In Silico Investigation of the Anti-Tumor Mechanisms of Epigallocatechin-3-Gallate


Abstract
:
1. Introduction
2. Results
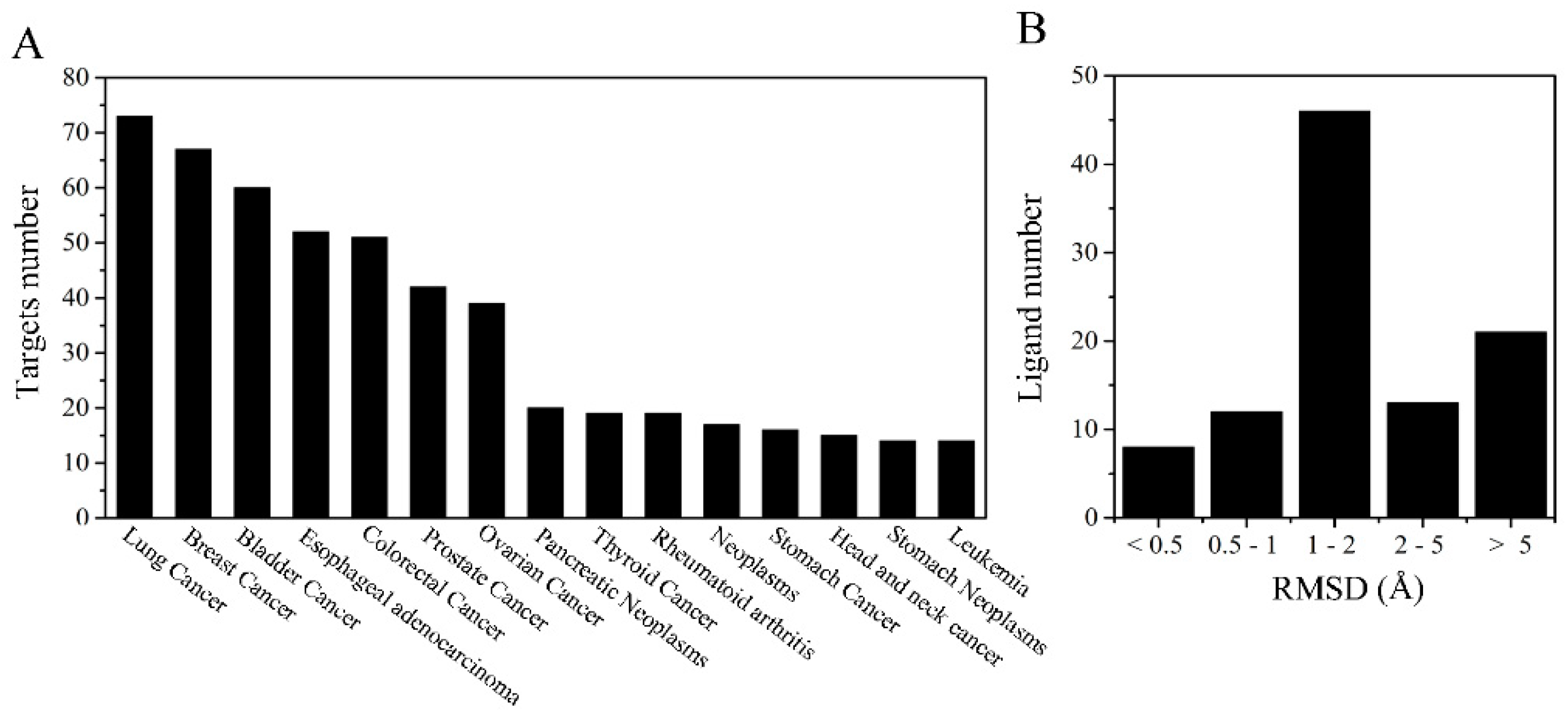
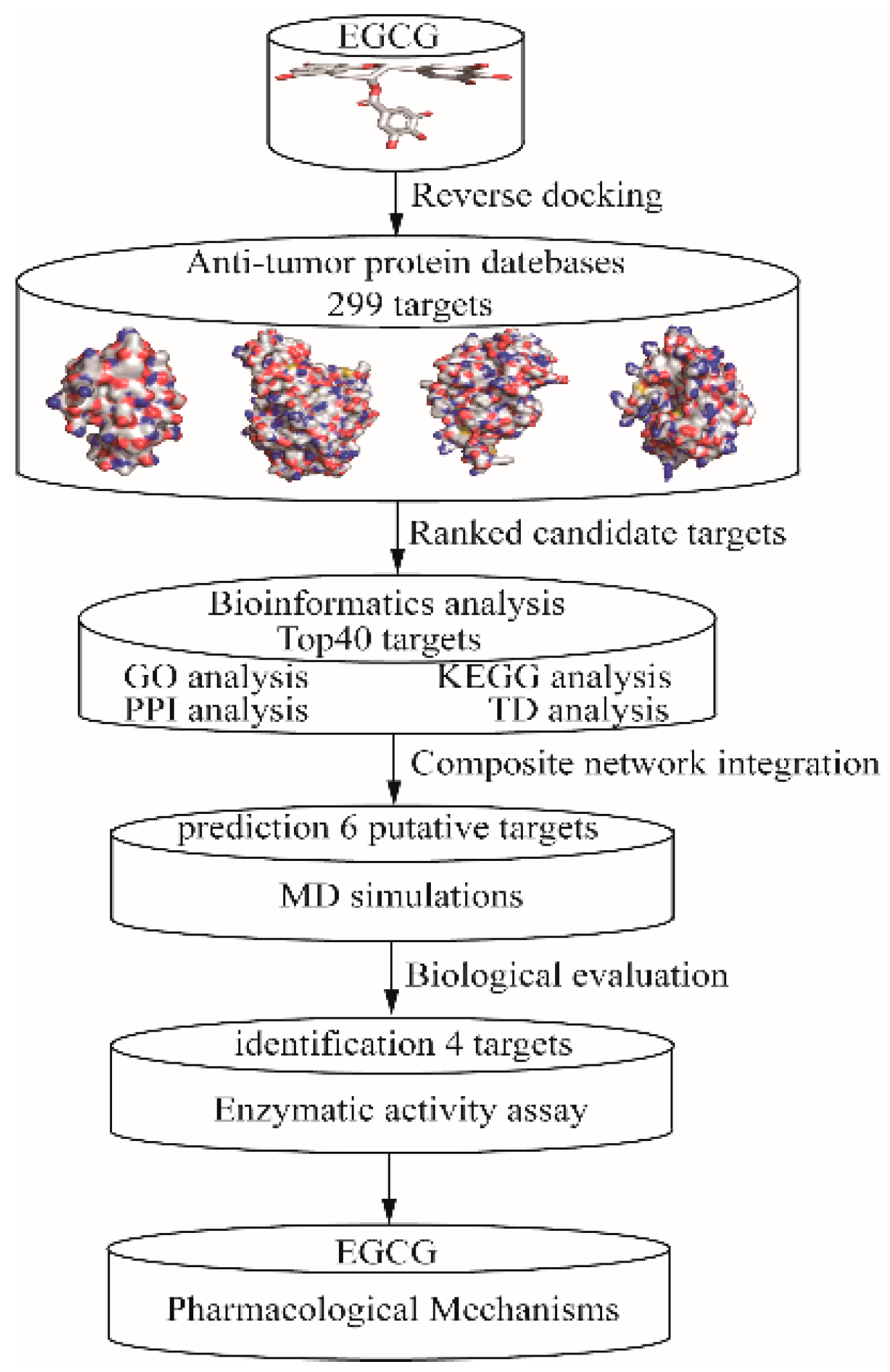
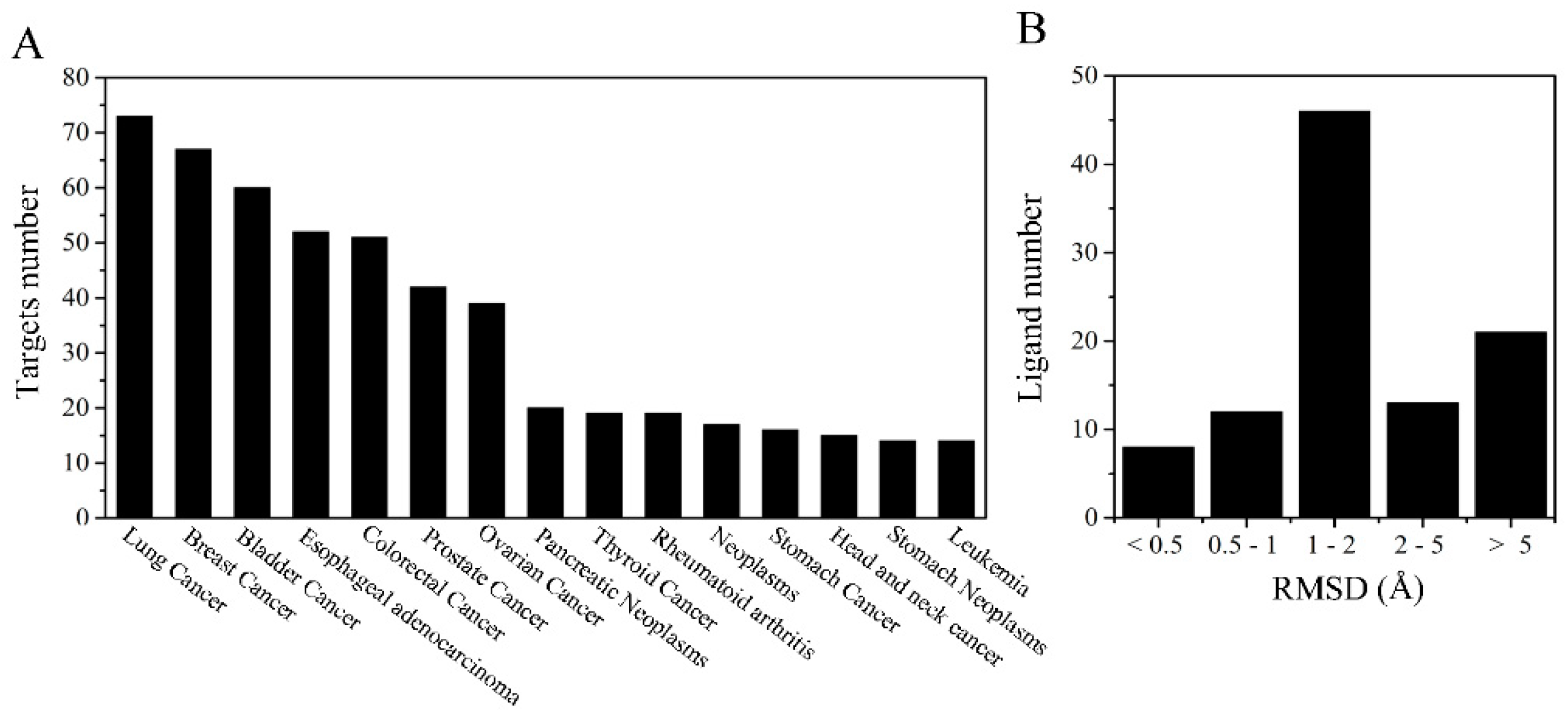
2.1. Target Proteins Database and Reverse Virtual Screening
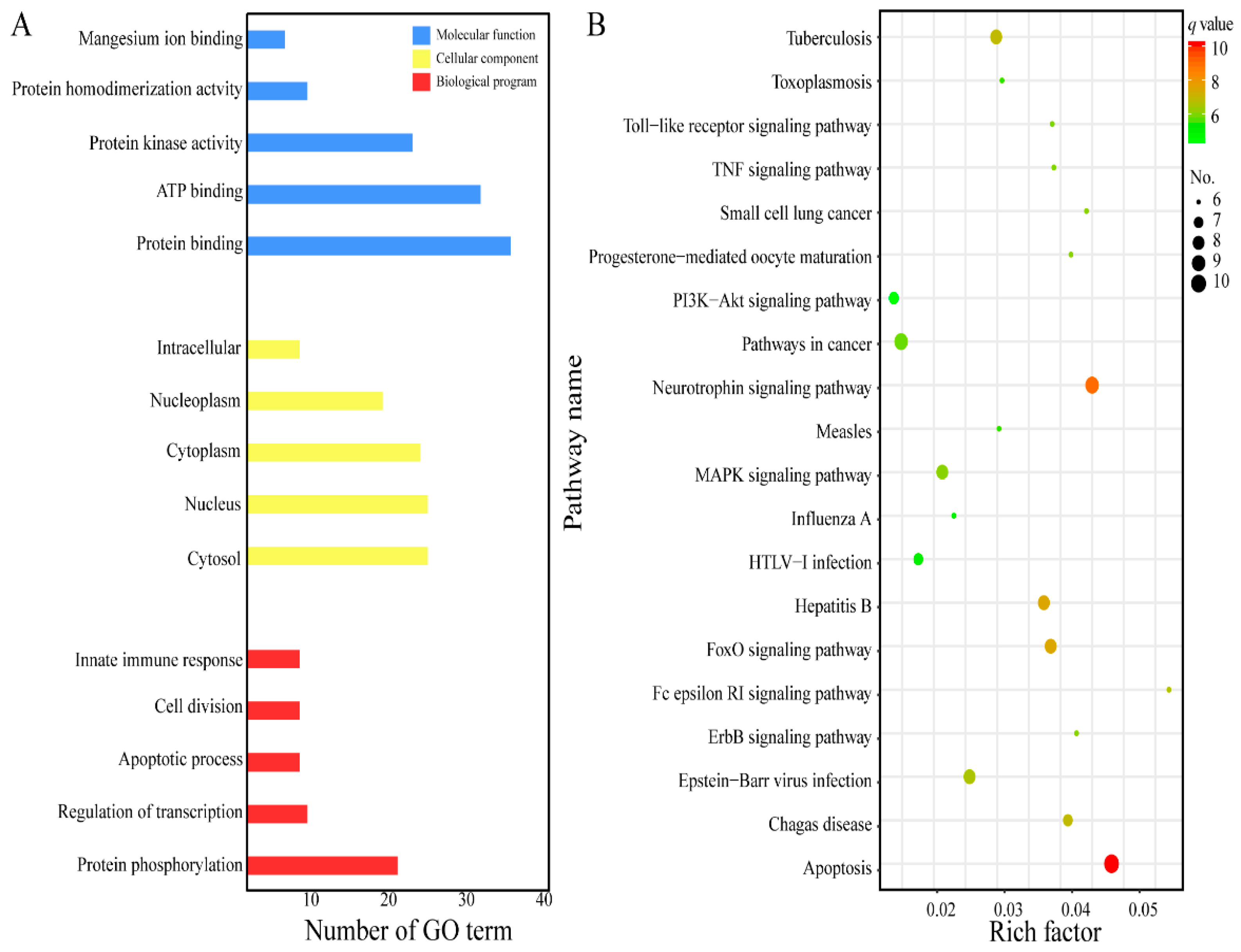
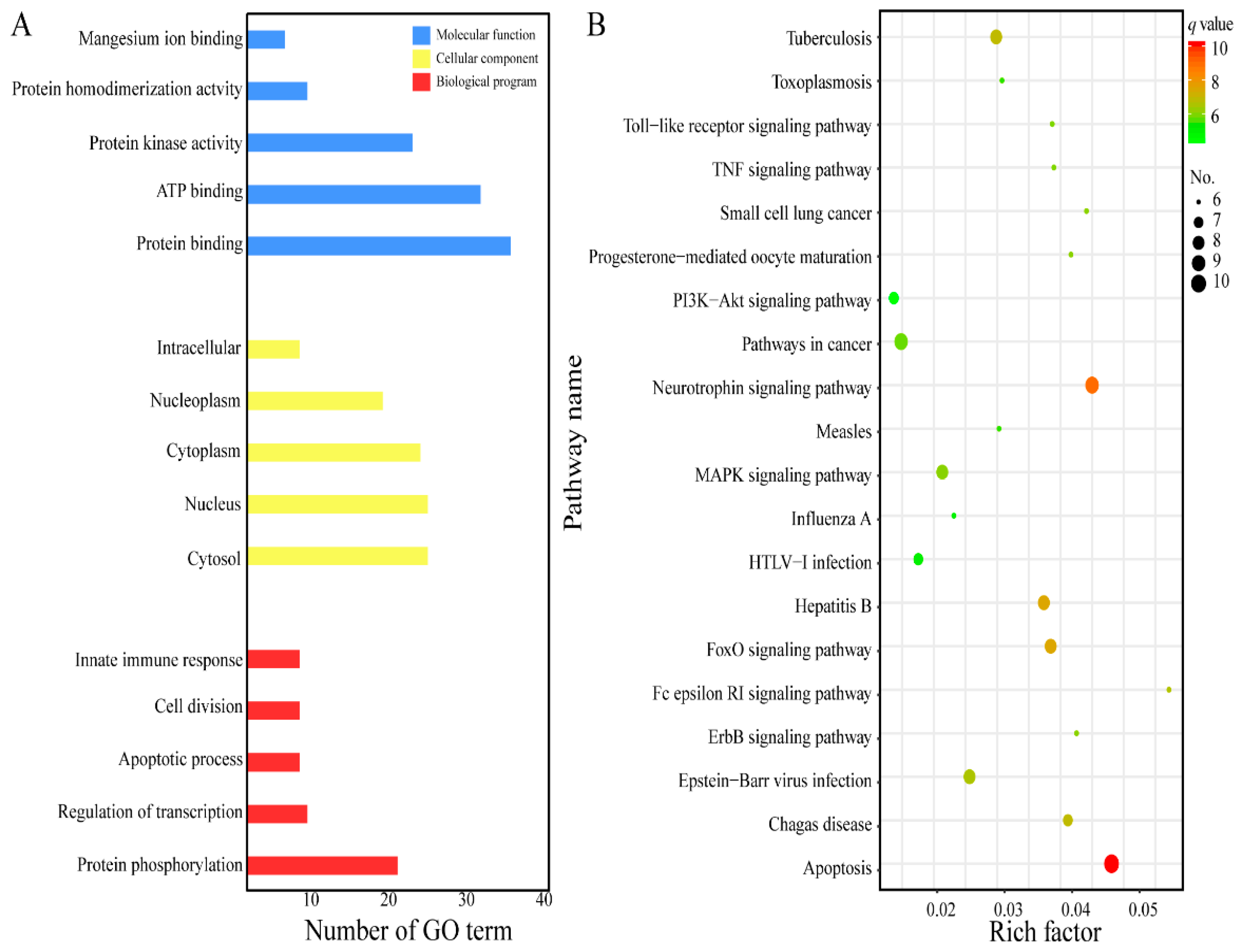
2.2. GO Analysis and KEGG Pathway Enrichment
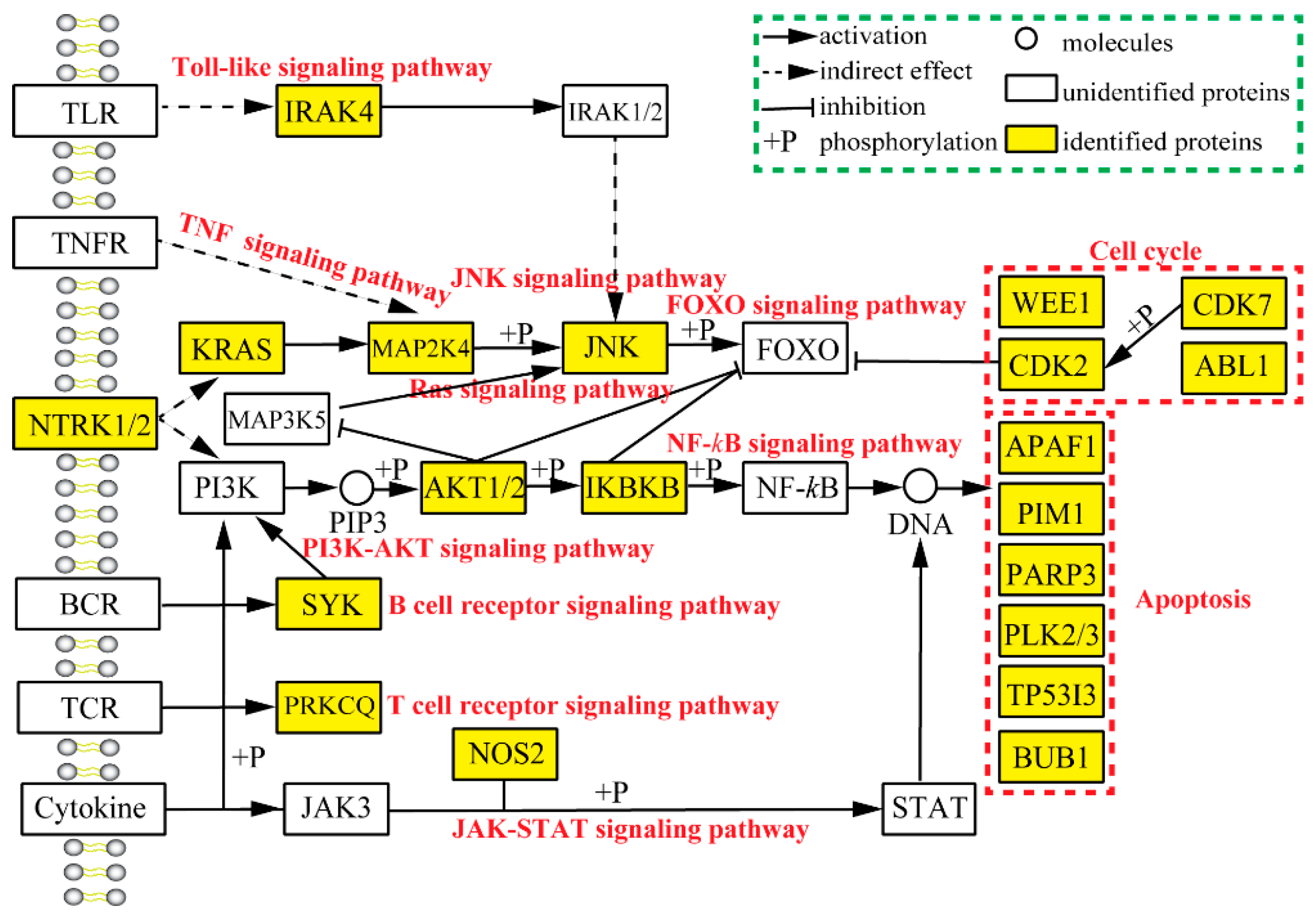
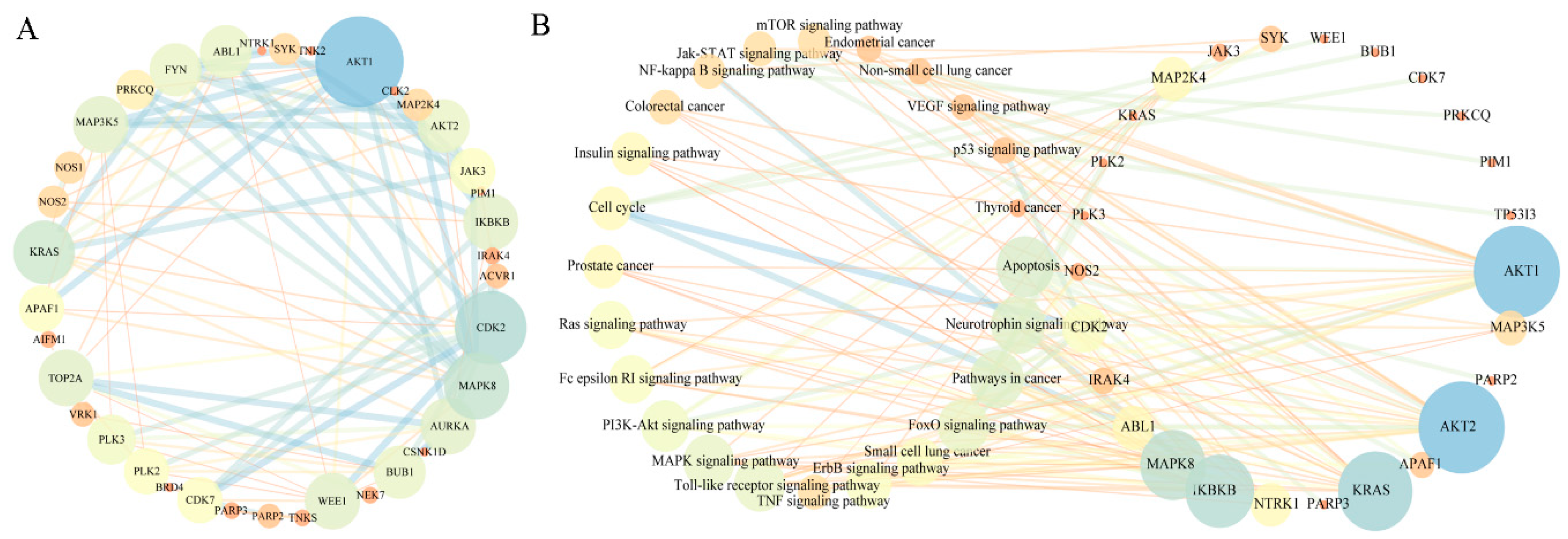
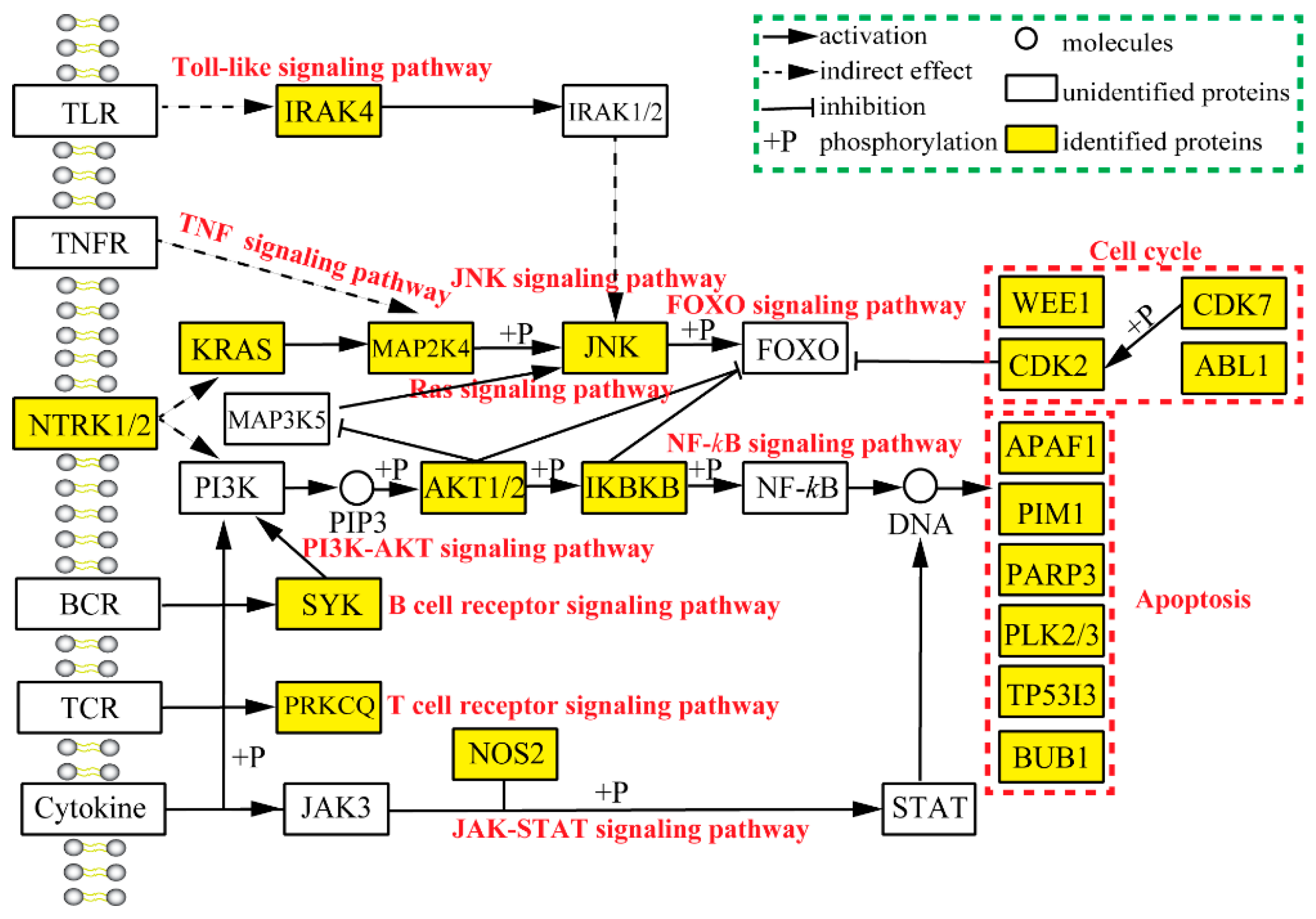
2.3. Analysis of Pharmacological Network
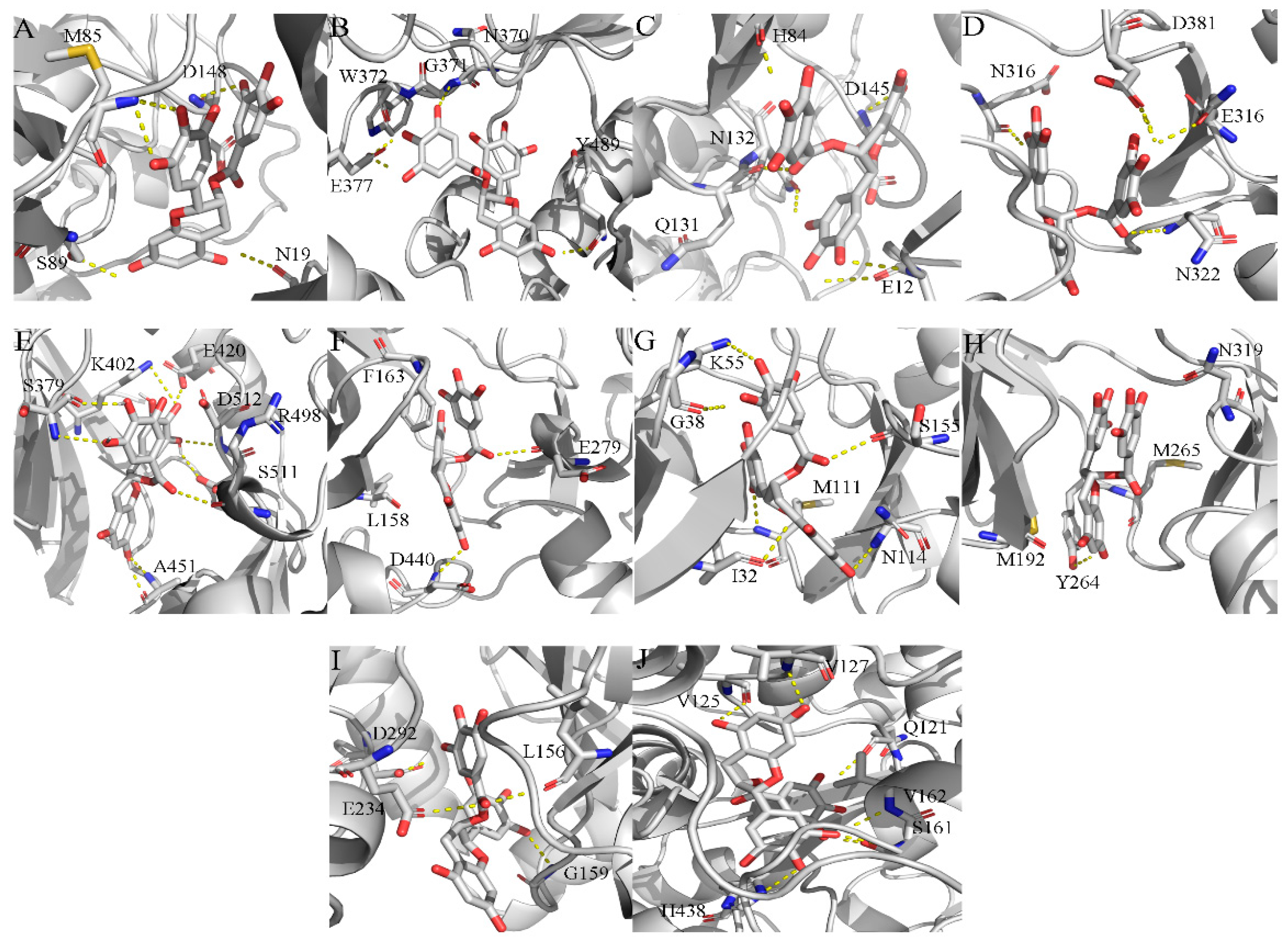
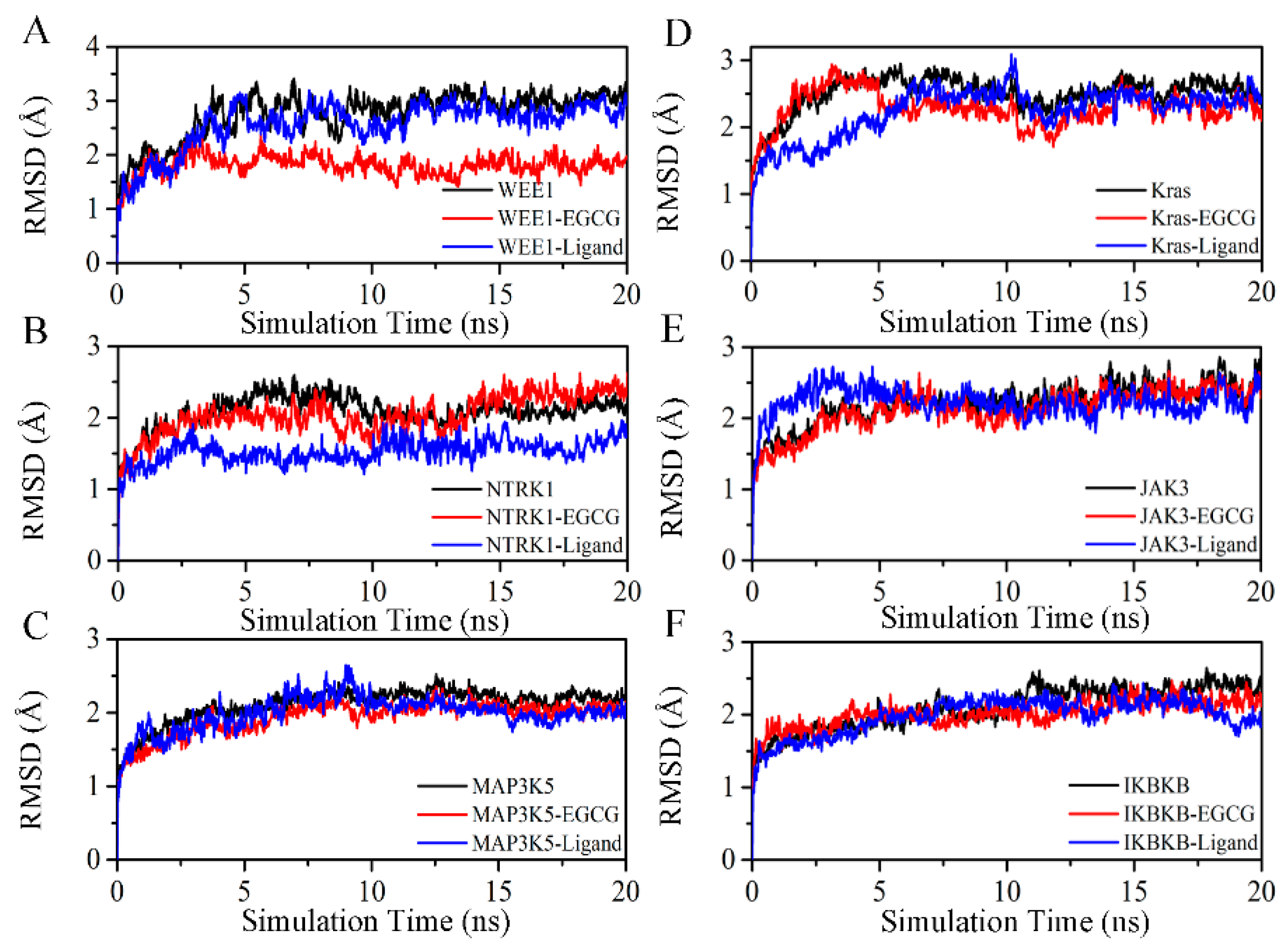
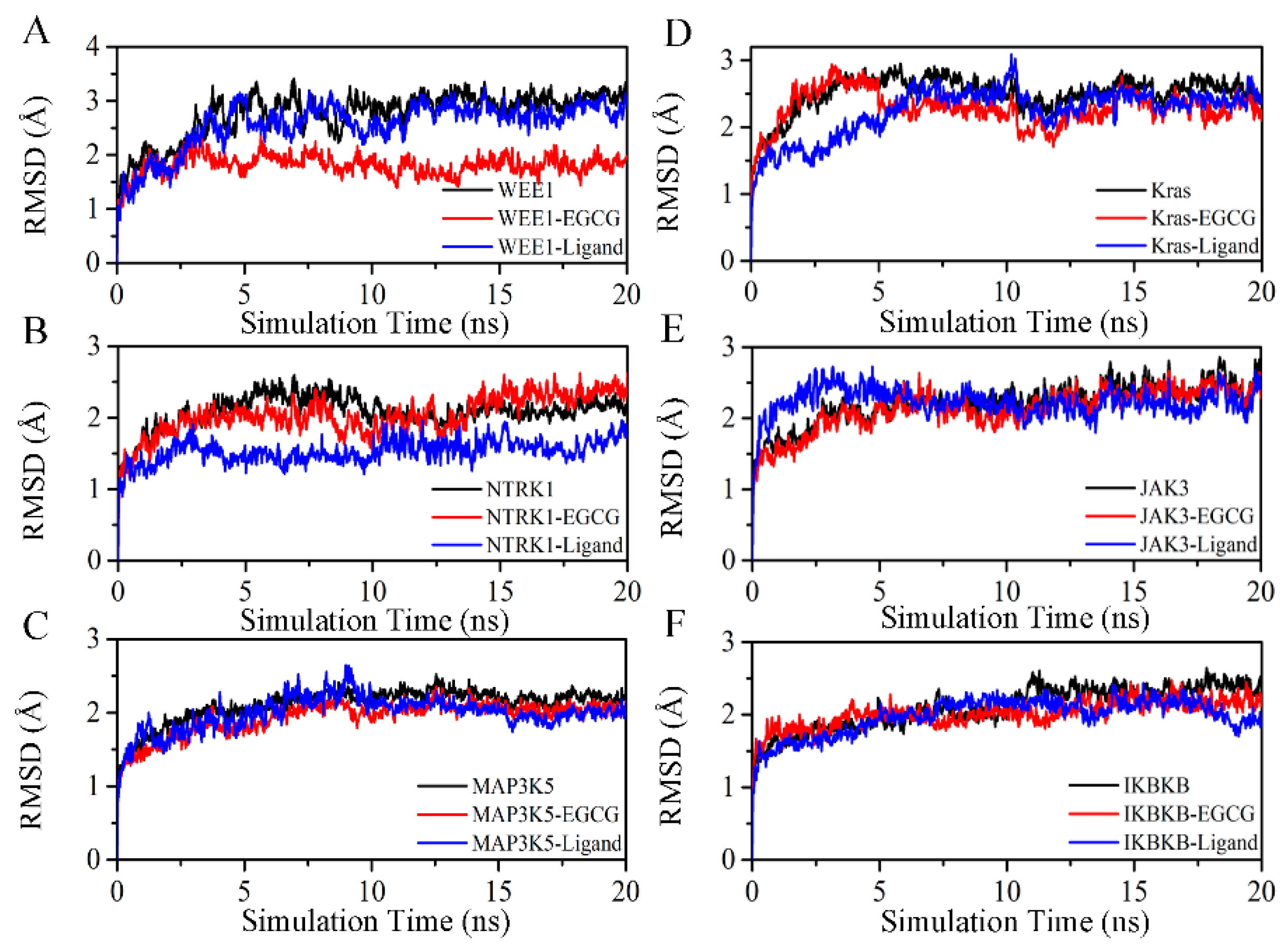
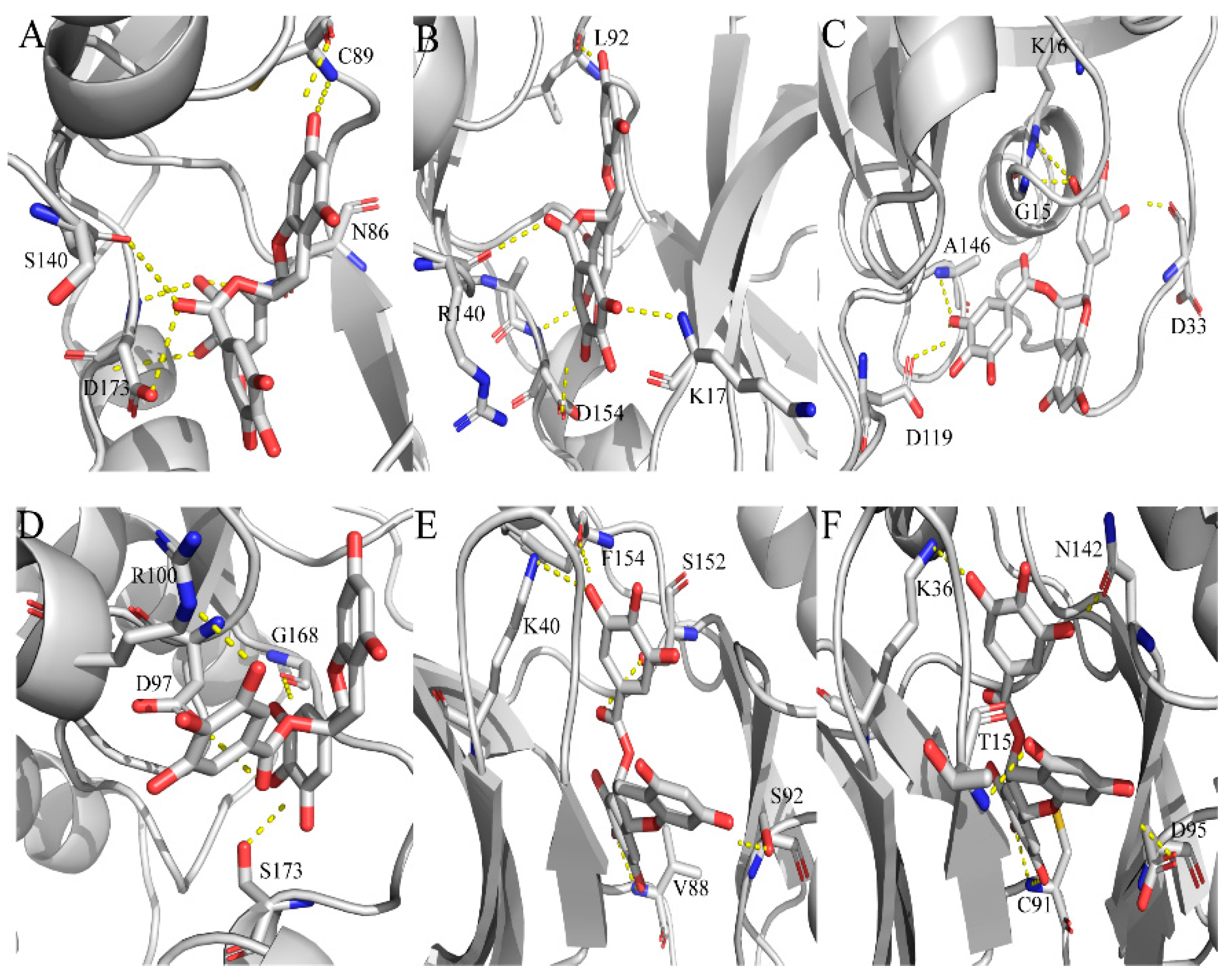
2.4. Exploration and Identification of the Potential Target Proteins
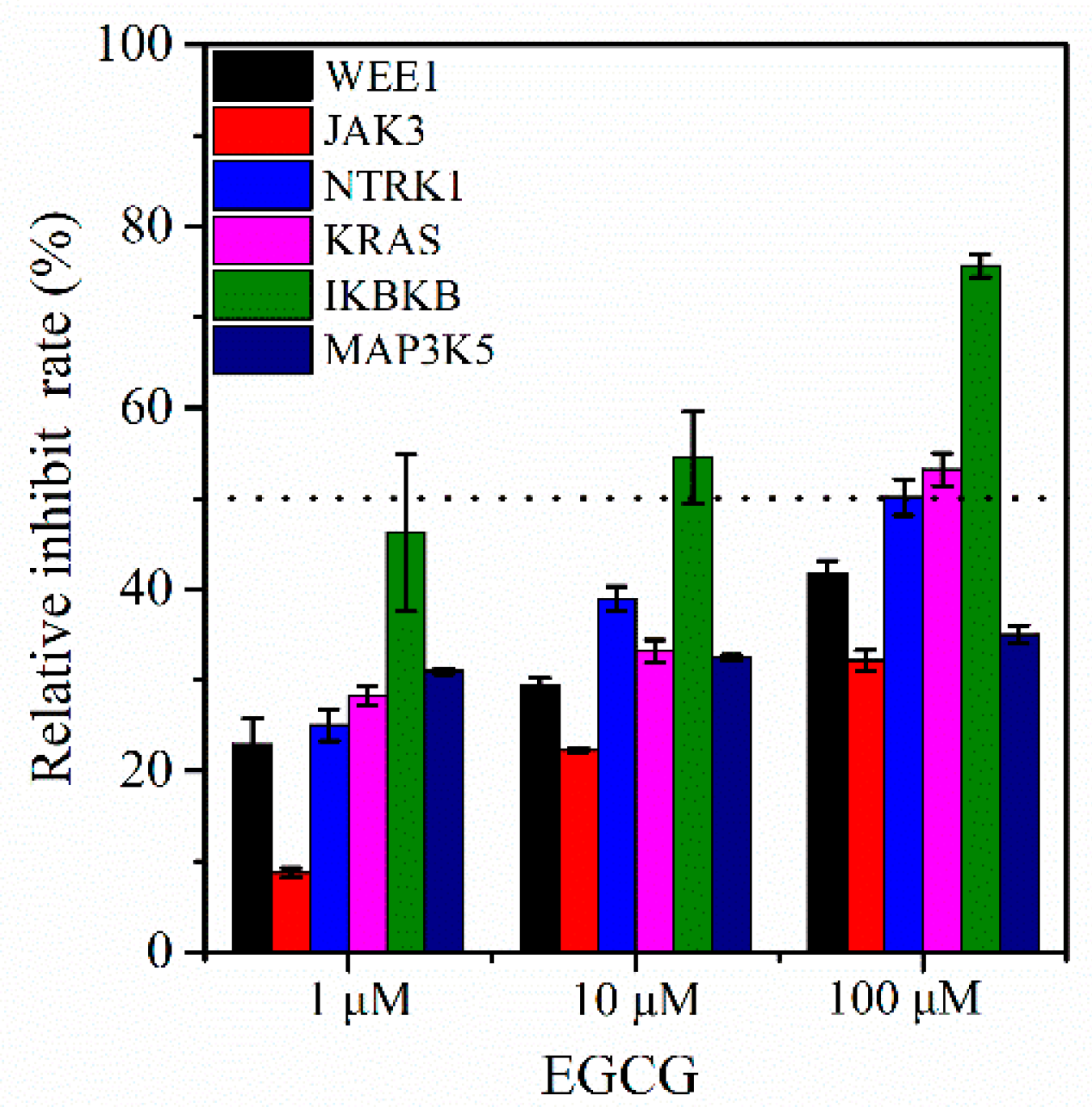
2.5. Enzymatic Activity Assay In Vitro
3. Discussion
4. Materials and Methods
4.1. Construction of Anti-tumor Target Proteins Database
4.2. Reverse Virtual Screening
4.3. GO Analysis and KEGG Pathway Enrichment
4.4. Pharmacological Network Analysis
4.5. Molecular Dynamic Simulation
4.6. Enzymatic Activity Assay In Vitro
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviation
| EC | Epicatechin |
| GC | Gallocatechin |
| EGC | Epigallocatechin |
| CG | Catechin-3-gallate |
| ECG | Epicatechin-3-gallate |
| GCG | Gallocatechin-3-gallate |
| EGCG | Epigallocatechin-3-gallate |
| Pin1 | Peptidyl-prolyl cis/trans isomerase NIMA-interacting 1 |
| MD | Molecular Dynamic |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| DAVID | The Database for Annotation, Visualization and Integrated Discovery |
| PPI | Protein–protein Interaction |
| TP | Target–pathway |
| NVT | Constant number of particles, volume and temperature |
| NPT | Constant number of particles, pressure and temperature |
| PME | Particle Mesh Ewald |
| LINCS | Linear Constraint Solver |
| RMSD | Root Mean Square Deviation |
| GAD | Genetic Association Database |
| FYN | Proto-oncogene tyrosine-protein kinase FYN |
| NOS2 | Nitric Oxide Synthase 2 |
| CDK2 | Cyclin-Dependent Kinase 2 |
| CDK7 | Cyclin-Dependent Kinase 7 |
| ABL1 | Abelson Murine Leukemia Viral Oncogene Homolog 1 |
| SYK | Spleen tyrosine kinase |
| AKT2 | Serine-threonine protein kinase AKT2 |
| MAPK8 | Mitogen-activated protein kinase 8 |
| AKT1 | Serine-threonine protein kinase AKT1 |
| KRAS | Kirsten Rat Sarcoma Viral Oncogene |
| IRAK4 | Interleukin-1 Receptor-Associated Kinase 4 |
| WEE1 | Mitosis inhibitor protein kinase WEE1 |
| JAK3 | Tyrosine-protein kinase JAK3 |
| APAF1 | Apoptotic Protease Activating Factor 1 |
| NTRK1 | High affinity nerve growth factor receptor 1 |
| NTRK2 | High affinity nerve growth factor receptor 2 |
| MAP3K5 | Mitogen-Activated Protein Kinase Kinase Kinase 5 |
| IKBKB | Inhibitor of nuclear factor kappa-B kinase subunit beta |
| TP53I3 | Quinone oxidoreductase PIG3 |
| PIM1 | Serine/threonine-protein kinase PIM1 |
| PARP3 | Poly ADP-ribose polymerase 3 |
| PLK2 | Serine/threonine-protein kinase PLK2 |
| PLK3 | Serine/threonine-protein kinase PLK3 |
| BUB1 | Mitotic checkpoint serine/threonine-protein kinase BUB1 |
| AKT1 | RAC-alpha serine/threonine-protein kinase 1 |
| AKT2 | RAC-alpha serine/threonine-protein kinase 2 |
| PRKCQ | Protein kinase C theta type |
| JAK | Tyrosine-protein kinase JAK |
| STAT | Signal transducer and activator of transcription |
| NF-kB | Nuclear factor NF-kappa-B |
| FOXO | Forkhead Box O |
| PI3K | Phosphoinositide 3-kinase |
| iNOS | Inducible nitric oxide synthase |
| ICAM-1 | Intercellular cell adhesion molecule-1 |
| TNF | Tumor Necrosis Factor |
| TNFR | Tumor necrosis factor receptor |
| TLR | Toll-like receptors |
| BCR | B cell receptor |
| TCR | T cell receptor |
| NFKB1 | Nuclear factor NF-kappa-B |
| MAPK | Mitogen-activated protein kinase |
References
- Park, S.Y.; Lee, Y.-K.; Kim, Y.-M.; Park, O.J.; Shin, J.-I. Control of AMP-activated protein kinase, Akt and mTOR in EGCG-treated HT-29 colon cancer cells. Food Sci. Biotechnol. 2013, 22, 147–151. [Google Scholar] [CrossRef]
- Wang, Y.-Q.; Lu, J.-L.; Liang, Y.-R.; Li, Q.-S. Suppressive effects of egcg on cervical cancer. Molecules 2018, 23, 2334. [Google Scholar] [CrossRef]
- Benvenuto, M.; Fantini, M.; Masuelli, L.; De Smaele, E.; Zazzeroni, F.; Tresoldi, I.; Calabrese, G.; Galvano, F.; Modesti, A.; Bei, R. Inhibition of ErbB receptors, Hedgehog and NF-kappaB signaling by polyphenols in cancer. Front. Biosci. 2013, 18, 1290–1310. [Google Scholar]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef]
- Gao, Z.; Han, Y.; Hu, Y.; Wu, X.; Wang, Y.; Zhang, X.; Fu, J.; Zou, X.; Zhang, J.; Chen, X. Targeting HO-1 by epigallocatechin-3-gallate reduces contrast-induced renal injury via anti-oxidative stress and anti-inflammation pathways. PLoS ONE 2016, 11, e0149032. [Google Scholar] [CrossRef]
- Riegsecker, S.; Wiczynski, D.; Kaplan, M.J.; Ahmed, S. Potential benefits of green tea polyphenol EGCG in the prevention and treatment of vascular inflammation in rheumatoid arthritis. Life Sci. 2013, 93, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, E.; Suzuki, H.; Menegazzi, M. Antiinflammatory action of EGCG, the main component of green tea, through STAT-1 inhibition. Ann. N. Y. Acad. Sci. 2002, 973, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Deana, R.; Turetta, L.; Donella-Deana, A.; Donà, M.; Brunati, A.M.; De Michiel, L.; Garbisa, S. Green tea epigallocatechin-3-gallate inhibits platelet signalling pathways triggered by both proteolytic and non-proteolytic agonists. Thromb. Haemost. 2003, 89, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Rashidi, B.; Malekzadeh, M.; Goodarzi, M.; Masoudifar, A.; Mirzaei, H. Green tea and its anti-angiogenesis effects. Biomed. Pharmacother. 2017, 89, 949–956. [Google Scholar] [CrossRef]
- García-Vilas, J.A.; Quesada, A.R.; Medina, M.Á. Screening of synergistic interactions of epigallocatechin-3-gallate with antiangiogenic and antitumor compounds. Synergy 2016, 3, 5–13. [Google Scholar] [CrossRef]
- Diniz, C.; Suliburska, J.; Ferreira, I.M. New insights into the antiangiogenic and proangiogenic properties of dietary polyphenols. Mol. Nutr. Food Res. 2017, 61, 1600912. [Google Scholar] [CrossRef]
- Zheng, R.; Chen, T.-S.; Lu, T. A comparative reverse docking strategy to identify potential antineoplastic targets of tea functional components and binding mode. Int. J. Mol. Sci. 2011, 12, 5200–5212. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Saleem, M.; Ahmad, N.; Mukhtar, H. Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res. 2006, 66, 2500–2505. [Google Scholar] [CrossRef]
- Saeki, K.; Hayakawa, S.; Nakano, S.; Ito, S.; Oishi, Y.; Suzuki, Y.; Isemura, M. In vitro and in silico studies of the molecular interactions of epigallocatechin-3-o-gallate (egcg) with proteins that explain the health benefits of green tea. Molecules 2018, 23, 1295. [Google Scholar] [CrossRef]
- Gupta, S.; Hastak, K.; Afaq, F.; Ahmad, N.; Mukhtar, H. Essential role of caspases in epigallocatechin-3-gallate-mediated inhibition of nuclear factor kappaB and induction of apoptosis. Oncogene 2004, 23, 2507. [Google Scholar] [CrossRef]
- Shankar, S.; Suthakar, G.; Srivastava, R.K. Epigallocatechin-3-gallate inhibits cell cycle and induces apoptosis in pancreatic cancer. Front. Biosci. 2007, 12, 5039–5051. [Google Scholar] [CrossRef]
- Masuda, M.; Suzui, M.; Weinstein, I.B. Effects of epigallocatechin-3-gallate on growth, epidermal growth factor receptor signaling pathways, gene expression and chemosensitivity in human head and neck squamous cell carcinoma cell lines. Clin. Cancer Res. 2001, 7, 4220–4229. [Google Scholar]
- Bartholome, A.; Kampkötter, A.; Tanner, S.; Sies, H.; Klotz, L.-O. Epigallocatechin gallate-induced modulation of FoxO signaling in mammalian cells and C. elegans: FoxO stimulation is masked via PI3K/Akt activation by hydrogen peroxide formed in cell culture. Arch. Biochem. Biophys. 2010, 501, 58–64. [Google Scholar] [CrossRef]
- Byun, E.H.; Fujimura, Y.; Yamada, K.; Tachibana, H. TLR4 signaling inhibitory pathway induced by green tea polyphenol epigallocatechin-3-gallate through 67-kDa laminin receptor. J. Immunol. 2010, 185, 33–45. [Google Scholar] [CrossRef]
- Byun, E.-H.; Omura, T.; Yamada, K.; Tachibana, H. Green tea polyphenol epigallocatechin-3-gallate inhibits TLR2 signaling induced by peptidoglycan through the polyphenol sensing molecule 67-kDa laminin receptor. FEBS Lett. 2011, 585, 814–820. [Google Scholar] [CrossRef]
- Urusova, D.V.; Shim, J.-H.; Kim, D.J.; Jung, S.K.; Zykova, T.A.; Carper, A.; Bode, A.M.; Dong, Z. Epigallocatechin-gallate suppresses tumorigenesis by directly targeting Pin1. Cancer Prev. Res. 2011, 4, 1366–1377. [Google Scholar] [CrossRef]
- Xi, L.; Wang, Y.; He, Q.; Zhang, Q.; Du, L. Interaction between Pin1 and its natural product inhibitor epigallocatechin-3-gallate by spectroscopy and molecular dynamics simulations. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2016, 169, 134–143. [Google Scholar] [CrossRef]
- Nakano, S.; Megro, S.-I.; Hase, T.; Suzuki, T.; Isemura, M.; Nakamura, Y.; Ito, S. Computational molecular docking and X-ray crystallographic studies of catechins in new drug design strategies. Molecules 2018, 23, 2020. [Google Scholar] [CrossRef]
- Negri, A.; Naponelli, V.; Rizzi, F.; Bettuzzi, S. Molecular Targets of Epigallocatechin—Gallate (EGCG): A Special Focus on Signal Transduction and Cancer. Nutrients 2018, 10, 1936. [Google Scholar] [CrossRef]
- Li, H.; Sun, X.; Yu, F.; Xu, L.; Miu, J.; Xiao, P. In silico Investigation of the Pharmacological Mechanisms of Beneficial Effects of Ginkgo biloba L. on Alzheimer’s Disease. Nutrients 2018, 10, 589. [Google Scholar] [CrossRef]
- Chen, F.; Wang, Z.; Wang, C.; Xu, Q.; Liang, J.; Xu, X.; Yang, J.; Wang, C.; Jiang, T.; Yu, R. Application of reverse docking for target prediction of marine compounds with anti-tumor activity. J. Mol. Graph. Model. 2017, 77, 372–377. [Google Scholar] [CrossRef]
- Kharkar, P.S.; Warrier, S.; Gaud, R.S. Reverse docking: A powerful tool for drug repositioning and drug rescue. Future Med. Chem. 2014, 6, 333–342. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, G.; Zhou, Y.; Lin, C.; Chen, S.; Lin, Y.; Mai, S.; Huang, Z. Reverse screening methods to search for the protein targets of chemopreventive compounds. Front. Chem. 2018, 6, 1–28. [Google Scholar] [CrossRef]
- Ma, C.; Tang, K.; Liu, Q.; Zhu, R.; Cao, Z. Calmodulin as a Potential Target by Which Berberine Induces Cell Cycle Arrest in Human Hepatoma B el7402 Cells. Chem. Biol. Drug Des. 2013, 81, 775–783. [Google Scholar] [CrossRef]
- Liu, X.; Ouyang, S.; Yu, B.; Liu, Y.; Huang, K.; Gong, J.; Zheng, S.; Li, Z.; Li, H.; Jiang, H. PharmMapper server: A web server for potential drug target identification using pharmacophore mapping approach. Nucleic Acids Res. 2010, 38 (Suppl. 2), W609–W614. [Google Scholar] [CrossRef]
- Zhang, S.; Lu, W.; Liu, X.; Diao, Y.; Bai, F.; Wang, L.; Shan, L.; Huang, J.; Li, H.; Zhang, W. Fast and effective identification of the bioactive compounds and their targets from medicinal plants via computational chemical biology approach. MedChemComm 2011, 2, 471–477. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Xia, P.; Zhang, J.; Wang, Y.; Zhang, R.; Giesy, J.P.; Shi, W.; Yu, H. A high-throughput, computational system to predict if environmental contaminants can bind to human nuclear receptors. Sci. Total Environ. 2017, 576, 609–616. [Google Scholar] [CrossRef]
- Park, K.; Cho, A.E. Using reverse docking to identify potential targets for ginsenosides. J. Ginseng Res. 2017, 41, 534–539. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nature Protoc. 2008, 4, 44. [Google Scholar] [CrossRef]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; Von Mering, C. STRING v9. 1: Protein–protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2012, 41, D808–D815. [Google Scholar] [CrossRef]
- Li, J.; Zhou, N.; Luo, K.; Zhang, W.; Li, X.; Wu, C.; Bao, J. In silico discovery of potential VEGFR-2 inhibitors from natural derivatives for anti-angiogenesis therapy. Int. J. Mol. Sci. 2014, 15, 15994–16011. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- He, Z.; Tang, F.; Ermakova, S.; Li, M.; Zhao, Q.; Cho, Y.Y.; Ma, W.Y.; Choi, H.S.; Bode, A.M.; Yang, C.S. Fyn is a novel target of (−)-epigallocatechin gallate in the inhibition of JB6 Cl41 cell transformation. Mol. Carcinog. 2008, 47, 172–183. [Google Scholar] [CrossRef]
- Ahmed, S.; Rahman, A.; Hasnain, A.; Lalonde, M.; Goldberg, V.; Haqqi, T. Green tea polyphenol EGCG inhibits the IL-1 beta-induced activity an d expression of COX-2 and NOS-2 in human chondrocytes. Free Radic. Biol. Med. 2002, 33, 1097–1105. [Google Scholar] [CrossRef]
- Oliva, J.; Bardag-Gorce, F.; Tillman, B.; French, S.W. Protective effect of quercetin, EGCG, catechin and betaine against oxidative stress induced by ethanol in vitro. Exp. Mol. Pathol. 2011, 90, 295–299. [Google Scholar] [CrossRef]
- Hung, P.-F.; Wu, B.-T.; Chen, H.-C.; Chen, Y.-H.; Chen, C.-L.; Wu, M.-H.; Liu, H.-C.; Lee, M.-J.; Kao, Y.-H. The antimitogenic effect of green tea (−)-epigallocatechin gallate on 3T3-L1 preadipocytes depends on the Erk and Cdk2 pathways. Am. J. Physiol. Cell Physiol. 2005, 288, 1094–1108. [Google Scholar] [CrossRef]
- Wu, B.-T.; Hung, P.-F.; Chen, H.-C.; Huang, R.-N.; Chang, H.-H.; Kao, Y.-H. The apoptotic effect of green tea (−)-epigallocatechin gallate on 3T3-L1 preadipocytes depends on the Cdk2 pathway. J. Agric. Food Chem. 2005, 53, 5695–5701. [Google Scholar] [CrossRef]
- Chen, H.; Adams, E.; Van Schepdael, A. Study of Abl1 tyrosine kinase inhibitors by liquid chromatography–electrospray ionization-mass spectrometry. Talanta 2013, 107, 88–94. [Google Scholar] [CrossRef]
- Van Aller, G.S.; Carson, J.D.; Tang, W.; Peng, H.; Zhao, L.; Copeland, R.A.; Tummino, P.J.; Luo, L. Epigallocatechin gallate (EGCG), a major component of green tea, is a dual phosphoinositide-3-kinase/mTOR inhibitor. Biochem. Biophys. Res. Commun. 2011, 406, 194–199. [Google Scholar] [CrossRef]
- Park, S.B.; Bae, J.W.; Kim, J.M.; Lee, S.G.; Han, M. Antiproliferative and apoptotic effect of epigallocatechin-3-gallate on Ishikawa cells is accompanied by sex steroid receptor downregulation. Int. J. Mol. Med. 2012, 30, 1211–1218. [Google Scholar] [CrossRef]
- Singh, A.K.; Umar, S.; Riegsecker, S.; Chourasia, M.; Ahmed, S. Regulation of Transforming Growth Factor β–Activated Kinase Activation by Epigallocatechin-3-Gallate in Rheumatoid Arthritis Synovial Fibroblasts: Suppression of K63-Linked Autoubiquitination of Tumor Necrosis Factor Receptor–Associated Factor 6. Arthritis Rheumatol. 2016, 68, 347–358. [Google Scholar] [CrossRef]
- Wu, P.-P.; Kuo, S.-C.; Huang, W.-W.; Yang, J.-S.; Lai, K.-C.; Chen, H.-J.; Lin, K.-L.; Chiu, Y.-J.; Huang, L.-J.; Chung, J.-G. (–)-Epigallocatechin gallate induced apoptosis in human adrenal cancer NCI-H295 cells through caspase-dependent and caspase-independent pathway. Anticancer Res. 2009, 29, 1435–1442. [Google Scholar]
- Wang, W.; Wu, J.; Zhang, Q.; Li, X.; Zhu, X.; Wang, Q.; Cao, S.; Du, L. Mitochondria-mediated apoptosis was induced by oleuropein in H1299 cells involving activation of p38 MAP kinase. J. Cell. Biochem. 2019, 120, 5480–5494. [Google Scholar] [CrossRef]
- Li, X.; Sun, R.; Chen, W.; Lu, B.; Li, X.; Wang, Z.; Bao, J. A systematic in silico mining of the mechanistic implications and therapeutic potentials of estrogen receptor (ER)-α in breast cancer. PLoS ONE 2014, 9, e91894. [Google Scholar] [CrossRef]
- Wang, W.; Li, X.; Wang, Q.; Zhu, X.; Zhang, Q.; Du, L. The acidic pH-induced structural changes in apo-CP43 by spectral methodologies and molecular dynamics simulations. J. Mol. Struct. 2018, 1152, 177–188. [Google Scholar] [CrossRef]
- Zhao, M.-L.; Wang, W.; Nie, H.; Cao, S.-S.; Du, L.-F. In silico structure prediction and inhibition mechanism studies of AtHDA14 as revealed by homology modeling, docking, molecular dynamics simulation. Comput. Biol. Chem. 2018, 75, 120–130. [Google Scholar] [CrossRef]
- Yang, F.; Oz, H.S.; Barve, S.; De Villiers, W.J.; McClain, C.J.; Varilek, G.W. The green tea polyphenol (−)-epigallocatechin-3-gallate blocks nuclear factor-κB activation by inhibiting IκB kinase activity in the intestinal epithelial cell line IEC-6. Mol. Pharmacol. 2001, 60, 528–533. [Google Scholar]
- Oz, H.S.; Ebersole, J.L. Green tea polyphenols mediated apoptosis in intestinal epithelial cells by a FADD-dependent pathway. J. Cancer Ther. 2010, 1, 105. [Google Scholar] [CrossRef]
- Masuda, M.; Suzui, M.; Lim, J.T.; Weinstein, I.B. Epigallocatechin-3-gallate inhibits activation of HER-2/neu and downstream signaling pathways in human head and neck and breast carcinoma cells. Clin. Cancer Res. 2003, 9, 3486–3491. [Google Scholar]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef]
- Shankar, S.; Marsh, L.; Srivastava, R.K. EGCG inhibits growth of human pancreatic tumors orthotopically implanted in Balb C nude mice through modulation of FKHRL1/FOXO3a and neuropilin. Mol. Cell. Biochem. 2013, 372, 83–94. [Google Scholar] [CrossRef]
- Senggunprai, L.; Kukongviriyapan, V.; Prawan, A.; Kukongviriyapan, U. Quercetin and EGCG exhibit chemopreventive effects in cholangiocarcinoma cells via suppression of JAK/STAT signaling pathway. Phytother. Res. 2014, 28, 841–848. [Google Scholar] [CrossRef]
- Lauro, G.; Romano, A.; Riccio, R.; Bifulco, G. Inverse virtual screening of antitumor targets: Pilot study on a small database of natural bioactive compounds. J. Nat. Prod. 2011, 74, 1401–1407. [Google Scholar] [CrossRef]
- Verma, N.; Rai, A.K.; Kaushik, V.; Brünnert, D.; Chahar, K.R.; Pandey, J.; Goyal, P. Identification of gefitinib off-targets using a structure-based systems biology approach; their validation with reverse docking and retrospective data mining. Sci. Rep. 2016, 6, 33949. [Google Scholar] [CrossRef]
- Zahler, S.; Tietze, S.; Totzke, F.; Kubbutat, M.; Meijer, L.; Vollmar, A.M.; Apostolakis, J. Inverse in silico screening for identification of kinase inhibitor targets. Chem. Biol. 2007, 14, 1207–1214. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39 (Suppl. 2), W316–W322. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Hess, B.; Kutzner, C.; Van Der Spoel, D.; Lindahl, E. GROMACS 4: Algorithms for highly efficient, load-balanced and scalable molecular simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J. GROMACS: Fast, flexible and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Wang, X.; Lu, K.; Luo, H.; Liang, D.; Long, X.; Yuan, Y.; Wu, C.; Bao, J. In silico identification of small molecules as novel LXR agonists for the treatment of cardiovascular disease and cancer. J. Mol. Model. 2018, 24, 57. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compound are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB_ID | Symbol | Uniprot_ID | Ligand_ID | Lig_Score | EGCG_Score | Experiment |
|---|---|---|---|---|---|---|
| 3UYT | CSNK1D | P48730 | 0CK | −9.6 | −9.2 | No |
| 4B6L | PLK3 | Q9H4B4 | 9ZP | −8.2 | −9.2 | No |
| 4K4E | TNKS | O95271 | 4KE | −9.2 | −9.2 | No |
| 3MVH | AKT1 | P31749 | WFE | −1.8 | −9.1 | Yes, [1,45] |
| 2CLQ | MAP3K5 | Q99683 | STU | −12.0 | −9.1 | No |
| 3GFT | KRAS | P01116 | GNP | −7.6 | −9.1 | No |
| 4TVJ | PARP2 | Q9UGN5 | 09L | −12.5 | −10.8 | No |
| 2DQ7 | FYN | P06241 | STU | −12.1 | −10.0 | Yes, [39] |
| 4KIK | IKBKB | O14920 | KSA | −14.0 | −10.0 | No |
| 3EQR | TNK2 | Q07912 | T74 | −10.2 | −9.9 | No |
| 3MTF | ACVR1 | Q04771 | A3F | −9.3 | −9.9 | No |
| 4GV0 | PARP3 | Q9Y6F1 | 8ME | −9.4 | −9.9 | No |
| 1M6I | AIFM1 | O95831 | FAD | −7.4 | −9.8 | No |
| 1UA2 | CDK7 | P50613 | ATP | −8.0 | −9.8 | No |
| 1Z6T | APAF1 | O14727 | ADP | −8.1 | −9.8 | Yes, [48] |
| 1ZXM | TOP2A | P11388 | ANP | −8.2 | −9.8 | No |
| 2WQN | NEK7 | Q8TDX7 | ADP | −8.0 | −9.6 | No |
| 4NOS | NOS2 | P35228 | H2B | −4.0 | −9.6 | Yes, [40,41] |
| 4R8Q | BUB1 | O43683 | ADP | −7.1 | −9.6 | No |
| 1YVJ | JAK3 | P52333 | 4ST | −11.9 | −9.5 | No |
| 2IW9 | CDK2 | P24941 | 4SP | −7.2 | −9.5 | Yes, [42,43] |
| 2V7A | ABL1 | P00519 | 627 | −7.6 | −9.5 | Yes, [44] |
| 4I5M | PLK2 | Q9NYY3 | R78 | −8.4 | −9.50 | No |
| 6CIC | NOS1 | P29475 | 7R2 | −8.8 | −9.50 | No |
| 1XBC | SYK | P43405 | STU | −12.5 | −9.4 | Yes, [8] |
| 2JDR | AKT2 | P31751 | L20 | −10.2 | −9.4 | Yes, [1,45] |
| 2JED | PRKCQ | Q04759 | LG8 | −10.8 | −9.4 | No |
| 3ALN | MAP2K4 | P45985 | ANP | −7.9 | −9.4 | No |
| 3ELJ | MAPK8 | P45983 | GS7 | −8.0 | −9.4 | Yes, [46] |
| 3NR9 | CLK2 | P49760 | NR9 | −8.7 | −9.4 | No |
| 4IFC | PRPF4B | Q13523 | ADP | −8.0 | −9.4 | No |
| 1X8B | WEE1 | P30291 | 824 | −11.6 | −9.3 | No |
| 2J8Z | TP53I3 | Q53FA7 | NAP | −7.8 | −9.3 | No |
| 2NRU | IRAK4 | Q9NWZ3 | T12 | −8.4 | −9.3 | Yes, [47] |
| 3OP5 | VRK1 | Q99986 | REB | −2.1 | −9.3 | No |
| 4AOJ | NTRK1 | P04629 | V4Z | −8.8 | −9.3 | No |
| 5DN3 | AURKA | O14965 | 5DN | −9.5 | −9.3 | No |
| 5YQX | BRD4 | O60885 | E0K | −7.6 | −9.3 | No |
| 2XIK | STK25 | O00506 | J60 | −1.9 | −9.2 | No |
| 3JYA | PIM1 | P11309 | LWG | −8.3 | −9.2 | No |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Xiong, X.; Li, X.; Zhang, Q.; Yang, W.; Du, L. In Silico Investigation of the Anti-Tumor Mechanisms of Epigallocatechin-3-Gallate. Molecules 2019, 24, 1445. https://doi.org/10.3390/molecules24071445
Wang W, Xiong X, Li X, Zhang Q, Yang W, Du L. In Silico Investigation of the Anti-Tumor Mechanisms of Epigallocatechin-3-Gallate. Molecules. 2019; 24(7):1445. https://doi.org/10.3390/molecules24071445
Chicago/Turabian StyleWang, Wang, Xiuhong Xiong, Xue Li, Qinyang Zhang, Wentao Yang, and Linfang Du. 2019. "In Silico Investigation of the Anti-Tumor Mechanisms of Epigallocatechin-3-Gallate" Molecules 24, no. 7: 1445. https://doi.org/10.3390/molecules24071445
APA StyleWang, W., Xiong, X., Li, X., Zhang, Q., Yang, W., & Du, L. (2019). In Silico Investigation of the Anti-Tumor Mechanisms of Epigallocatechin-3-Gallate. Molecules, 24(7), 1445. https://doi.org/10.3390/molecules24071445