
The Pros and Cons of Cystic Fibrosis (CF) Patient Use of Herbal Supplements Containing Pulmonaria officinalis L. Extract: the Evidence from an In Vitro Study on Staphylococcus aureus CF Clinical Isolates

 ,
,  , ,
, ,  ,
,
Abstract

1. Introduction
2. Results and Discussion
2.1. Short Description of Pulmonaria Officinalis L. Fractionated Extract
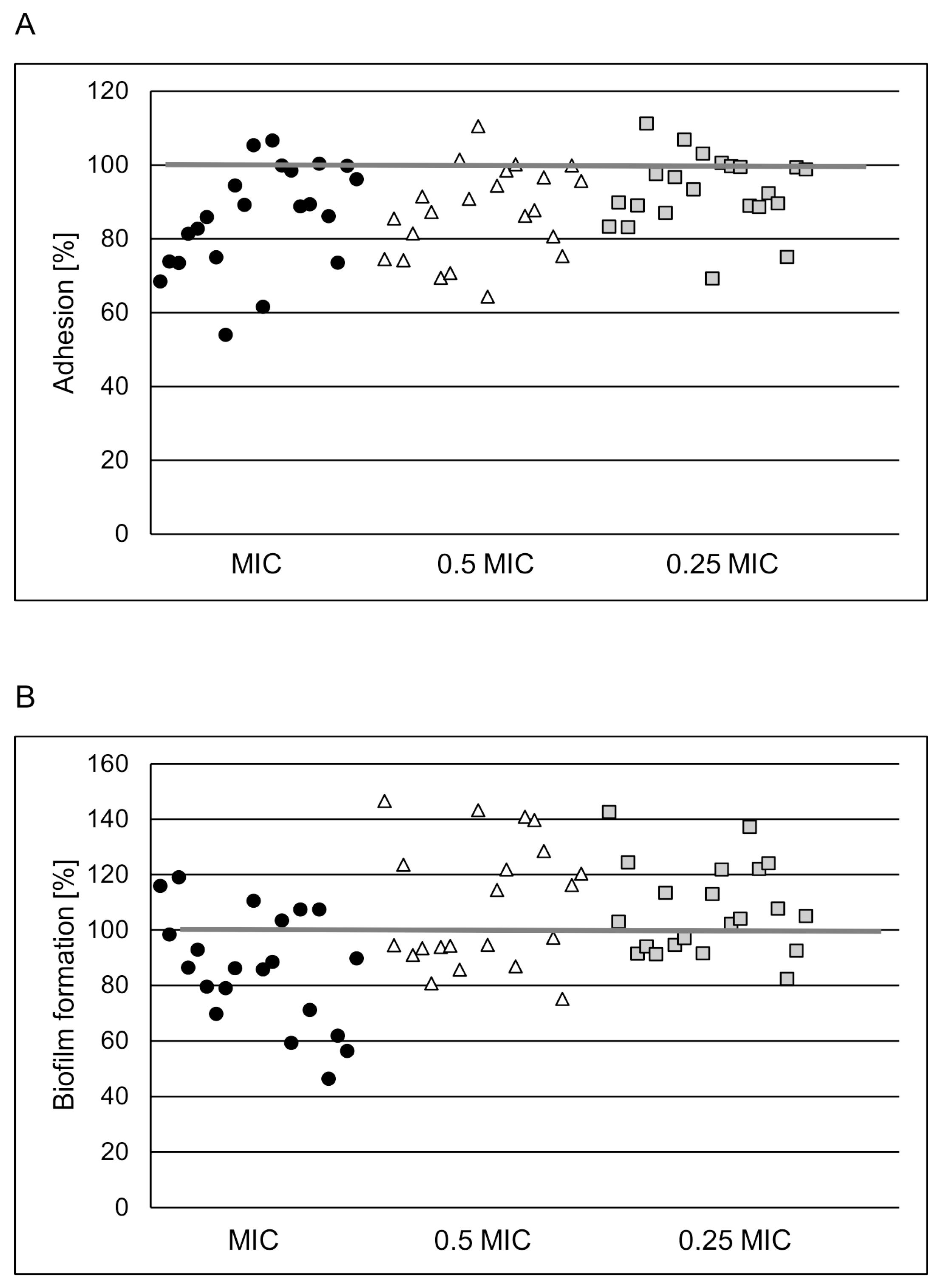
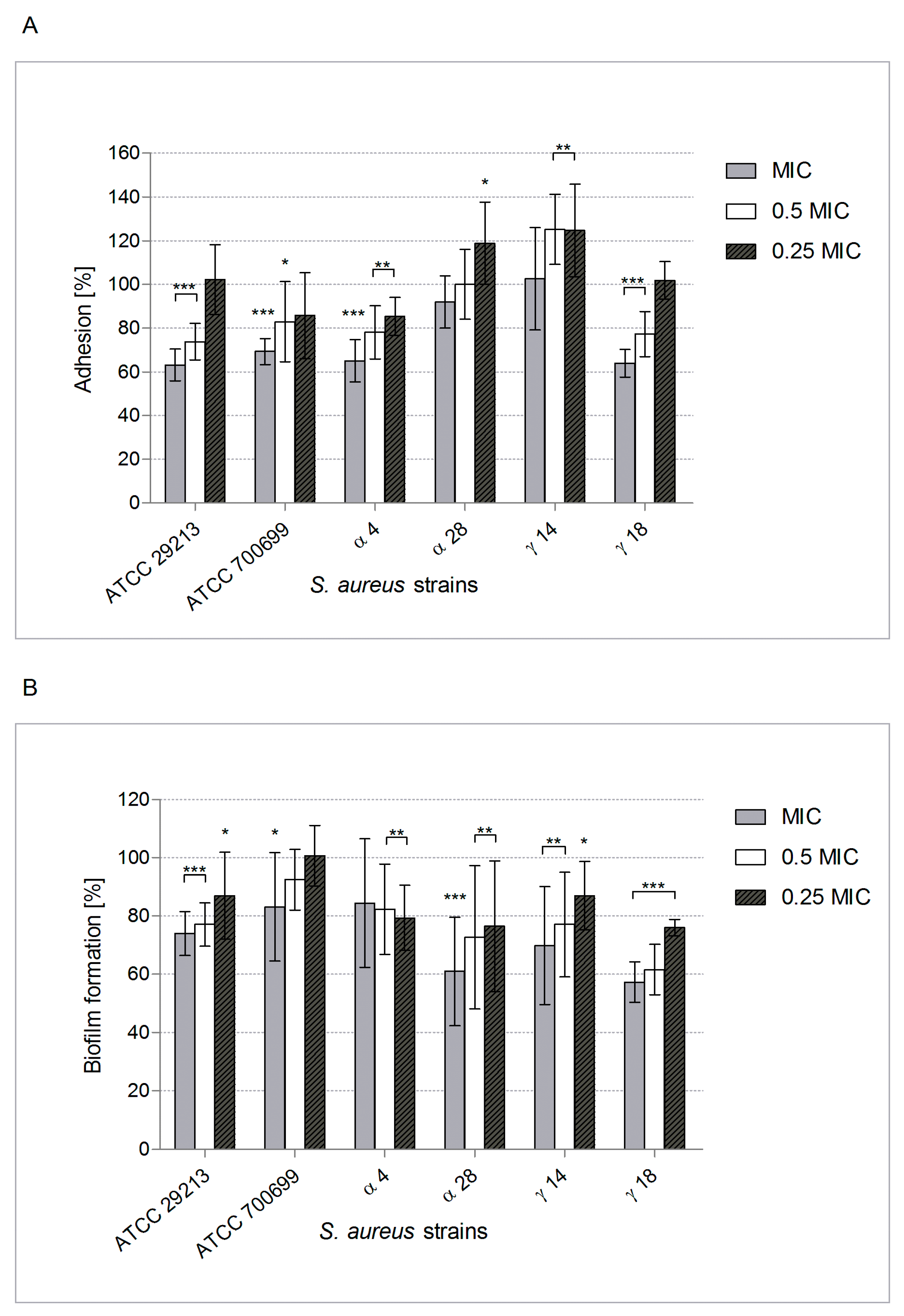
2.2. POE Antiadhesive and Anti-Biofilm Properties
2.3. POE Impact on Some of the Main Staphylococcus Aureus Virulence Attributes
3. Materials and Methods
3.1. Pulmonaria Officinalis Fractionated Extract (POE)—Preparation and Characterization
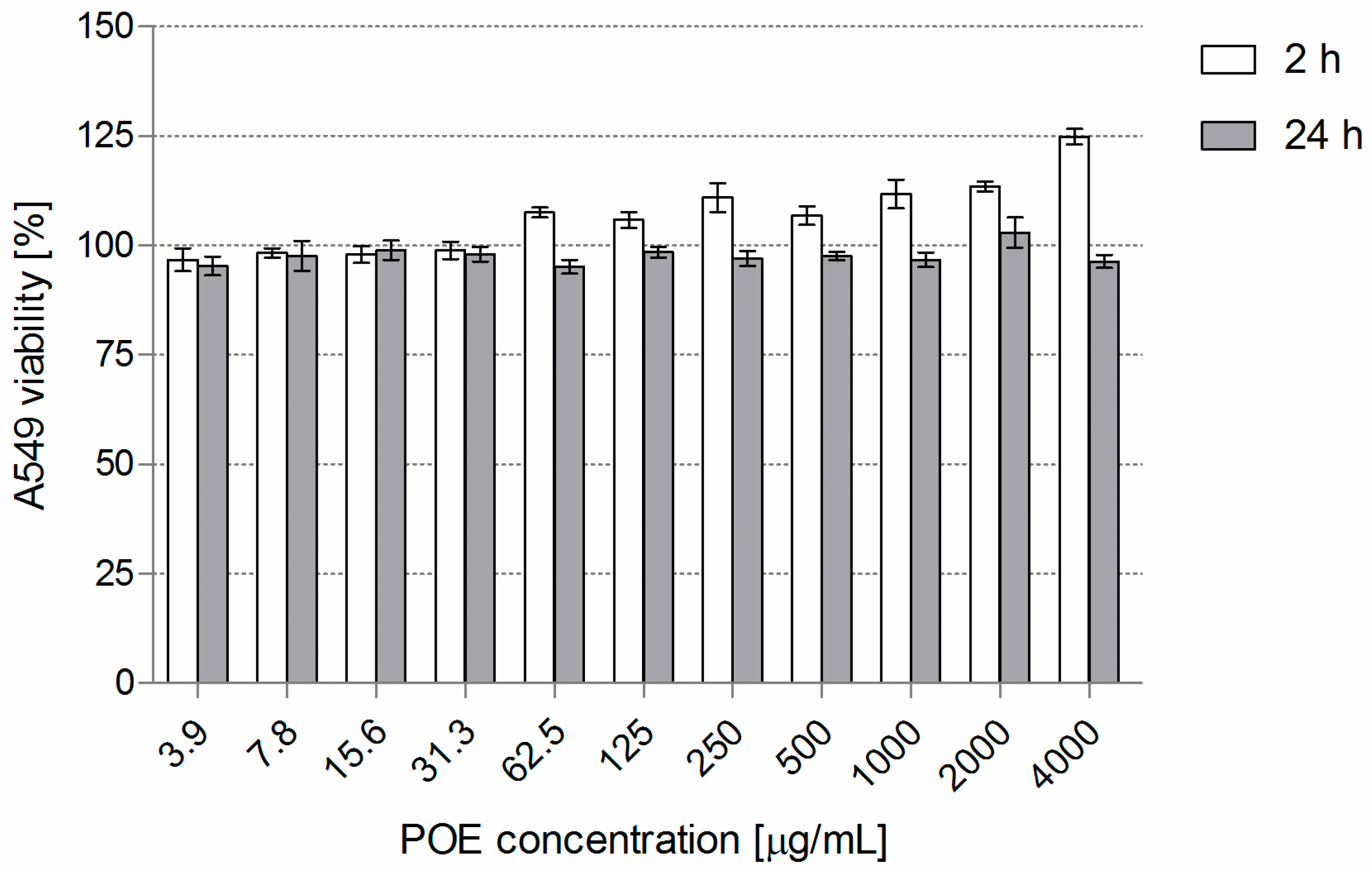
3.2. POE Cytotoxicity
3.3. Bacteria
3.4. POE Minimum Inhibitory Concentration (MIC)
3.5. Impact of POE on S. a. Adherence and Biofilm Formation on Abiotic Surface and Conditioned with Mucin or Elastin
3.6. Anti-Biofilm Potential of POE
3.7. POE Influence on S. a. Protein A (SpA) Surface Expression and α-Hemolysin (Hla) Synthesis
3.8. Effect of POE on Staphylococcal Sortase A (SrtA) Activity
3.9. S. a. Adherence to A549 Cells
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2018, 17, 1–32. [Google Scholar] [CrossRef]
- Heras, B.; Scanlon, M.J.; Martin, J.L. Targeting virulence not viability in the search for future antibacterials. Br. J. Clin. Pharmacol. 2015, 79, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Bennet, B.C. Doctrine of signatures: An explanation of medicinal plant discovery or dissemination of knowledge? Econ. Bot. 2007, 61, 246–255. [Google Scholar] [CrossRef]
- Krzyżanowska-Kowalczyk, J.; Kołodziejczyk-Czepas, J.; Kowalczyk, M.; Pecio, Ł.; Nowak, P.; Stochmal, A. Yunnaneic acid B, a component of Pulmonaria officinalis extract, prevents peroxynitrite-induced oxidative stress in vitro. J. Agric. Food. Chem. 2017, 65, 3827–3834. [Google Scholar] [CrossRef]
- National Database of Health Protection Products. Available online: http://kbpoz.gs1.pl/ (accessed on 12 November 2018).
- European Cystic Fibrosis Society Patient Registry. Annual Data Report (Year 2016) Version 1.2018; European Cystic Fibrosis Society Patient Registry: Karup, Denmark, 2016; pp. 1–141. [Google Scholar]
- Castellani, C.; Assael, B.M. Cystic fibrosis: A clinical view. Cell. Mol. Life Sci. 2017, 74, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Ehre, C.; Ridley, C.; Thornton, D.J. Cystic fibrosis: An inherited disease affecting mucin-producing organs. Int. J. Biochem. Cell Biol. 2014, 52, 136–145. [Google Scholar] [CrossRef]
- Magalhães, A.P.; Lopes, S.P.; Pereira, M.O. Insights into cystic fibrosis polymicrobial consortia: The role of species interactions in biofilm development, phenotype, and response to in-use antibiotics. Front. Microbiol. 2016, 7, 2146. [Google Scholar] [CrossRef] [PubMed]
- Cystic Fibrosis Foundation. CFF Patient Registry Highlights 2016; Cystic Fibrosis Foundation: Bethesda, MD, USA, 2016; pp. 1–4. [Google Scholar]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef]
- Goerke, C.; Wolz, C. Adaptation of Staphylococcus aureus to the cystic fibrosis lung. Int. J. Med. Microbiol. 2010, 300, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, D.; Harper, L.; Shopsin, B.; Torres, V.J. Staphylococcus aureus pathogenesis in diverse host environments. Pathog. Dis. 2017, 75, ftx005. [Google Scholar] [CrossRef] [PubMed]
- Bien, J.; Sokolova, O.; Bozko, P. Characterization of virulence factors of Staphylococcus aureus: Novel function of known virulence factors that are implicated in activation of airway epithelial proinflammatory response. J. Pathog. 2011, 2011, 601905. [Google Scholar] [CrossRef] [PubMed]
- Falugi, F.; Kim, H.K.; Missiakas, D.M.; Schneewind, O. Role of protein A in the evasion of host adaptive immune responses by Staphylococcus aureus. mBio 2013, 4, e00575-13. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Lin, Y.C.; Gillman, A.N.; Parks, P.J.; Schlievert, P.M.; Peterson, M.L. Alpha-toxin promotes Staphylococcus aureus mucosal biofilm formation. Front. Cell Infect. Microbiol. 2012, 2, 64. [Google Scholar] [CrossRef]
- Dey, I.; Shah, K.; Bradbury, N.A. Natural compounds as therapeutic agents in the treatment cystic fibrosis. J. Genet. Syndr. Gene Ther. 2016, 7, 284. [Google Scholar] [CrossRef] [PubMed]
- Krzyżanowska-Kowalczyk, J.; Pecio, Ł.; Mołdoch, J.; Ludwiczuk, A.; Kowalczyk, M. Novel phenolic constituents of Pulmonaria officinalis L. LC-MS/MS comparison of spring and autumn metabolite profiles. Molecules 2018, 23, 2277. [Google Scholar]
- Borges, A.; Abreu, A.C.; Dias, C.; Saavedra, M.J.; Borges, F.; Simões, M. New perspectives on the use of phytochemicals as an emergent strategy to control bacterial infections including biofilms. Molecules 2016, 21, 877. [Google Scholar] [CrossRef] [PubMed]
- Kępa, M.; Miklasińska-Majdanik, M.; Wojtyczka, R.D.; Idzik, D.; Korzeniowski, K.; Smoleń-Dzirba, J.; Wąsik, T.J. Antimicrobial potential of caffeic acid against Staphylococcus aureus clinical strains. BioMed Res. Int. 2018, 2018, 7413504. [Google Scholar] [CrossRef]
- Wu, Y.; Bai, J.; Zhong, K.; Huang, Y.; Qi, H.; Jiang, Y.; Gao, H. Antibacterial activity and membrane-disruptive mechanism of 3-p-trans-coumaroyl-2-hydroxyquinic acid, a novel phenolic compound from pine needles of Cedrus deodara, against Staphylococcus aureus. Molecules 2016, 21, 1084. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Bai, J.; Liu, X.; Liu, L.; Zhong, K.; Huang, Y.; Gao, H. Antibacterial effect of 3-p-trans-coumaroyl-2-hydroxyquinic acid, a phenolic compound from needles of Cedrus deodara, on cellular functions of Staphylococcus aureus. RSC Adv. 2018, 8, 4969–4975. [Google Scholar] [CrossRef]
- Slobodníková, L.; Fialová, S.; Hupková, H.; Grancai, D. Rosmarinic acid interaction with planktonic and biofilm Staphylococcus aureus. Nat. Prod. Commun. 2013, 8, 1747–1750. [Google Scholar] [PubMed]
- Ekambaram, S.P.; Perumal, S.S.; Balakrishnan, A.; Marappan, N.; Gajendran, S.S.; Viswanathan, V. Antibacterial synergy between rosmarinic acid and antibiotics against methicillin-resistant Staphylococcus aureus. J. Intercult. Ethnopharmacol. 2016, 5, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, J.H.; Cho, H.S.; Joo, S.W.; Cho, M.H.; Lee, J. Anti-biofilm activities of quercetin and tannic acid against Staphylococcus aureus. Biofouling 2013, 29, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Ming, D.; Wang, D.; Cao, F.; Xiang, H.; Mu, D.; Cao, J.; Li, B.; Zhong, L.; Dong, X.; Zhong, X.; et al. Kaempferol inhibits the primary attachement phase of biofilm formation in Staphylococcus aureus. Front. Microbiol. 2017, 8, 2263. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.H.; Kim, S.B.; Kong, R.; Choi, J.G.; Kim, Y.C.; Shin, D.W.; Kang, O.H.; Kwon, D.Y. Curcumin reverse methicillin resistance in Staphylococcus aureus. Molecules 2014, 19, 18283–18295. [Google Scholar] [CrossRef] [PubMed]
- Sardi, J.C.O.; Polaquini, C.R.; Freires, I.A.; Galvão, L.C.C.; Lazarini, J.G.; Torrezan, G.S.; Regasini, L.O.; Rosalen, P.L. Antibacterial activity of diacetylcurcumin against Staphylococcus aureus results in decreased biofilm and cellular adhesion. J. Med. Microbiol. 2017, 66, 816–824. [Google Scholar] [CrossRef]
- Teow, S.Y.; Liew, K.; Ali, S.A.; Khoo, A.S.B.; Peh, S.C. Antibacterial action of curcumin against Staphylococcus aureus: A brief review. J. Trop. Med. 2016, 2016, 2853045. [Google Scholar] [CrossRef] [PubMed]
- Slobodníková, L.; Fialová, S.; Rendeková, K.; Kováč, J.; Mučaji, P. Antibiofilm activity of plant polyphenols. Molecules 2016, 21, 1717. [Google Scholar]
- Gomez-Fernandes, M.; Laabei, M.; Pagan, N.; Hidalgo, J.; Molinos, S.; Hernandez, R.V.; Domínguez-Villanueva, D.; Jenkins, A.T.A.; Lacoma, A.; Prat, C. Accessory gene regulator (Agr) functionality in Staphylococcus aureus derived from lower respiratory tract infections. PLoS ONE 2017, 12, e0175552. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, Y.; Riehle, A.; Ma, J.; Kamler, M.; Gulbins, E.; Grassmé, H. Staphylococcus aureus survives in cystic fibrosis macrophages, forming a reservoir for chronic pneumonia. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.; Martins, N.; Barros, L.; Rodrigues, M.E.; Oliveira, M.B.P.P.; Henriques, M.; Ferreira, I.C. Plant phenolic extracts as an effective strategy to control Staphylococcus aureus, the dairy industry pathogen. Ind. Crops Prod. 2018, 112, 515–520. [Google Scholar] [CrossRef]
- Hodille, E.; Rose, W.; Diep, B.A.; Goutelle, S.; Lina, G.; Dumitrescu, O. The role of antibiotics in modulating virulence in Staphylococcus aureus. Clin. Microbiol. Rev. 2017, 30, 887–917. [Google Scholar] [CrossRef]
- Wang, L.; Bi, C.; Cai, H.; Liu, B.; Zhong, X.; Deng, X.; Wang, T.; Xiang, H.; Niu, X.; Wang, D. The therapeutic effect of chlorogenic acid against Staphylococcus aureus infection through sortase A inhibition. Front. Microbiol. 2015, 6, 1031. [Google Scholar] [CrossRef] [PubMed]
- Cascioferro, S.; Raffa, D.; Maggio, B.; Raimondi, M.V.; Schillaci, D.; Daidone, G. Sortase A inhibitors: Recent advances and future perspectives. J. Med. Chem. 2015, 58, 9108–9123. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, M.; Giannone, R.J.; Bemis, D.A.; Kania, S.A. Molecular basis of surface anchored protein A deficiency in the Staphylococcus aureus strain Wood 46. PLoS ONE 2017, 12, e0183913. [Google Scholar] [CrossRef] [PubMed]
- Różalska, B.; Sadowska, B.; Żuchowski, J.; Więckowska-Szakiel, M.; Budzyńska, A.; Wójcik, U.; Stochmal, A. Phenolic and nonpolar fractions of Elaeagnus rhamnoides (L.) A. Nelson extracts as virulence modulators—in vitro study on bacteria, fungi, and epithelial cells. Molecules 2018, 23, 1498. [Google Scholar] [CrossRef]
- Sadowska, B.; Bonar, A.; von Eiff, C.; Proctor, R.A.; Chmiela, M.; Rudnicka, W.; Róźalska, B. Characteristics of Staphylococcus aureus, isolated from airways of cystic fibrosis patients, and their small colony variants. FEMS Immunol. Med. Microbiol. 2002, 32, 191–197. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing—EUCAST. 2012. Available online: http://www.eucast.org (accessed on 5 June 2018).
- Micota, B.; Sadowska, B.; Podsędek, A.; Paszkiewicz, M.; Sosnowska, D.; Różalska, B. Is it true that plant-derived polyphenols are always beneficial for the human? In vitro study on Leonurus cardiaca extract properties in the context of the pathogenesis of Staphylococcus aureus infections. J. Med. Microbiol. 2016, 65, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. aureus Strain | Hla ± S.D. [ng/mL] | ||
|---|---|---|---|
| Control (Untreated) | POE 0.5 × MIC | POE 0.25 × MIC | |
| ATCC 29213 | 1035.0 ± 12.0 | 520.5 ± 60.7 * | 632.6 ± 80.6 * |
| ATCC 700699 | 103.9 ± 8.9 | 60.7 ± 9.1 * | 76.5 ± 12.1 * |
| α4 | 905.0 ± 70.5 | 839.4 ± 39.9 | 901.1 ± 18.1 |
| α28 | 736.7 ± 12.1 | 426.1 ± 21.3 * | 529.8 ± 40.8 * |
| γ14 | 1053.0 ± 181.9 | 526.5 ± 53.7 * | 518.7 ± 48.2 * |
| γ18 | 109.6 ± 1.1 | 44.9 ± 6.3 * | 93.9 ± 4.4 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadowska, B.; Wójcik, U.; Krzyżanowska-Kowalczyk, J.; Kowalczyk, M.; Stochmal, A.; Rywaniak, J.; Burzyńska, J.; Różalska, B. The Pros and Cons of Cystic Fibrosis (CF) Patient Use of Herbal Supplements Containing Pulmonaria officinalis L. Extract: the Evidence from an In Vitro Study on Staphylococcus aureus CF Clinical Isolates. Molecules 2019, 24, 1151. https://doi.org/10.3390/molecules24061151
Sadowska B, Wójcik U, Krzyżanowska-Kowalczyk J, Kowalczyk M, Stochmal A, Rywaniak J, Burzyńska J, Różalska B. The Pros and Cons of Cystic Fibrosis (CF) Patient Use of Herbal Supplements Containing Pulmonaria officinalis L. Extract: the Evidence from an In Vitro Study on Staphylococcus aureus CF Clinical Isolates. Molecules. 2019; 24(6):1151. https://doi.org/10.3390/molecules24061151
Chicago/Turabian StyleSadowska, Beata, Urszula Wójcik, Justyna Krzyżanowska-Kowalczyk, Mariusz Kowalczyk, Anna Stochmal, Joanna Rywaniak, Julia Burzyńska, and Barbara Różalska. 2019. "The Pros and Cons of Cystic Fibrosis (CF) Patient Use of Herbal Supplements Containing Pulmonaria officinalis L. Extract: the Evidence from an In Vitro Study on Staphylococcus aureus CF Clinical Isolates" Molecules 24, no. 6: 1151. https://doi.org/10.3390/molecules24061151
APA StyleSadowska, B., Wójcik, U., Krzyżanowska-Kowalczyk, J., Kowalczyk, M., Stochmal, A., Rywaniak, J., Burzyńska, J., & Różalska, B. (2019). The Pros and Cons of Cystic Fibrosis (CF) Patient Use of Herbal Supplements Containing Pulmonaria officinalis L. Extract: the Evidence from an In Vitro Study on Staphylococcus aureus CF Clinical Isolates. Molecules, 24(6), 1151. https://doi.org/10.3390/molecules24061151