
Quantification of Furosine (Nε-(2-Furoylmethyl)-l-lysine) in Different Parts of Velvet Antler with Various Processing Methods and Factors Affecting Its Formation


Abstract
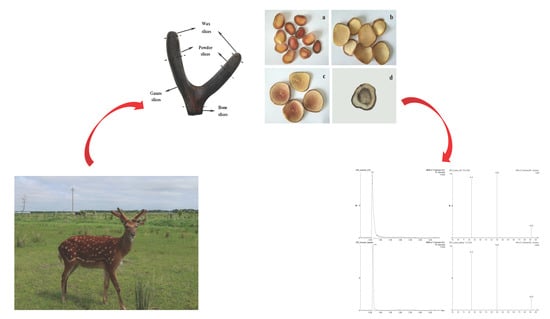
1. Introduction
2. Results and Discussion
2.1. Sample Pre-Treatment and Chromatography Conditions
2.2. Method Validation
2.3. Furosine Content in the Processed Velvet Antler
2.4. Factors Influencing the Furosine Content in the Processed Velvet Antler
2.4.1. Protein and Amino Acid Contents in the Processed Velvet Antler
2.4.2. Reducing Sugar Content in the Processed Velvet Antler
3. Materials and Methods
3.1. Materials
3.2. Sources and Preparation of the Velvet Antler
3.3. Preparation of the Different Processed Velvet Antler Slices
3.4. Sample Preparation
3.5. Quantification of Furosine
3.6. Protein Content Analysis
3.7. Amino Acid Content Analysis
3.8. Reducing Sugar Content Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jeon, B.; Kim, S.; Lee, S.; Park, P.; Sung, S.; Kim, J.; Moon, S. Effect of antler growth period on the chemical composition of velvet antler in sika deer (Cervus nippon). Z. Saugetierkd. 2009, 74, 374–380. [Google Scholar] [CrossRef]
- Wu, F.; Li, H.; Jin, L.; Li, X.; Ma, Y.; You, J.; Li, S.; Xu, Y. Deer antler base as a traditional Chinese medicine: A review of its traditional uses, chemistry and pharmacology. J. Ethnopharmacol. 2013, 145, 403–415. [Google Scholar] [CrossRef]
- Zhou, R.; Li, S. In vitro antioxidant analysis and characterisation of antler velvet extract. Food Chem. 2009, 114, 1321–1327. [Google Scholar] [CrossRef]
- Sui, Z.; Yuan, H.; Liang, Z.; Zhao, Q.; Wu, Q.; Xia, S.; Zhang, L.; Huo, Y.; Zhang, Y. An activity-maintaining sequential protein extraction method for bioactive assay and proteome analysis of velvet antlers. Talanta 2013, 107, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Sui, Z.; Zhang, L.; Huo, Y.; Zhang, Y. Bioactive components of velvet antlers and their pharmacological properties. J. Pharm. Biomed. 2014, 87, 229–240. [Google Scholar] [CrossRef]
- Tseng, S.H.; Sung, C.H.; Chen, L.G.; Lai, Y.J.; Chang, W.S.; Sung, H.C.; Wang, C.C. Comparison of chemical compositions and osteoprotective effects of different sections of velvet antler. J. Ethnopharmacol. 2014, 151, 352–360. [Google Scholar] [CrossRef]
- Singh, R.; Barden, A.; Mori, T.; Beilin, L. Advanced glycation end-products: A review. Diabetologia 2001, 2001, 129–146. [Google Scholar] [CrossRef]
- Reynaert, N.L.; Gopal, P.; Rutten, E.P.A.; Wouters, E.F.M.; Schalkwijk, C.G. Advanced glycation end products and their receptor in age-related, non-communicable chronic inflammatory diseases; Overview of clinical evidence and potential contributions to disease. Int. J. Biochem. Cell Biol. 2016, 81, 403–418. [Google Scholar] [CrossRef]
- Kuhla, A.; Ludwig, S.C.; Kuhla, B.; Münch, G.; Vollmar, B. Advanced glycation end products are mitogenic signals and trigger cell cycle reentry of neurons in Alzheimer’s disease brain. Neurobiol. Aging 2015, 36, 753–761. [Google Scholar] [CrossRef] [PubMed]
- de Vos, L.C.; Lefrandt, J.D.; Dullaart, R.P.; Zeebregts, C.J.; Smit, A.J. Advanced glycation end products: An emerging biomarker for adverse outcome in patients with peripheral artery disease. Atherosclerosis 2016, 254, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Drenth, H.; Zuidema, S.U.; Krijnen, W.P.; Bautmans, I.; Schans, C.V.D.; Hobbelen, H. Advanced glycation end-products are associated with the presence and severity of paratonia in early stage Alzheimer disease. J. Am. Med. Dir. Assoc. 2017, 18, 636.e7–636.e12. [Google Scholar] [CrossRef] [PubMed]
- Gautieri, A.; Passini, F.S.; Silván, U.; Guizar-Sicairos, M.; Carimati, G.; Volpi, P.; Moretti, M.; Redaelli, A.; Berli, M.; Snedeker, J.G. Advanced glycation end-products: Mechanics of aged collagen from molecule to tissue. Matrix Biol. 2016, 59, 95–108. [Google Scholar] [CrossRef]
- Vlassara, H.; Palace, M.R. Glycoxidation: The menace of diabetes and aging. Mt. Sinai J. Med. 2003, 70, 232–241. [Google Scholar]
- Chuang, Y.C.; Wu, M.S.; Wu, T.H.; Su, Y.K.; Lee, Y.M. Pyridoxamine ameliorates the effects of advanced glycation end products on subtotal nephrectomy induced chronic renal failure rats. J. Funct. Foods 2012, 4, 679–686. [Google Scholar] [CrossRef]
- Buraczynska, M.; Zaluska, W.; Buraczynska, K.; Markowska-Gosik, D.; Ksiazek, A. Receptor for advanced glycation end products (RAGE) gene polymorphism and cardiovascular disease in end-stage renal disease patients. Hum. Immunol. 2015, 76, 843–848. [Google Scholar] [CrossRef]
- Hashimoto, K.; Kunikata, H.; Yasuda, M.; Ito, A.; Aizawa, N.; Sawada, S.; Kondo, K.; Satake, C.; Takano, Y.; Nishiguchi, K.M. The relationship between advanced glycation end products and ocular circulation in type 2 diabetes. J. Diabetes Complicat. 2016, 30, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Moriya, S.; Yamazaki, M.; Murakami, H.; Maruyama, K.; Uchiyama, S. Two soluble isoforms of receptors for advanced glycation end products (RAGE) in carotid Atherosclerosis: The difference of soluble and endogenous secretory RAGE. J. Stroke Cerebrovasc. 2014, 23, 2540–2546. [Google Scholar] [CrossRef] [PubMed]
- Gong, R.Z.; Wang, Y.; Wang, Y.F.; Chen, B.; Gao, K.; Sun, Y.S. Simultaneous Determination of Nε-(carboxymethyl) lysine and Nε-(carboxyethyl) lysine in different sections of antler velvet after various processing methods by UPLC-MS/MS. Molecules 2018, 23, 3316. [Google Scholar] [CrossRef]
- Troise, A.D.; Fiore, A.; Wiltafsky, M.; Fogliano, V. Quantification of Nε-(2-Furoylmethyl)-l-lysine (furosine), Nε-(Carboxymethyl)-l-lysine (CML), Nε-(Carboxyethyl)-l-lysine (CEL) and total lysine through stable isotope dilution assay and tandem mass spectrometry. Food Chem. 2015, 188, 357–364. [Google Scholar] [CrossRef]
- He, J.; Zeng, M.; Zheng, Z.; He, Z.; Chen, J. Simultaneous determination of Nε-(carboxymethyl) lysine and Nε-(carboxyethyl) lysine in cereal foods by LC–MS/MS. Eur. Food Res. Technol. 2014, 238, 367–374. [Google Scholar] [CrossRef]
- Hull, G.L.J.; Woodside, J.V.; Ames, J.M.; Cuskelly, G.J. Nε-(carboxymethyl)lysine content of foods commonly consumed in a Western style diet. Food Chem. 2012, 131, 170–174. [Google Scholar] [CrossRef]
- Erbersdobler, H.F.; Somoza, V. Forty years of furosine—Forty years of using Maillard reaction products as indicators of the nutritional quality of foods. Mol. Nutr. Food Res. 2010, 51, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.; Beaulieu, L.P.; Fraser, M.H.; Mcintyre, K.; Owen, P.; Martineau, L.; Cuerrier, A.; Johns, T.; Haddad, P.S.; Bennett, S.A.L.; et al. Inhibition of advanced glycation end product formation by medicinal plant extracts correlates with phenolic metabolites and antioxidant activity. Planta Med. 2010, 77, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Xing, L.; Wang, J.Q.; Zheng, N. Toxicology studies of furosine in vitro/in vivo and exploration of the related mechanism. Toxicol. Lett. 2018, 291, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Bosch, L.; Alegrı, A.A.; Farré, R.; Clemente, G. Effect of storage conditions on furosine formation in milk–cereal based baby foods. Food Chem. 2008, 107, 1681–1686. [Google Scholar] [CrossRef]
- Villamiel, M.; Castillo, M.D.D.; Corzo, N.; Olano, A. Presence of furosine in honeys. J. Sci. Food Agric. 2001, 81, 790–793. [Google Scholar] [CrossRef]
- Trevisan, A.J.; de Almeida Lima, D.; Sampaio, G.R.; Soares, R.A.; Markowicz Bastos, D.H. Influence of home cooking conditions on Maillard reaction products in beef. Food Chem. 2016, 196, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; He, J.; Li, F.; Tao, G.; Zhang, S.; Zhang, S.; Qin, F.; Zeng, M.; Chen, J. Nε-(carboxymethyl)lysine and Nε-(carboxyethyl)lysine in tea and the factors affecting their formation. Food Chem. 2017, 232, 683–688. [Google Scholar] [CrossRef]
- Schettgen, T.; Tings, A.; Brodowsky, C.; Müller-Lux, A.; Musiol, A.; Kraus, T. Simultaneous determination of the advanced glycation end product Nε-carboxymethyllysine and its precursor, lysine, in exhaled breath condensate using isotope-dilution-hydrophilic-interaction liquid chromatography coupled to tandem mass spectrometry. Anal. Bioanal. Chem. 2007, 387, 2783–2791. [Google Scholar] [CrossRef]
- Renterghem, R.V.; Block, J.D. Furosine in consumption milk and milk powders. Int. Dairy J. 1996, 6, 371–382. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Nomi, Y.; Homma, T.; Kasai, M.; Otsuka, Y. Determination of furosine and fluorescence as markers of the Maillard reaction for the evaluation of meat products during actual cooking conditions. Food Sci. Technol. Res. 2012, 18, 67–76. [Google Scholar] [CrossRef]
- Dittrich, R.; Hoffmann, I.; Stahl, P.; Müller, A.; Beckmann, M.W.; Pischetsrieder, M. Concentrations of Nε-carboxymethyllysine in human breast milk, infant formulas, and urine of infants. J. Agric. Food Chem. 2006, 54, 6924–6928. [Google Scholar] [CrossRef]
- Li, C.; Zhao, H.; Liu, Z.; Mcmahon, C. Deer antler—A novel model for studying organ regeneration in mammals. Int. J. Biochem. Cell Biol. 2014, 56, 111–122. [Google Scholar] [CrossRef]
- Xu, X.B.; Ma, F.; Yu, S.J.; Guan, Y.G. Simultaneous analysis of Nε-(carboxymethyl)lysine, reducing sugars, and lysine during the dairy thermal process. J. Dairy Sci. 2013, 96, 5487–5493. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, X.; Meng, L.; Wang, Y. Qualitative and quantitative analysis of furosine in fresh and processed ginsengs. J. Ginseng Res. 2018, 42, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, R.S.S.; Silva, A.S.A.D.; Bon, E.P.D.S. Amino acids interference on the quantification of reducing sugars by the 3,5-dinitrosalicylic acid assay mislead carbohydrase activity measurements. Carbohydr. Res. 2012, 363, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Busardò, F.P.; Gottardi, M.; Tini, A.; Mortali, C.; Giorgetti, R.; Pichini, S. Ultra-high-performance liquid chromatography tandem mass spectrometry assay for determination of endogenous GHB and GHB-glucuronide in nails. Molecules 2018, 10, 2686. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Kobayashi, M.; Kobayashi, A.; Kobayashi, K.; Asamura, H. High-frequency heating extraction method for sensitive drug analysis in human nails. Molecules 2018, 12, 3231. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds furosine, mannose, glucosamine, ribose, glucuronic acid, galacturonic acid, aminogalactose, glucose, galactose and 17 amino acid standards are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Processing Methods | Parts | mg FML/kg Protein a | mmol FML/mol Lysine b | mg FML/kg |
|---|---|---|---|---|
| freeze-dried | wax slices | 193.94 ± 1.21 | 0.53 ± 0.14 | 138.77 ± 1.78 |
| powder slices | 155.05 ± 1.43 | 0.50 ± 0.09 | 88.56 ± 1.84 | |
| gauze slices | 154.69 ± 1.19 | 0.49 ± 0.21 | 81.58 ± 1.23 | |
| bone slices | 153.68 ± 1.98 | 0.50 ± 0.16 | 75.35 ± 1.42 | |
| entire | 148.51 ± 1.85 | 0.47 ± 0.20 | 84.54 ± 1.53 | |
| boiled | wax slices | 241.22 ± 2.13 | 0.79 ± 0.23 | 155.35 ± 1.68 |
| powder slices | 226.21 ± 1.98 | 0.67 ± 0.22 | 137.27 ± 1.72 | |
| gauze slices | 202.98 ± 1.84 | 0.62 ± 0.19 | 109.59 ± 1.66 | |
| bone slices | 155.02 ± 1.78 | 0.57 ± 0.24 | 79.26 ± 2.03 | |
| entire | 168.10 ± 1.22 | 0.56 ± 0.26 | 102.54 ± 1.22 | |
| processed without blood | wax slices | 80.33 ± 1.25 | 0.26 ± 0.12 | 64.11 ± 2.04 |
| powder slices | 61.65 ± 1.31 | 0.20 ± 0.14 | 34.96 ± 1.75 | |
| gauze slices | 49.26 ± 1.43 | 0.17 ± 0.21 | 29.01 ± 2.10 | |
| bone slices | 50.35 ± 1.13 | 0.17 ± 0.19 | 27.24 ± 1.45 | |
| entire | 60.29 ± 1.46 | 0.25 ± 0.17 | 34.02 ± 1.58 | |
| processed with blood | wax slices | 139.88 ± 1.87 | 0.44 ± 0.22 | 106.62 ± 1.95 |
| powder slices | 126.42 ± 1.54 | 0.42 ± 0.19 | 83.89 ± 1.63 | |
| gauze slices | 122.46 ± 1.44 | 0.41 ± 0.25 | 75.25 ± 1.39 | |
| bone slices | 129.15 ± 1.32 | 0.35 ± 0.17 | 64.13 ± 1.88 | |
| entire | 115.18 ± 2.13 | 0.38 ± 0.14 | 68.24 ± 1.21 |
| Processing Methods | Parts | Protein/% | Total Amino Acids/g/100 g | Lysine/g/100 g |
|---|---|---|---|---|
| freeze-dried | wax slices | 81.56 ± 0.04 | 88.64 ± 3.30 | 5.87 ± 0.20 |
| powder slices | 57.12 ± 0.03 | 62.51 ± 1.84 | 3.96 ± 0.11 | |
| gauze slices | 52.74 ± 0.10 | 61.75 ± 0.90 | 3.76 ± 0.03 | |
| bone slices | 49.03 ± 0.25 | 58.20 ± 1.17 | 3.41 ± 0.21 | |
| entire | 56.93 ± 0.34 | 68.00 ± 1.23 | 4.02 ± 0.13 | |
| boiled | wax slices | 81.25 ± 0.12 | 87.48 ± 1.78 | 5.67 ± 0.22 |
| powder slices | 58.11 ± 0.18 | 62.29 ± 0.82 | 3.92 ± 0.14 | |
| gauze slices | 53.99 ± 0.33 | 60.97 ± 0.28 | 3.69 ± 0.10 | |
| bone slices | 51.13 ± 0.25 | 55.62 ± 1.74 | 3.15 ± 0.12 | |
| entire | 60.99 ± 0.44 | 67.59 ± 1.82 | 4.09 ± 0.29 | |
| processed without blood | wax slices | 79.81 ± 0.09 | 86.74 ± 0.18 | 5.56 ± 0.11 |
| powder slices | 56.69 ± 0.11 | 66.65 ± 0.74 | 4.01 ± 0.09 | |
| gauze slices | 58.88 ± 0.31 | 62.35 ± 0.48 | 3.76 ± 0.11 | |
| bone slices | 54.11 ± 0.24 | 52.34 ± 0.72 | 3.01 ± 0.12 | |
| entire | 56.43 ± 0.28 | 63.55 ± 0.82 | 3.58 ± 0.15 | |
| processed with blood | wax slices | 82.09 ± 0.74 | 91.02 ± 0.46 | 5.66 ± 0.12 |
| powder slices | 61.49 ± 0.33 | 74.06 ± 0.34 | 4.81 ± 0.02 | |
| gauze slices | 61.45 ± 0.41 | 66.94 ± 0.49 | 4.26 ± 0.07 | |
| bone slices | 49.65 ± 0.56 | 61.06 ± 0.46 | 3.48 ± 0.15 | |
| entire | 59.25 ± 0.35 | 64.31 ± 0.56 | 4.03 ± 0.17 |
| Compound | Freeze-Dried | Boiled | ||||||
| Wax Slices | Powder Slices | Gauze Slices | Bone Slices | Wax Slices | Powder Slices | Gauze Slices | Bone Slices | |
| mannose | 13.31 ± 1.01 | 9.49 ± 0.78 | 9.75 ± 0.65 | 7.66 ± 0.55 | 10.10 ± 0.98 | 6.32 ± 0.42 | 5.86 ± 0.45 | 4.31 ± 0.33 |
| glucosamine | 70.21 ± 2.03 | 60.32 ± 1.84 | 58.45 ± 1.21 | 48.04 ± 1.97 | 66.34 ± 2.33 | 56.30 ± 1.04 | 51.75 ± 1.32 | 44.04 ± 1.74 |
| ribose | 14.16 ± 1.21 | 9.37 ± 0.96 | 7.94 ± 1.01 | 6.60 ± 0.95 | 7.63 ± 1.04 | 5.52 ± 0.97 | 4.07 ± 0.87 | 3.58 ± 0.74 |
| glucuronic acid | 20.02 ± 1.19 | 16.51 ± 1.20 | 12.09 ± 0.98 | 7.46 ± 0.77 | 15.34 ± 1.04 | 13.74 ± 0.96 | 8.05 ± 0.81 | 6.18 ± 0.77 |
| galacturonic acid | 3.55 ± 0.94 | 2.64 ± 0.81 | 1.26 ± 0.72 | 0.27 ± 0.18 | 2.75 ± 0.55 | 1.52 ± 0.53 | 1.29 ± 0.19 | 0.20 ± 0.12 |
| aminogalactose | 18.36 ± 1.32 | 15.77 ± 1.09 | 11.39 ± 0.75 | 3.96 ± 0.63 | 16.98 ± 0.94 | 11.32 ± 1.02 | 9.88 ± 0.91 | 2.39 ± 0.74 |
| glucose | 18.10 ± 1.52 | 13.46 ± 1.04 | 11.95 ± 1.22 | 10.13 ± 1.09 | 16.25 ± 1.43 | 10.86 ± 1.51 | 9.24 ± 1.06 | 7.13 ± 1.33 |
| galactose | 28.89 ± 1.82 | 26.54 ± 2.07 | 26.58 ± 1.67 | 22.33 ± 1.33 | 22.93 ± 1.47 | 16.62 ± 1.83 | 16.75 ± 1.42 | 19.43 ± 1.17 |
| Compound | Processed with Blood | Processed without Blood | ||||||
| Wax Slices | Powder Slices | Gauze Slices | Bone Slices | Wax Slices | Powder Slices | Gauze Slices | Bone Slices | |
| mannose | 11.20 ± 1.07 | 9.60 ± 1.21 | 6.30 ± 0.97 | 5.83 ± 0.84 | 9.22 ± 1.07 | 7.15 ± 0.92 | 4.77 ± 0.67 | 3.22 ± 0.54 |
| glucosamine | 52.34 ± 2.07 | 35.47 ± 2.22 | 31.34 ± 1.98 | 28.10 ± 2.13 | 57.36 ± 2.45 | 38.11 ± 1.56 | 33.71 ± 1.42 | 30.13 ± 1.08 |
| ribose | 5.63 ± 0.71 | 4.78 ± 0.63 | 3.22 ± 0.58 | 2.10 ± 0.41 | 4.17 ± 0.51 | 3.90 ± 0.35 | 2.48 ± 0.24 | 1.16 ± 0.16 |
| glucuronic acid | 15.14 ± 0.74 | 13.83 ± 0.81 | 11.62 ± 0.53 | 5.01 ± 0.37 | 13.55 ± 1.02 | 8.76 ± 0.91 | 7.01 ± 0.87 | 3.65 ± 0.54 |
| galacturonic acid | 2.55 ± 0.32 | 1.90 ± 0.28 | 1.62 ± 0.17 | 0.82 ± 0.19 | 2.19 ± 0.23 | 1.72 ± 0.31 | 1.60 ± 0.22 | 0.20 ± 0.12 |
| aminogalactose | 16.78 ± 0.79 | 12.92 ± 0.82 | 9.46 ± 0.77 | 7.06 ± 0.54 | 20.24 ± 0.86 | 16.77 ± 0.72 | 12.34 ± 0.83 | 0.57 ± 0.14 |
| glucose | 16.25 ± 0.44 | 12.68 ± 0.58 | 9.21 ± 0.79 | 7.38 ± 0.88 | 13.65 ± 0.92 | 9.50 ± 0.77 | 7.62 ± 0.81 | 5.91 ± 0.69 |
| galactose | 22.73 ± 1.07 | 18.23 ± 0.98 | 16.44 ± 1.21 | 13.07 ± 0.78 | 18.08 ± 1.23 | 17.59 ± 1.42 | 15.22 ± 1.33 | 12.04 ± 0.79 |
| Compound | Precursor Ion (m/z) | Product Ion (m/z) | Cone Voltage (V) | Collision Energy (ev) | Dwell Time (ms) |
|---|---|---|---|---|---|
| furosine | 255 | 192 | 25 | 15 | 36 |
| 255 | 130 | 25 | 15 | 36 | |
| 255 | 84 | 25 | 25 | 36 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, R.-z.; Wang, Y.-h.; Gao, K.; Zhang, L.; Liu, C.; Wang, Z.-s.; Wang, Y.-f.; Sun, Y.-s. Quantification of Furosine (Nε-(2-Furoylmethyl)-l-lysine) in Different Parts of Velvet Antler with Various Processing Methods and Factors Affecting Its Formation. Molecules 2019, 24, 1255. https://doi.org/10.3390/molecules24071255
Gong R-z, Wang Y-h, Gao K, Zhang L, Liu C, Wang Z-s, Wang Y-f, Sun Y-s. Quantification of Furosine (Nε-(2-Furoylmethyl)-l-lysine) in Different Parts of Velvet Antler with Various Processing Methods and Factors Affecting Its Formation. Molecules. 2019; 24(7):1255. https://doi.org/10.3390/molecules24071255
Chicago/Turabian StyleGong, Rui-ze, Yan-hua Wang, Kun Gao, Lei Zhang, Chang Liu, Ze-shuai Wang, Yu-fang Wang, and Yin-shi Sun. 2019. "Quantification of Furosine (Nε-(2-Furoylmethyl)-l-lysine) in Different Parts of Velvet Antler with Various Processing Methods and Factors Affecting Its Formation" Molecules 24, no. 7: 1255. https://doi.org/10.3390/molecules24071255
APA StyleGong, R.-z., Wang, Y.-h., Gao, K., Zhang, L., Liu, C., Wang, Z.-s., Wang, Y.-f., & Sun, Y.-s. (2019). Quantification of Furosine (Nε-(2-Furoylmethyl)-l-lysine) in Different Parts of Velvet Antler with Various Processing Methods and Factors Affecting Its Formation. Molecules, 24(7), 1255. https://doi.org/10.3390/molecules24071255