
Radicinin, a Fungal Phytotoxin as a Target-Specific Bioherbicide for Invasive Buffelgrass (Cenchrus ciliaris) Control

 ,
,  ,
,  ,
,  ,
, 
Abstract
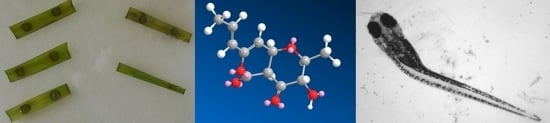
:
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Fungal Strains
3.3. Isolation of Fungal Metabolites
3.4. Leaf Puncture Bioassays
3.5. Statistical Analyses
3.6. Fish Embryo Toxicity (FET) Test
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abella, S.R.; Chiquoine, L.P.; Backer, D.M. Ecological characteristics of sites invaded by buffelgrass (Pennisetum ciliare). Invasive Plant Sci. Manag. 2012, 5, 443–453. [Google Scholar] [CrossRef]
- Marshall, V.M.; Lewis, M.M.; Ostendorf, B. Buffelgrass (Cenchrus ciliaris) as an invader and threat to biodiversity in arid environments: A review. J. Arid. Environ. 2012, 78, 1–12. [Google Scholar] [CrossRef]
- De La Barrera, E.; Castellanos, A.E. High temperature effects on gas exchange for the invasive buffelgrass (Pennisetum ciliare L.). Weed Biol. Manag. 2007, 7, 128–131. [Google Scholar] [CrossRef]
- Burquez-Montijo, A.M.; Miller, M.E.; Yrizar, A.M.; Tellman, B. Mexican grasslands, Thornscrub, and the transformation of the Sonoran Desert by invasive exotic buffelgrass (Pennisetum ciliare). In Invasive Exotic Species in the Sonoran Region; Tellman, B., Ed.; The University of Arizona Press, The Arizona-Sonora Desert Museum: Tucson, AZ, USA, 2002; pp. 126–146. [Google Scholar]
- Stevens, J.; Falk, D.A. Can buffelgrass invasions be controlled in the American Southwest? Using invasion ecology theory to understand buffelgrass success and develop comprehensive restoration and management. Ecol. Restor. 2009, 27, 417–427. [Google Scholar] [CrossRef]
- Olsson, A.D.; Betancourt, J.L.; Crimmins, M.A.; Marsh, S.E. Constancy of local spread rates for buffelgrass (Pennisetum ciliare L.) in the Arizona Upland of the Sonoran Desert. J. Arid. Environ. 2012, 87, 136–143. [Google Scholar] [CrossRef]
- Rogstad, A. The Buffelgrass Strategic Plan; Rogstad, A., Ed.; The University of Arizona Press, The Arizona-Sonora Desert Museum: Tucson, AZ, USA, 2008. [Google Scholar]
- Bean, T.M. Tools for Improved Management of Buffelgrass in the Sonoran Desert. Ph.D. Dissertation, The University of Arizona, Phoenix, AZ, USA, 2014. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Handbook of Sustainable Weed Management; Singh, H.P., Batish, D.R., Kohli, R.K., Eds.; The Haworth Press Inc.: New York, NY, USA, 2006. [Google Scholar]
- Dayan, F.E.; Duke, S.O. Natural compounds as next generation herbicides. Plant Physiol. 2014, 166, 1090. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, B.C.; Sparks, T.C. Natural products for pest control: An analysis of their role, value and future. Pest. Man. Sci. 2014, 70, 1169–1185. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef] [PubMed]
- Harding, D.P.; Raizada, M.N. Controlling weeds with fungi, bacteria and viruses: A review. Front. Plant. Sci. 2015, 6, 659. [Google Scholar] [CrossRef] [PubMed]
- Cordeau, S.; Triolet, M.; Wayman, S.; Steinberg, C.; Guillemin, J.P. Bioherbicides: Dead in the water? A review of the existing products for integrated weed management. Crop. Prot. 2016, 87, 44–49. [Google Scholar] [CrossRef]
- Charudattan, R. Biological control of weeds by means of plant pathogens: Significance for integrated weed management in modern agro-ecology. BioControl 2001, 46, 229–260. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Alqarawi, A.A.; AbdAllah, E.F. Bioherbicides: Current knowledge on weed control mechanism. Ecotoxicol. Environ. Saf. 2018, 158, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Bailey, K.L. The bioherbicide approach to weed control using plant pathogens. In Integrated Pest Management: Current Concepts and Ecological Perspective; Abrol, D.P., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 245–266. [Google Scholar]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Meyer, S.; Clement, S.; Cimmino, A.; Cristofaro, M.; Evidente, A. Cochliotoxin, a dihydropyranopyran-4,5-dione, and its analogues produced by Cochliobolus australiensis display phytotoxic activity against buffelgrass (Cenchrus ciliaris). J. Nat. Prod. 2017, 80, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Meyer, S.; Clement, S.; Pescitelli, G.; Cimmino, A.; Cristofaro, M.; Evidente, A. Chloromonilinic acids C and D, two phytotoxic tetrasubstituted 3-chromanonacrylic acids isolated from Cochliobolus australiensis with potential herbicidal activity against buffelgrass (Cenchrus ciliaris). J. Nat. Prod. 2017, 80, 2771–2777. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Meyer, S.; Górecki, M.; Mandoli, A.; Di Bari, L.; Pescitelli, G.; Cimmino, A.; Cristofaro, A.; Clement, S.; Evidente, A. Pyriculins A and B, two monosubstituted hex-4-ene-2,3-diols and other phytotoxic metabolites produced by Pyricularia grisea isolated from buffelgrass (Cenchrus ciliaris). Chirality 2017, 29, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, T.J.; Rolshausen, P.E.; Roper, M.C.; Reader, J.M.; Steinhaus, M.J.; Rapicavoli, J.; Vosburg, D.A.; Maloney, K.N. Radicinin from Cochliobolus sp. inhibits Xylella fastidiosa, the causal agent of Pierce’s Disease of grapevine. Phytochemistry 2015, 116, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Robeson, D.J.; Gray, G.R.; Strobel, G.A. Production of the phytotoxins radicinin and radicinol by Alternaria chrysanthemi. Phytochemistry 1982, 21, 2359–2362. [Google Scholar] [CrossRef]
- Lahnsteiner, F. The sensitivity and reproducibility of the zebrafish (Danio rerio) embryo test for the screening of waste water quality and for testing the toxicity of chemicals. Altern. Lab. Anim. 2008, 36, 299–311. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds 1–14 are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
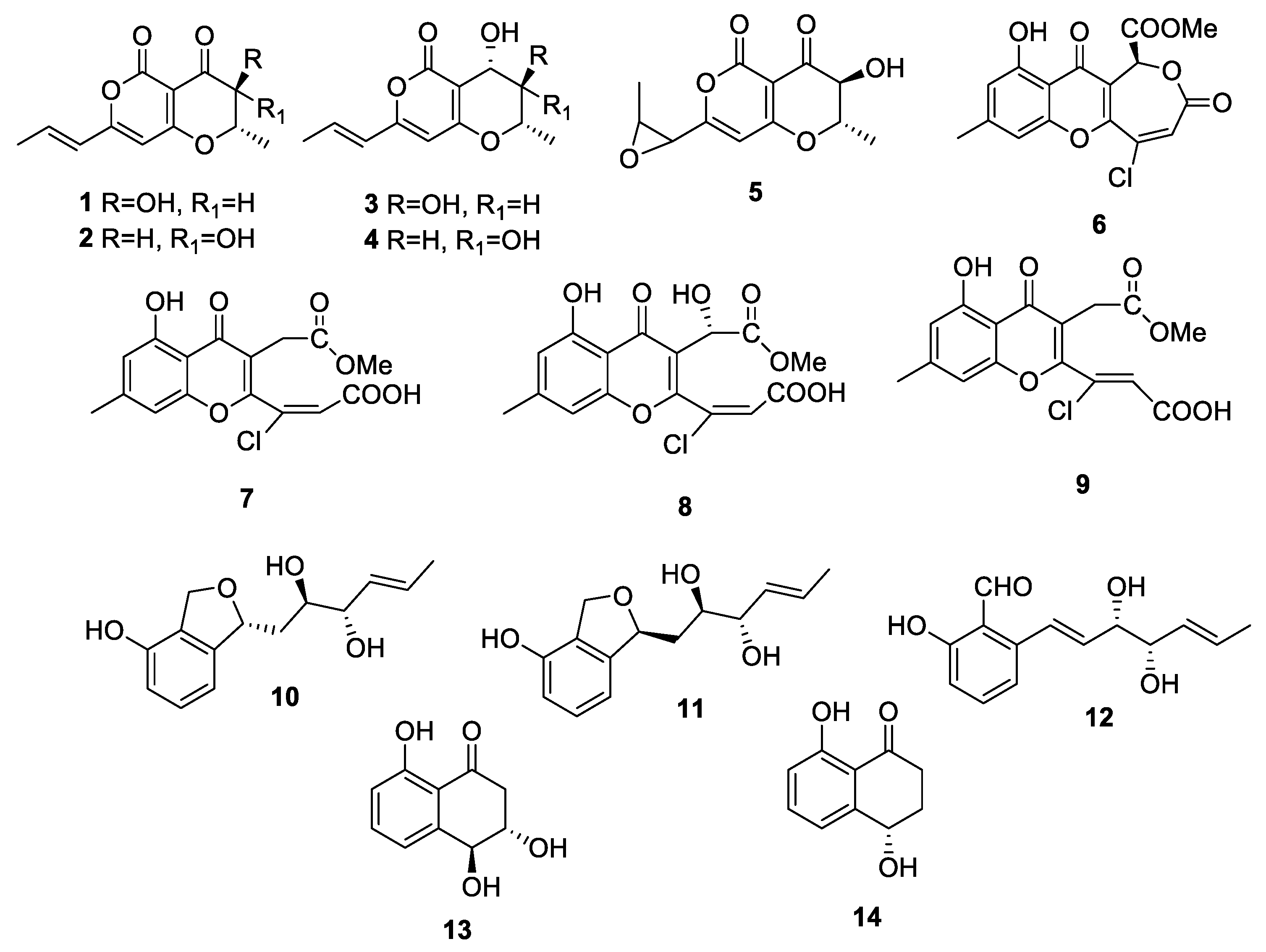
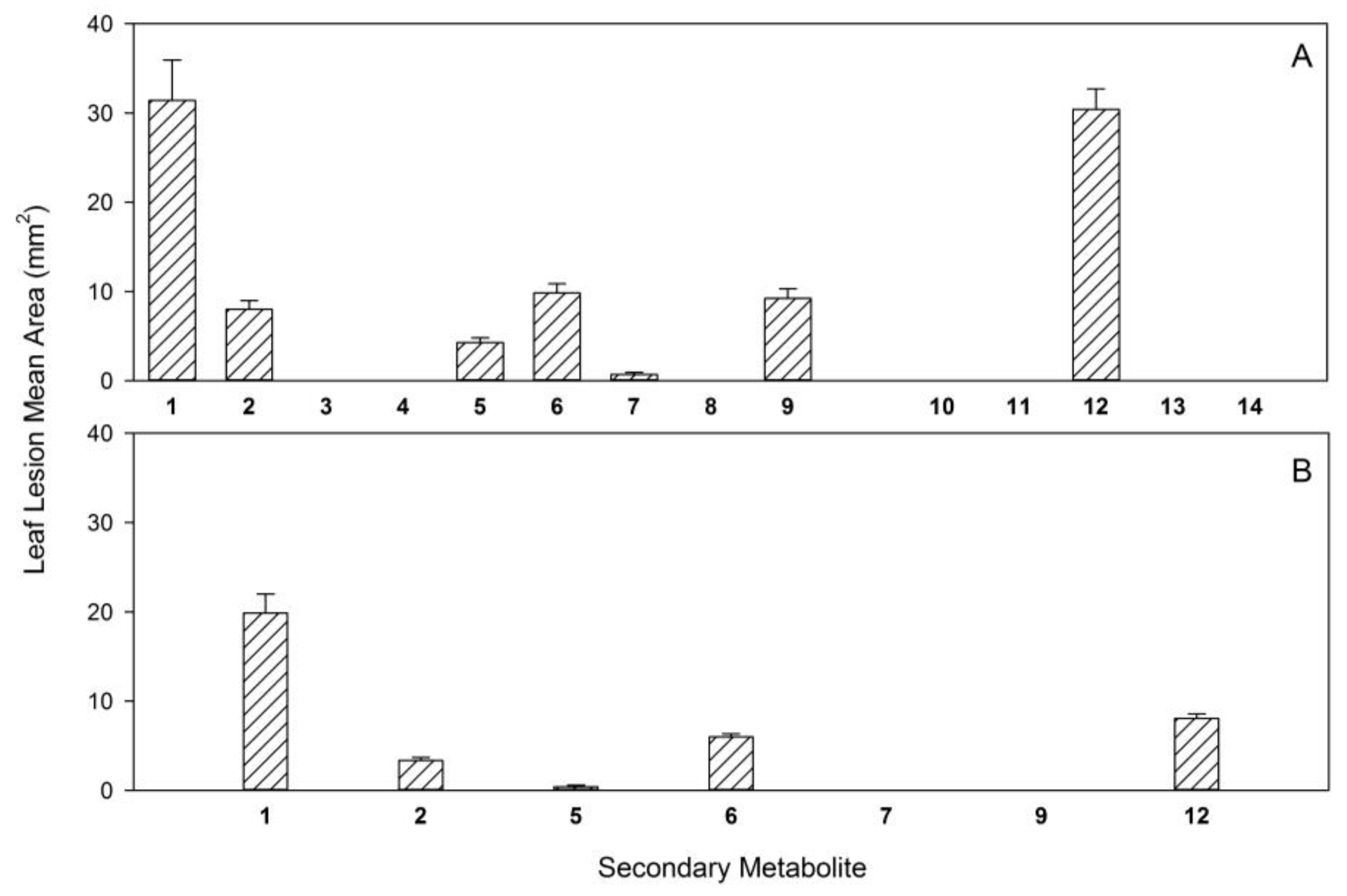
| Compound | Seedling Bioassay Toxicity 1 | Reference |
|---|---|---|
| From Cochliobolus australiensis | ||
| radicinin (1) | high | [19] |
| 3-epi-radicinin (2) | high | [19] |
| radicinol (3) | nil | [19] |
| 3-epi-radicinol (4) | nil | [19] |
| cochliotoxin (5) | high | [19] |
| chloromonilicin (6) | not tested | [20] |
| chloromonilinic acid B (7) | high | [20] |
| chloromonilinic acid C (8) | high | [20] |
| chloromonilinic acid D (9) | high | [20] |
| From Pyricularia grisea | ||
| pyriculin A (10) | low | [21] |
| pyriculin B (11) | nil | [21] |
| (10S, 11S)-(−)-epi-pyriculol (12) | high | [21] |
| trans-3,4-dihydro-3,4,8-trihydroxy-1(2H)-naphtalenone (13) | nil | [21] |
| (4S)-(+)-isosclerone (14) | nil | [21] |
| Species | Family | Common Name | Growth Form | Primary Geographic Distribution |
|---|---|---|---|---|
| Digitaria californica | Poaceae | Arizona Cottontop | C4 perennial grass | Sonoran and Chihuahuan Deserts |
| Heteropogon contortus | Poaceae | Tanglehead | C4 perennial grass | Sonoran and Chihuahuan Deserts |
| Baileya multiradiata | Asteraceae | Desert Marigold | Perennial dicot | Mojave, Sonoran and Chihuahuan Deserts |
| Stanleya pinnata | Brassicaceae | Prince’s Plume | Perennial dicot | Western North America (widespread) |
| Encelia frutescens | Asteraceae | Brittlebush | Shrub | Mojave, Sonoran Deserts |
| Lepidium fremontii | Brassicaceae | Fremont Pepperbush | Shrub | Mojave Desert |
| Type 3 Tests of Fixed Effects | ||||
|---|---|---|---|---|
| Effect | Num DF 1 | Den DF 2 | F Value 3 | p-Value 4 |
| species | 6 | 498 | 150.49 | <0001 |
| toxin | 1 | 498 | 0.48 | 0.4876 |
| species × toxin | 6 | 498 | 16.74 | <0001 |
| concentration | 1 | 498 | 295.74 | <0001 |
| species × concentration | 6 | 498 | 17.84 | <0001 |
| toxin × concentration | 1 | 498 | 0.00 | 0.9616 |
| species × toxin × concentration | 6 | 498 | 11.68 | <0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masi, M.; Freda, F.; Sangermano, F.; Calabrò, V.; Cimmino, A.; Cristofaro, M.; Meyer, S.; Evidente, A. Radicinin, a Fungal Phytotoxin as a Target-Specific Bioherbicide for Invasive Buffelgrass (Cenchrus ciliaris) Control. Molecules 2019, 24, 1086. https://doi.org/10.3390/molecules24061086
Masi M, Freda F, Sangermano F, Calabrò V, Cimmino A, Cristofaro M, Meyer S, Evidente A. Radicinin, a Fungal Phytotoxin as a Target-Specific Bioherbicide for Invasive Buffelgrass (Cenchrus ciliaris) Control. Molecules. 2019; 24(6):1086. https://doi.org/10.3390/molecules24061086
Chicago/Turabian StyleMasi, Marco, Fabrizio Freda, Felicia Sangermano, Viola Calabrò, Alessio Cimmino, Massimo Cristofaro, Susan Meyer, and Antonio Evidente. 2019. "Radicinin, a Fungal Phytotoxin as a Target-Specific Bioherbicide for Invasive Buffelgrass (Cenchrus ciliaris) Control" Molecules 24, no. 6: 1086. https://doi.org/10.3390/molecules24061086
APA StyleMasi, M., Freda, F., Sangermano, F., Calabrò, V., Cimmino, A., Cristofaro, M., Meyer, S., & Evidente, A. (2019). Radicinin, a Fungal Phytotoxin as a Target-Specific Bioherbicide for Invasive Buffelgrass (Cenchrus ciliaris) Control. Molecules, 24(6), 1086. https://doi.org/10.3390/molecules24061086