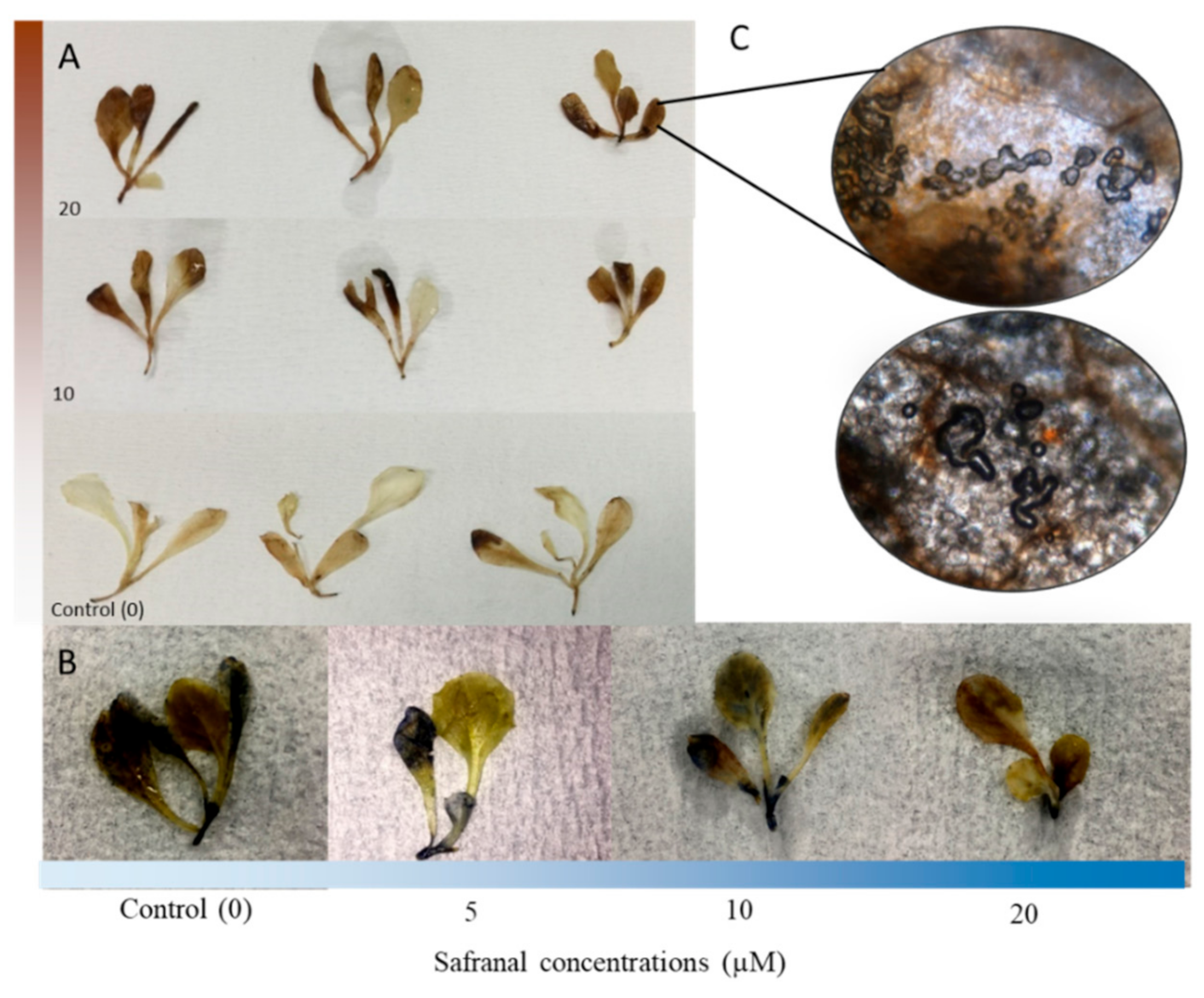
2.2. The Effect of Safranal on the Activity of CAT and GPOX, and Accumulation of Ros in Plant Tissues
2.2.1. Accumulation of ROS in Plant Tissues
2.2.2. CAT and GPOX Enzyme Assay Results

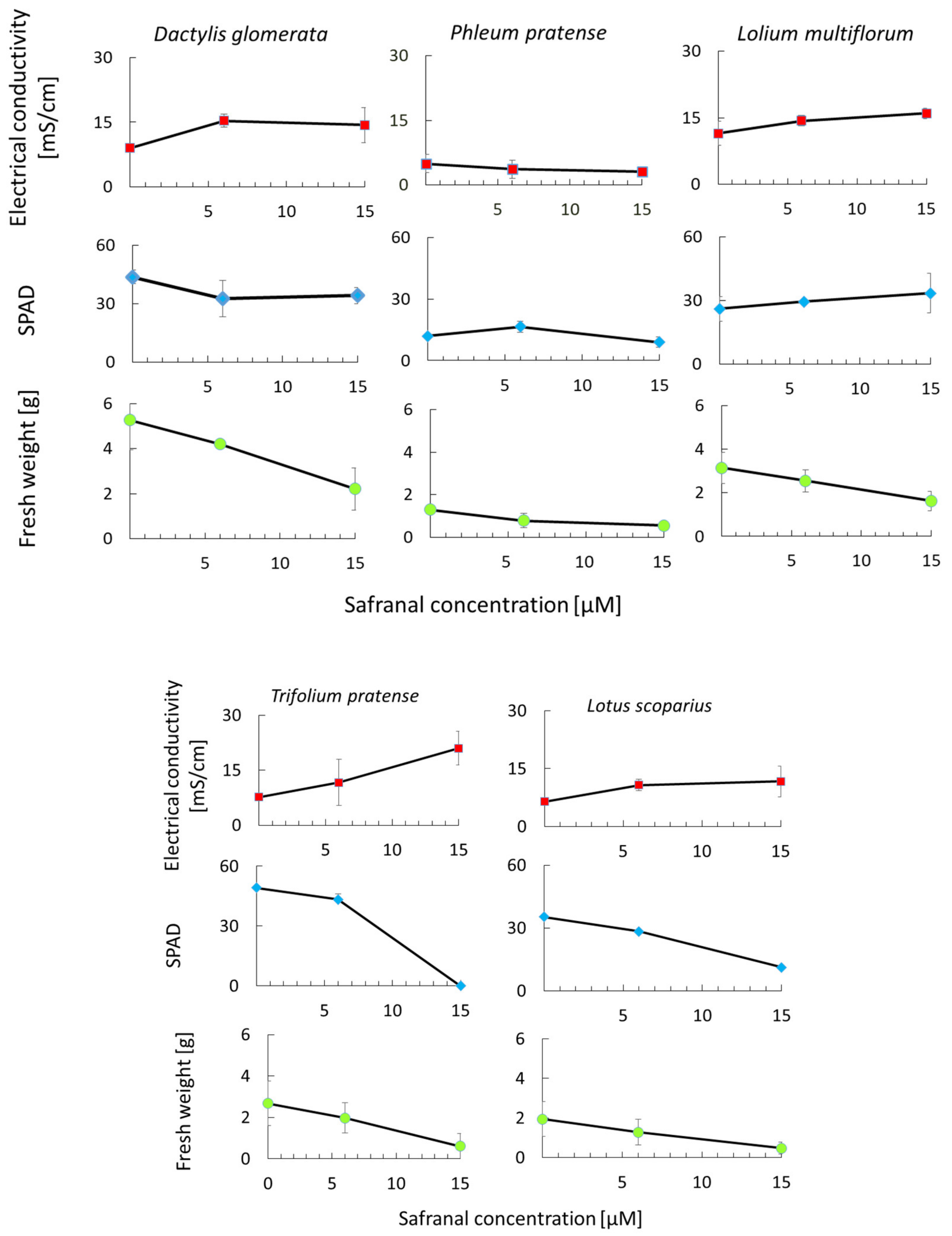
2.3. The Effect of Safranal on Growth and Physiological Characteristics of Weeds
3. Materials and Methods
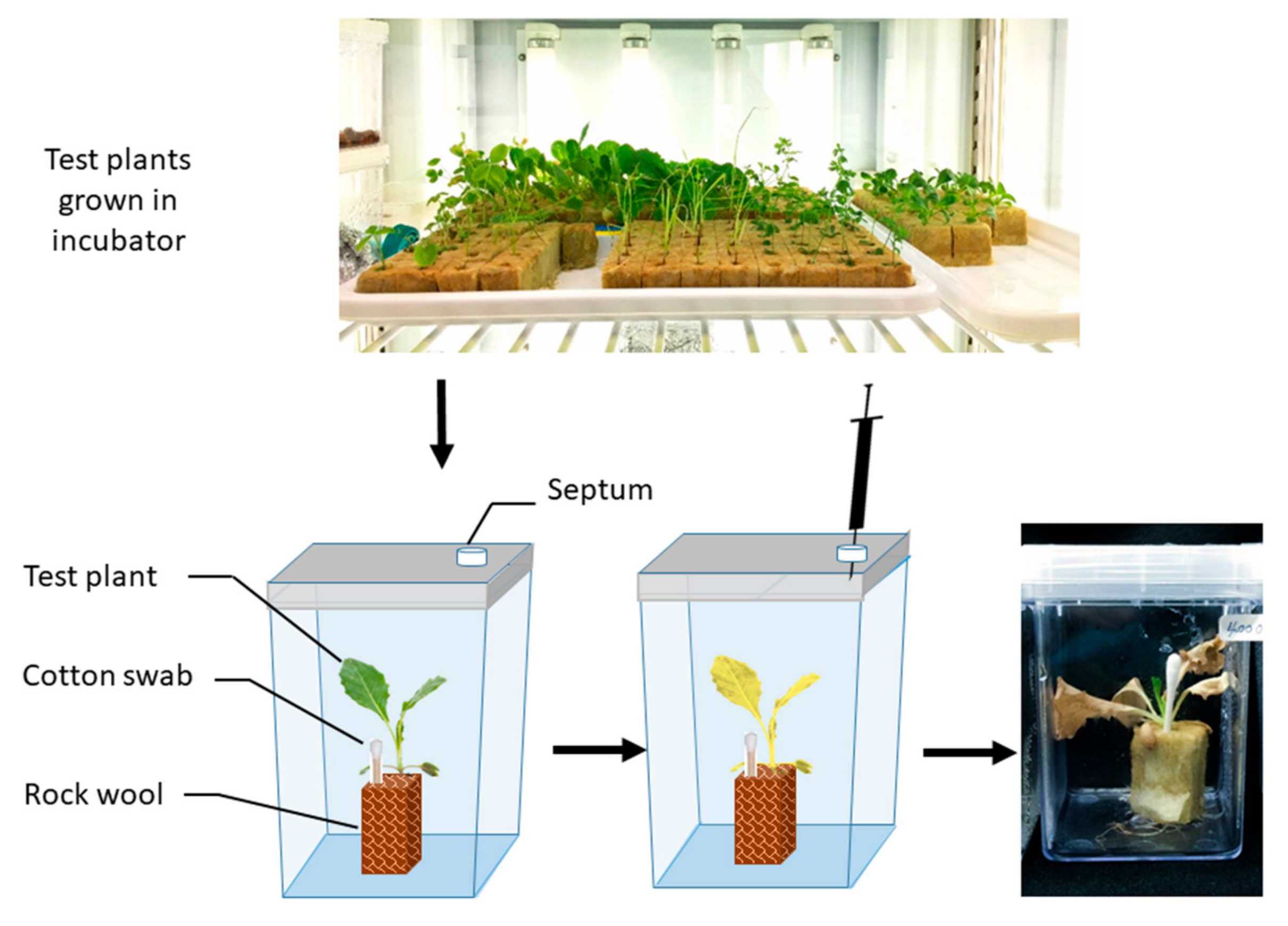
3.1. The Allelopathic Activity of Safranal on Growth and ROS Scavenging Enzymes of L. sativa in Vitro Condition
Plant Material
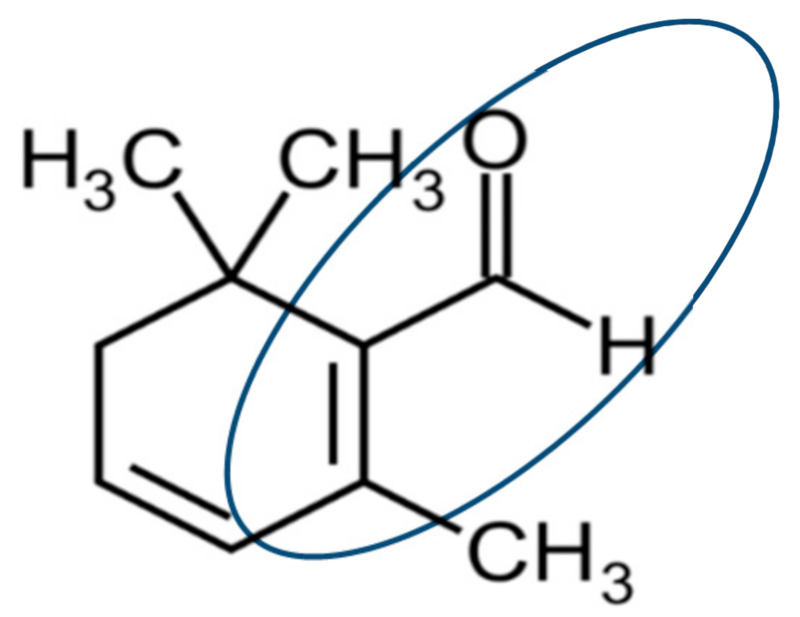
3.2. Headspace Gas Chromatography-Mass Spectrometry (HS-GC-MS)
3.3. Quantification and Visualization of H2O2 Using DAB Staining Method
3.4. Catalase Activity
- Blank 1: 1.0 mL substrate, 1.0 mL molybdate and 0.2 mL sample;
- Blank 2: 1.0 mL substrate, 1.0 mL molybdate and 0.2 mL buffer
- Blank 3 contained:1.0 mL buffer, 1.0 mL molybdate and 0.2 mL buffer.
3.5. Visualisation of Superoxide (O-2) and Determination of Peroxidase Activity in L. sativa Seedlings
- Δt: Time changed in min
- TV: Total volume
- UV: Volume used
- FW: Fresh weight of leaf tissue
3.6. In Situ Effect of Safranal on Growth and Physiological Characteristics of L. sativa and Some Common Weeds
3.6.1. Plant Material
3.6.2. Measurements of Fresh Weight, Chlorophyll Content, and Electrolyte Leakage
3.7. Data Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alagna, F.; Kallenbach, M.; Pompa, A.; De Marchis, F.; Rao, R.; Baldwin, I.T.; Bonaventure, G.; Baldoni, L. Olive fruits infested with olive fly larvae respond with an ethylene burst and the emission of specific volatiles. J. Integr. Plant Biol. 2016, 58, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, NY, USA, 1984; ISBN 9780080925394. [Google Scholar]
- Gnanavel, I. Eco-friendly weed control options for sustainable agriculture. Sci. Int. 2015, 3, 37–47. [Google Scholar] [CrossRef][Green Version]
- Oluwaseun, A.C.; Kola, O.J.; Micheal, B.O.; Mishra, P.; Sing, J.R. Isolation, structural elucidation and bioherbicidal activity of an eco-friendly bioactive 2-(hydroxymethyl) phenol, from Pseudomonas aeruginosa (C1501) and its ecotoxicological evaluation on soil. Environ. Technol. Innovation 2019, 13, 304–317. [Google Scholar]
- Vyvyan, J.R. Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 2002, 58, 1631–1636. [Google Scholar] [CrossRef]
- Romdhane, S.; Devers-Lamrani, M.; Beguet, J.; Bertrand, C.; Calvayrac, C.; Salvia, M.-V.; Jrad, A.B.; Dayan, F.E.; Spor, A.; Barthelmebs, L.; et al. Assessment of the ecotoxicological impact of natural and synthetic β-triketone herbicides on the diversity and activity of the soil bacterial community using omic approaches. Sci. Total Environ. 2019, 651, 241–249. [Google Scholar] [CrossRef]
- Amini, S.; Azizi, M.; Joharchi, M.R.; Shafei, M.; Moradinezhad, F.; Fujii, Y. Determination of allelopathic potential in some medicinal and wild plant species of Iran by dish pack method. Theor. Exp. Plant. Physiol. 2014, 26, 189–199. [Google Scholar] [CrossRef]
- Azizi, M.; Fuji, Y. Allelopathic effect of some medicinal plant substances on seed germination of Amaranthus retroflexus and Portulaca oleraceae. In Proceedings of the I International Symposium on Improving the Performance of Supply Chains in the Transitional Economies 699, Chiang Mai, Thailand, 19–23 July 2005; pp. 61–68. [Google Scholar]
- Azizi, M.; Farzad, S.; Jafarpour, B.; Rastegar, M.; Jahanbakhsh, V. In Inhibitory Effect of Some Medicinal Plants’ essential Oils on Postharvest Fungal Disease of Citrus Fruits. In Proceedings of the XXVII International Horticultural Congress-IHC2008: International Symposium on The Role of Postharvest Technology Globalisation of Horticulture, Seoul, Korea, 13 May 2008; pp. 279–286. [Google Scholar]
- Chizzola, R.; Saeidnejad, A.H.; Azizi, M.; Oroojalian, F.; Mardani, H. Bunium persicum: variability in essential oil and antioxidants activity of fruits from different Iranian wild populations. Genet. Resour. Crop. Ev. 2014, 61, 1621–1631. [Google Scholar] [CrossRef]
- Hamdi, A.; Majouli, K.; Vander Heyden, Y.; Flamini, G.; Marzouk, Z. Phytotoxic activities of essential oils and hydrosols of Haplophyllum tuberculatum. Ind. Crops Prod. 2017, 97, 440–447. [Google Scholar] [CrossRef]
- Bakand, S.; Winder, C.; Khalil, C.; Hayes, A. A novel in vitro exposure technique for toxicity testing of selected volatile organic compounds. J. Environ. Moni. 2006, 8, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Mardani, H.; Sekine, T.; Azizi, M.; Mishyna, M.; Fujii, Y. Identification of safranal as the main allelochemical from saffron (Crocus sativus). Nat. Prod. Commun. 2015, 10, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Mishyna, M.; Laman, N.; Prokhorov, V.; Maninang, J.S.; Fujii, Y. Identification of octanal as plant growth inhibitory volatile compound released from Heracleum sosnowskyi fruit. Nat. Prod. Commun 2015, 10, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Willard, P. Secrets of Saffron: The Vagabond Life of the World’s Most Seductive Spice; Beacon Press: Boston, MA, USA, 2002; p. 240. ISBN 13:978-0807050095. [Google Scholar]
- Hosseinzadeh, H.; Sadeghnia, H.R. Safranal, a constituent of Crocus sativus (saffron), attenuated cerebral ischemia induced oxidative damage in rat hippocampus. J. Pharm. Pharm. Sci. 2005, 8, 394–399. [Google Scholar]
- Rezaee, R.; Hosseinzadeh, H. Safranal: From an aromatic natural product to a rewarding pharmacological agent. Iran. J. Basi. Med. Sci. 2013, 16, 12. [Google Scholar]
- Samarghandian, S.; Shoshtari, M.E.; Sargolzaei, J.; Hossinimoghadam, H.; Farahzad, J.A. Anti-tumor activity of safranal against neuroblastoma cells. Pharmacogn. Mag. 2014, 10, 4192. [Google Scholar]
- Malaekeh-Nikouei, B.; Mousavi, S.H.; Shahsavand, S.; Mehri, S.; Nassirli, H.; Moallem, S.A. Assessment of cytotoxic properties of safranal and nanoliposomal safranal in various cancer cell lines. Phytother. Res. 2013, 27, 1868–1873. [Google Scholar] [CrossRef]
- Das, I.; Das, S.; Saha, T. Saffron suppresses oxidative stress in DMBA-induced skin carcinoma: A histopathological study. Acta Histochem. 2010, 112, 317–327. [Google Scholar] [CrossRef]
- Rahaiee, S.; Moini, S.; Hashemi, M.; Shojaosadati, S.A. Evaluation of antioxidant activities of bioactive compounds and various extracts obtained from saffron (Crocus sativus L.): A review. J. Food Sci. Technol. 2015, 52, 1881–1888. [Google Scholar] [CrossRef]
- Mashmoul, M.; Azlan, A.; Khaza’ai, H.; Yusof, B.N.M.; Noor, S.M. Saffron: A natural potent antioxidant as a promising anti-obesity drug. Antioxidants 2013, 2, 293–308. [Google Scholar] [CrossRef]
- Samanta, P.; Pal, S.; Mukherjee, A.K.; Ghosh, A.R. Biochemical effects of glyphosate based herbicide, Excel Mera 71 on enzyme activities of acetylcholinesterase (AChE), lipid peroxidation (LPO), catalase (CAT), glutathione-S-transferase (GST) and protein content on teleostean fishes. Ecotoxicol. Environ. Saf. 2014, 107, 120–125. [Google Scholar] [CrossRef]
- Salbego, J.; Pretto, A.; Gioda, C.R.; de Menezes, C.C.; Lazzari, R.; Neto, J.R.; Baldisserotto, B.; Loro, V.L. Herbicide formulation with glyphosate affects growth, acetylcholinesterase activity, and metabolic and hematological parameters in piava (Leporinus obtusidens). Arch. Environ. Contam. Toxicol. 2010, 58, 740–745. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, Y.; Jia, L.X.; Lin, J.L.; Liu, Y.; Pan, B.; Lin, Y. Biological responses of wheat (Triticum aestivum) plants to the herbicide simetryne in soils. Ecotoxicol. Environ. Saf. 2016, 127, 87–94. [Google Scholar] [CrossRef]
- Cruz-Ortega, R.; Lara-Núñez, A.; Anaya, A.L. Allelochemical stress can trigger oxidative damage in receptor plants: Mode of action of phytotoxicity. Plant Signaling Behav. 2007, 2, 269–270. [Google Scholar] [CrossRef]
- Liu, M.; Amini, A.; Ahmad, Z. Safranal and its analogs inhibit Escherichia coli ATP synthase and cell growth. Int. J. Biol. Macromol. 2016, 95, 145–152. [Google Scholar] [CrossRef]
- Amini, A.; Liu, M.; Ahmad, Z. Understanding the link between antimicrobial properties of dietary olive phenolics and bacterial ATP synthase. Int. J. Biol. Macromol. 2017, 101, 153–164. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, H. Prometryne-induced oxidative stress and impact on antioxidant enzymes in wheat. Ecotoxicol. Environ. Saf. 2009, 72, 1687–1693. [Google Scholar] [CrossRef]
- Mardani, H.; Bayat, H.; Saeidnejad, A.H.; Rezaie, E.E. Assessment of salicylic acid impacts on seedling characteristic of cucumber (Cucumis sativus L.) under water stress. Not. Sci. Biol. 2012, 4, 112. [Google Scholar] [CrossRef]
- Smirnov, N.N. Nutrition. In Physiology of the Cladocera; Smirnov, N.N., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 33–74, Chapter 4. [Google Scholar]
- Dölle, C.; Bindoff, L.A.; Tzoulis, C. 3, 3′-Diaminobenzidine staining interferes with PCR-based DNA analysis. Sci. Rep. 2018, 8, 1272. [Google Scholar]
- Lee, B.-h.; Lee, H.; Xiong, L.; Zhu, J.-K. A mitochondrial complex I defect impairs cold-regulated nuclear gene expression. Plant Cell 2002, 14, 1235–1251. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef]
- Ramel, F.; Sulmon, C.; Bogard, M.; Couée, I.; Gouesbet, G. Differential patterns of reactive oxygen species and antioxidative mechanisms during atrazine injury and sucrose-induced tolerance in Arabidopsis thaliana plantlets. BMC Plant Biol. 2009, 9, 28. [Google Scholar] [CrossRef]
- Sunmonu, T.; Van Staden, J. Phytotoxicity evaluation of six fast-growing tree species in South Africa. S. Afr. J. Bot. 2014, 90, 101–106. [Google Scholar] [CrossRef][Green Version]
- Farahmand, S.K.; Samini, F.; Samini, M.; Samarghandian, S. Safranal ameliorates antioxidant enzymes and suppresses lipid peroxidation and nitric oxide formation in aged male rat liver. Biogerontology 2013, 14, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Naghshineh, A.; Dadras, A.; Ghalandari, B.; Riazi, G.H.; Modaresi, S.M.S.; Afrasiabi, A.; Aslani, M.K. Safranal as a novel anti-tubulin binding agent with potential use in cancer therapy: An in vitro study. Chem. Biol. Interact 2015, 238, 151–160. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Choi, M.-Y.; Paik, C.-H.; Kalaivani, K. The toxicity and physiological effect of goniothalamin, a styryl-pyrone, on the generalist herbivore, Spodoptera exigua Hübner. Chemosphere 2008, 72, 1393–1400. [Google Scholar] [CrossRef]
- Pudełko, K.; Majchrzak, L.; Narożna, D. Allelopathic effect of fibre hemp (Cannabis sativa L.) on monocot and dicot plant species. Ind. Crops Prod. 2014, 56, 191–199. [Google Scholar] [CrossRef]
- Zhang, E.-P.; Zhang, S.-H.; Li, W.-B.Z.L.-L.; Li, T.-L. Effects of exogenic benzoic acid and cinnamic acid on the root oxidative damage of tomato seedlings. J. Hortic. For. 2010, 2, 22–29. [Google Scholar]
- Khan, A.L.; Hamayun, M.; Hussain, J.; Khan, H.; Gilani, S.A.; Kikuchi, A.; Watanabe, K.N.; Jung, E.H.; Lee, I. Assessment of allelopathic potential of selected medicinal plants of Pakistan. Afr. J. Biotechnol. 2009, 8. [Google Scholar]
- Frank, D.; O’Riordan, P.; Varelis, P.; Zabaras, D.; Watkins, P.; Ceccato, C.; Wijesundera, C.; Ferguson, A.; Hewett, E.; Gunson, F. Deconstruction and recreation of ‘Hayward’ volatile flavour using a trained sensory panel, olfactometry and a kiwifruit model matrix. Acta Hortic. 2007, 753, 107–118. [Google Scholar] [CrossRef]
- Burdon, J.; McLeod, D.; Lallu, N.; Gamble, J.; Petley, M.; Gunson, A. Consumer evaluation of “Hayward” kiwifruit of different at-harvest dry matter contents. Postharvest Biol. Technol. 2004, 34, 245–255. [Google Scholar] [CrossRef]
- Lee, J.; Joshi, N.; Pasini, R.; Dobson, R.C.; Allison, J.; Leustek, T. Inhibition of Arabidopsis growth by the allelopathic compound azetidine-2-carboxylate is due to the low amino acid specificity of cytosolic prolyl-tRNA synthetase. Plant J. 2016, 88, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Ma, N.N.; Wang, J.R.; Yang, D.Y.; Zhao, S.J.; Meng, Q.W. Overexpression of tomato enhancer of SOS3-1 (LeENH1) in tobacco enhanced salinity tolerance by excluding Na+ from the cytosol. Plant Physiol. Biochem. 2013, 70, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Hadwan, M.H.; Abed, H.N. Data supporting the spectrophotometric method for the estimation of catalase activity. Data Brief 2016, 6, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Cai, G.; Jiang, S.; Sun, L.; Li, D. Response of tobacco to the Pseudomonas syringae pv. Tomato DC3000 is mainly dependent on salicylic acid signaling pathway. FEMS Microbiol. Lett. 2013, 344, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.S.; Wang, S.J.; Yang, Z.M. Biological detection and analysis of mercury toxicity to alfalfa (Medicago sativa) plants. Chemosphere 2008, 70, 1500–1509. [Google Scholar] [CrossRef]
- Pilloff, R.K.; Devadas, S.K.; Enyedi, A.; Raina, R. The Arabidopsis gain-of-function mutant dll1 spontaneously develops lesions mimicking cell death associated with disease. Plant J. 2002, 30, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Tan, K.S.; Zhang, X.; Sun, A.Y.; Sun, G.Y.; Lee, J.C.-M. Hydrogen peroxide alters membrane and cytoskeleton properties and increases intercellular connections in astrocytes. J. Cell Sci. 2005, 118, 3695–3703. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organoleptic Properties | Safranal (CAS No 116-26-7) |
|---|---|
| Appearance | Pale yellow to Yellow liquid |
| Molecular Formula | C10H14O |
| Molecular Weight | 150.22 |
| Specific Gravity | 0.950–0.970 at 25 °C |
| Vapor Pressure | 0.134 mm/Hg at 25.0 °C. (est) |
| Water solubility | Insoluble |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mardani, H.; Maninang, J.; Appiah, K.S.; Oikawa, Y.; Azizi, M.; Fujii, Y. Evaluation of Biological Response of Lettuce (Lactuca sativa L.) and Weeds to Safranal Allelochemical of Saffron (Crocus sativus) by Using Static Exposure Method. Molecules 2019, 24, 1788. https://doi.org/10.3390/molecules24091788
Mardani H, Maninang J, Appiah KS, Oikawa Y, Azizi M, Fujii Y. Evaluation of Biological Response of Lettuce (Lactuca sativa L.) and Weeds to Safranal Allelochemical of Saffron (Crocus sativus) by Using Static Exposure Method. Molecules. 2019; 24(9):1788. https://doi.org/10.3390/molecules24091788
Chicago/Turabian StyleMardani, Hossein, John Maninang, Kwame Sarpong Appiah, Yosei Oikawa, Majid Azizi, and Yoshiharu Fujii. 2019. "Evaluation of Biological Response of Lettuce (Lactuca sativa L.) and Weeds to Safranal Allelochemical of Saffron (Crocus sativus) by Using Static Exposure Method" Molecules 24, no. 9: 1788. https://doi.org/10.3390/molecules24091788
APA StyleMardani, H., Maninang, J., Appiah, K. S., Oikawa, Y., Azizi, M., & Fujii, Y. (2019). Evaluation of Biological Response of Lettuce (Lactuca sativa L.) and Weeds to Safranal Allelochemical of Saffron (Crocus sativus) by Using Static Exposure Method. Molecules, 24(9), 1788. https://doi.org/10.3390/molecules24091788