
Protective Effects of Kuding Tea (Ilex kudingcha C. J. Tseng) Polyphenols on UVB-Induced Skin Aging in SKH1 Hairless Mice


Abstract
1. Introduction
2. Results
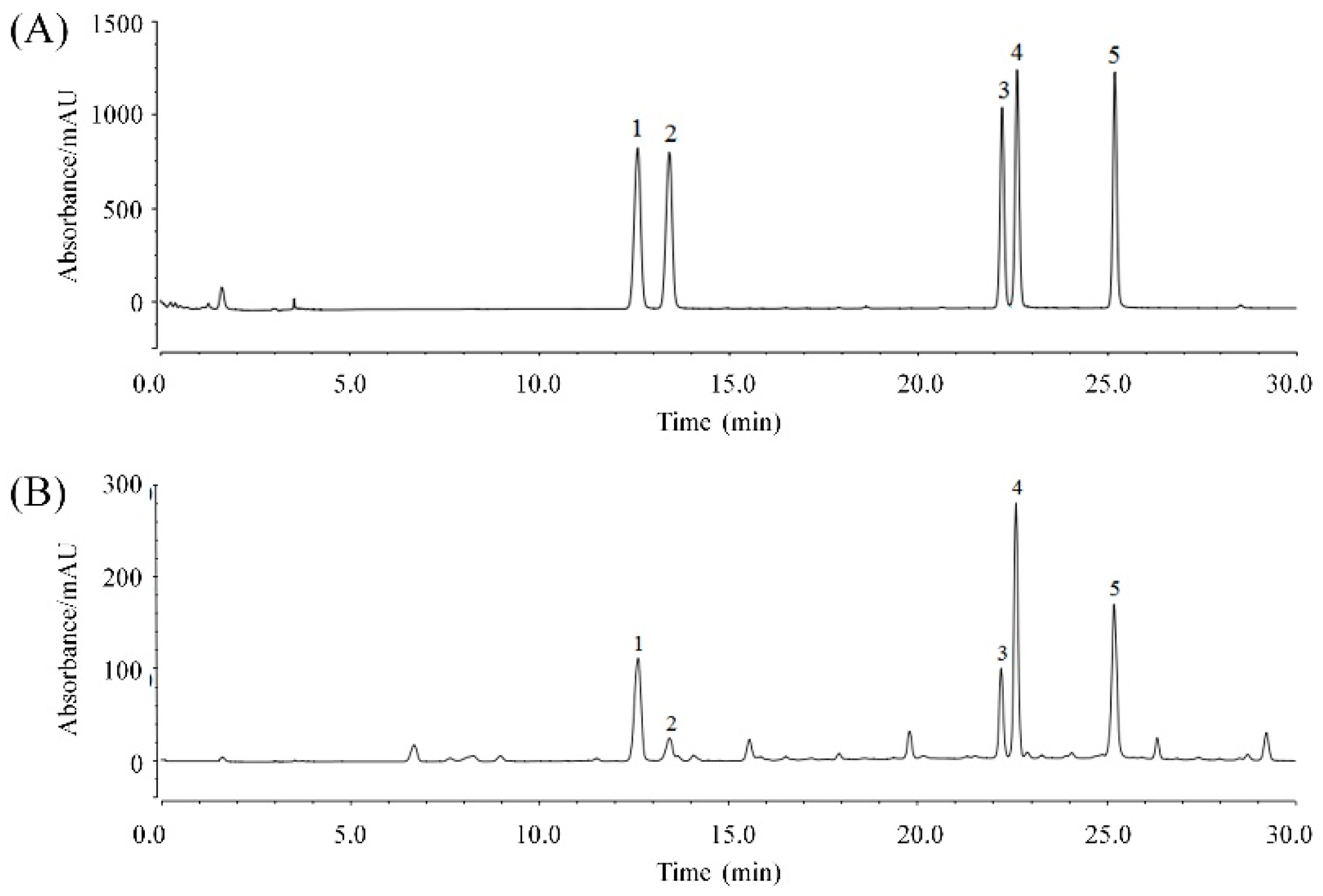
2.1. KTP Constituents
2.2. Serum Levels of T-SOD, CAT, and MDA
2.3. Serum Levels of IL-6, IL-1β, and TNF-α
2.4. Col I, Col III, Hydroxyproline, and Hyaluronic Acid Contents in Skin Tissue
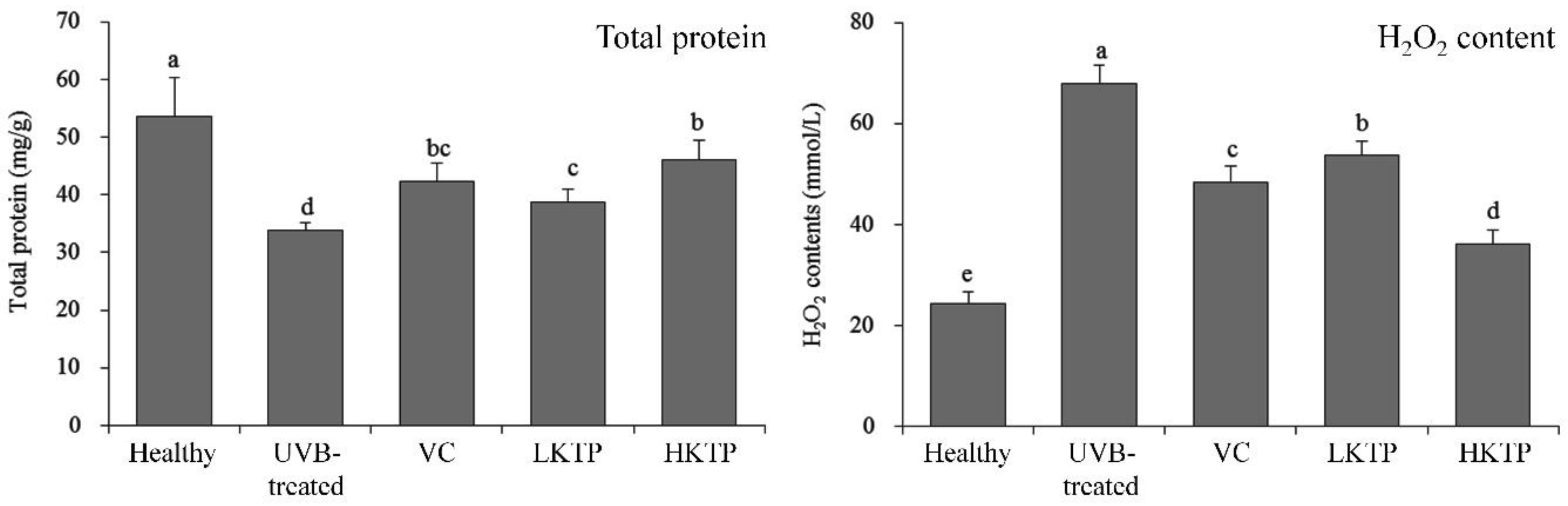
2.5. Total Protein and H2O2 of Skin Tissue
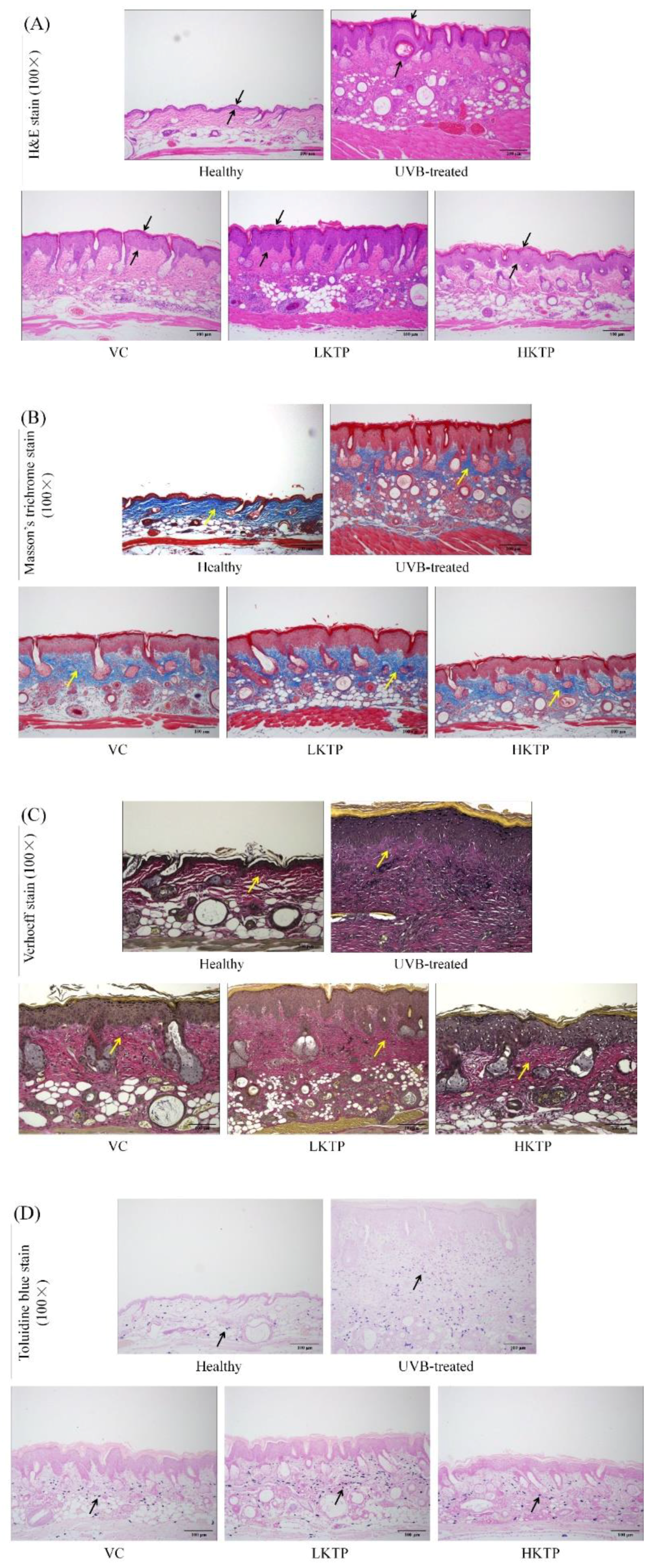
2.6. Histopathological Observations of Skin Tissue
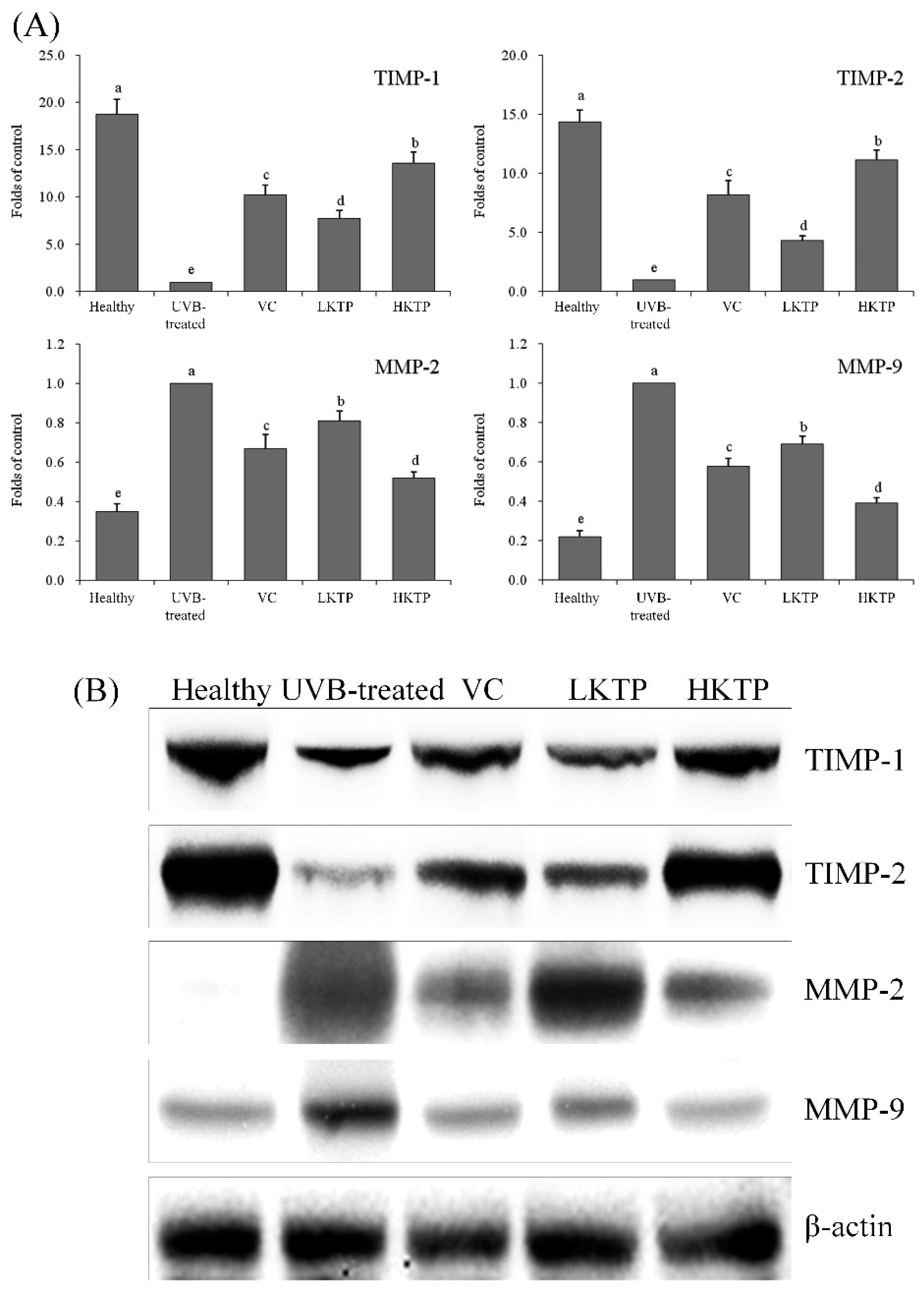
2.7. mRNA and Protein Expression of TIMP-1, TIMP-2, MMP-2, and MMP-9 in Skin Tissue
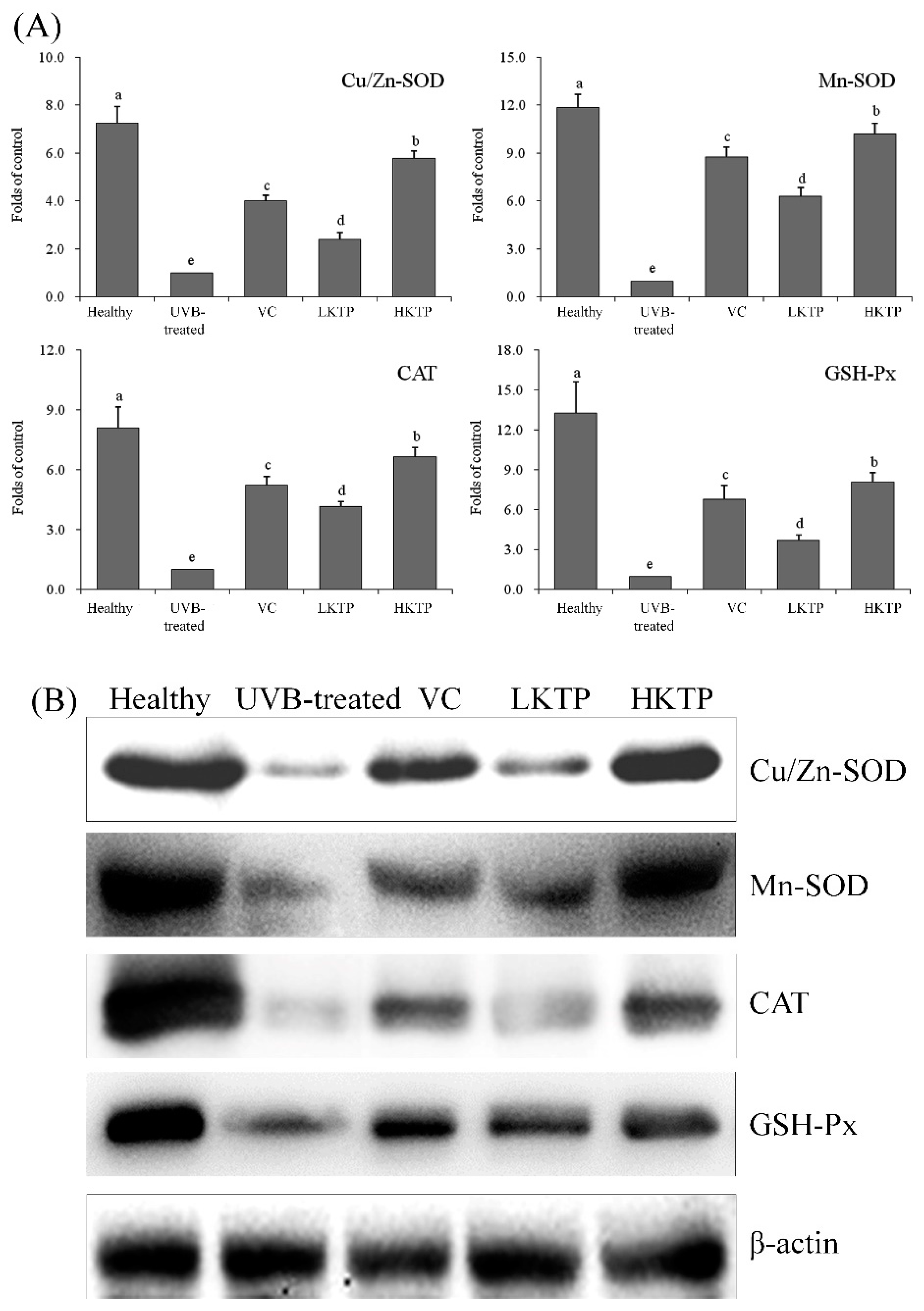
2.8. mRNA and Protein Expression of Cu/Zn-SOD, Mn-SOD, CAT, and GSH-Px in Skin Tissue
3. Discussion
4. Materials and Methods
4.1. KTP Extraction
4.2. High-Performance Liquid Chromatography
4.3. Skin Injury Induced by UV Radiation
4.4. Determination of Oxidation-Related Serum Levels
4.5. Determination of Serum Cytokine Levels
4.6. Pathological Observations of Skin Tissue
4.7. Determination of Protein Levels in Skin Tissue
4.8. Quantitative Polymerase Chain Reaction
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Surjadinata, B.B.; Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. UVA, UVB and UVC light enhances the biosynthesis of phenolic antioxidants in fresh-cut carrot through a synergistic effect with wounding. Molecules 2017, 22, 668. [Google Scholar] [CrossRef]
- Sayre, R.M.; Dowdy, J.C.; Shepherd, J.G. Reintroduction of a classic vitamin D ultraviolet source. J. Steroid Biochem. Mol. Biol. 2007, 103, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Schlereth, T.; Schönefeld, S.; Birklein, F.; Kirkpatrick, C.J.; Wessler, I. In vivo release of non-neuronal acetylcholine from human skin by dermal microdialysis: Effects of sunlight, UV-A and tactile stimulus. Life Sci. 2007, 80, 2239–2242. [Google Scholar] [CrossRef] [PubMed]
- Miyamae, Y.; Yamakawa, Y.; Kawabata, M.; Ozaki, Y. A combined near-infrared diffuse reflectance spectroscopy and principal component analysis method of assessment for the degree of photoaging and physiological aging of human skin. Anal. Sci. 2012, 28, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- De Fabo, E.C. Arctic stratospheric ozone depletion and increased UVB radiation: Potential impacts to human health. Int. J. Circumpolar Health 2005, 64, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Ichihashi, M.; Ando, H. The maximal cumulative solar UVB dose allowed to maintain healthy and young skin and prevent premature photoaging. Exp. Dermatol. 2014, 2014, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Rijken, F.; Bruijnzeel-Koomen, C.A. Photoaged skin: The role of neutrophils, preventive measures, and potential pharmacological targets. Clin. Pharmacol. Ther. 2011, 89, 120–124. [Google Scholar] [CrossRef]
- Wulf, H.C.; Sandby-Møller, J.; Kobayasi, T.; Gniadecki, R. Skin aging and natural photoprotection. Micron 2004, 35, 185–191. [Google Scholar] [CrossRef]
- Godos, J.; Marventano, S.; Mistretta, A.; Galvano, F.; Grosso, G. Dietary sources of polyphenols in the mediterranean healthy eating, aging and lifestyle (MEAL) study cohort. Int. J. Food Sci. Nutr. 2017, 68, 750–756. [Google Scholar] [CrossRef]
- Colomer, R.; Sarrats, A.; Lupu, R.; Puig, T. Natural polyphenols and their synthetic analogs as emerging anticancer agents. Curr. Drug Targets. 2017, 18, 147–159. [Google Scholar] [CrossRef]
- Venables, M.C.; Hulston, C.J.; Cox, H.R.; Jeukendrup, A.E. Green tea extract ingestion, fat oxidation, and glucose tolerance in healthy humans. Am. J. Clin. Nutr. 2008, 87, 778–784. [Google Scholar] [CrossRef]
- Sur, S.; Panda, C.K. Molecular aspects of cancer chemopreventive and therapeutic efficacies of tea and tea polyphenols. Nutrition 2017, 43–44, 8–15. [Google Scholar] [CrossRef]
- Afanas’ev, I.B. Signaling by reactive oxygen and nitrogen species in skin diseases. Curr. Drug Metab. 2010, 11, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Challa, A.; McCormick, T.S.; Cooper, K.D.; Mukhtar, H. Prevention of UVB-induced immunosuppression in mice by the green tea polyphenol (-)-epigallocatechin-3-gallate may be associated with alterations in IL-10 and IL-12 production. Carcinogenesis 1999, 20, 2117–2124. [Google Scholar] [CrossRef] [PubMed]
- Roh, E.; Kim, J.E.; Kwon, J.Y.; Park, J.S.; Bode, A.M.; Dong, Z.; Lee, K.W. Molecular mechanisms of green tea polyphenols with protective effects against skin photoaging. Crit. Rev. Food Sci. Nutr. 2017, 57, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Rahman, A.; Hasnain, A.; Lalonde, M.; Goldberg, V.M.; Haqqi, T.M. Green tea polyphenol epigallocatechin-3-gallate inhibits the IL-1 beta-induced activity and expression of cyclooxygenase-2 and nitric oxide synthase-2 in human chondrocytes. Free Radic. Biol. Med. 2002, 33, 1097–1105. [Google Scholar] [CrossRef]
- Wang, S.; Li, H. Radical scavenging activity of ribonuclease inhibitor from cow placenta. Biochemistry 2006, 71, 520–524. [Google Scholar] [CrossRef]
- Roginsky, V.; Alegria, A.E. Oxidation of tea extracts and tea catechins by molecular oxygen. J. Agric. Food Chem. 2005, 53, 4529–4535. [Google Scholar] [CrossRef] [PubMed]
- Sevin, A.; Oztaş, P.; Senen, D.; Han, U.; Karaman, C.; Tarimci, N.; Kartal, M.; Erdoğan, B. Effects of polyphenols on skin damage due to ultraviolet A rays: An experimental study on rats. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhang, Y.; Hu, N.; Sun, Q.; Ding, X.; Li, G.; Zheng, B.; Gu, M.; Huang, F.; Sun, Y.Q.; et al. Extract of Kuding tea prevents high-fat diet-induced metabolic disorders in C57BL/6 mice via liver X receptor (LXR) β antagonism. PLoS ONE 2012, 7, e51007. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, Q. Antioxidant, antimutagenic and in vitro anticancer effects of crude polyphenols in Insect tea made from Kuding tea leaves. Mod. Food Sci. Technol. 2015, 31, 24–29. [Google Scholar]
- Pullar, J.M.; Carr, A.C.; Vissers, M.C.M. The roles of vitamin C in skin health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.L.; Feng, X.; Zhu, K.; Zhao, X. Apoptosis inducing effects of crude polyphenols of Insect tea made by Kuding tea leaves in HepG2 human hepatoma cells. Food Technol. Food Ind. 2015, 36, 339–342. [Google Scholar]
- Zhang, Y.; Zhang, X.; Zhang, W.Q.; Zeng, X.X.; Hu, Q.H. Purification and Structural Identification of Polyphenols in Kudingcha. Food Sci. 2011, 32, 60–63. [Google Scholar]
- Park, S.H.; Cho, D.M.; Choi, B.D.; Choi, Y.J.; Choi, J.H. Antioxidative effects of skinned mugwort (Artemisia vulgaris L.) extracts on UV-irradiated hairless mouse skin. J. Korean Soc. Food Sci. Nutr. 2008, 37, 20–26. [Google Scholar] [CrossRef]
- Suo, H.; Zhao, X.; Qian, Y.; Sun, P.; Zhu, K.; Li, J.; Sun, B. Lactobacillus fermentum Suo attenuates HCl/ethanol induced gastric injury in mice through its antioxidant effects. Nutrients 2016, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Pang, L.; Li, J.; Song, J.L.; Qiu, L.H. Apoptosis inducing effects of Kuding tea polyphenols in human buccal squamous cell carcinoma cell line BcaCD885. Nutrients 2014, 6, 3084–3100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lu, C.; Yu, Z.; Wang, X.; Yan, L.; Zhang, J.; Li, H.; Wang, J.; Wen, A. Echinacoside alleviates UVB irradiation-mediated skin damage via inhibition of oxidative stress, DNA damage, and apoptosis. Oxid. Med. Cell Longev. 2017, 2017, 6851464. [Google Scholar] [CrossRef]
- Xian, D.; Gao, X.; Xiong, X.; Xu, J.; Yang, L.; Pan, L.; Zhong, J. Photoprotection against UV-induced damage by skin-derived precursors in hairless mice. J. Photochem. Photobiol. B 2017, 175, 73–82. [Google Scholar] [CrossRef]
- Zhang, S.; Dong, Z.; Peng, Z.; Lu, F. Anti-aging effect of adipose-derived stem cells in a mouse model of skin aging induced by D-galactose. PLoS ONE 2014, 9, e97573. [Google Scholar] [CrossRef]
- Heydari, S.; Ghanbarzadeh, S.; Anoush, B.; Ranjkesh, M.; Javadzadeh, Y.; Kouhsoltani, M.; Hamishehkar, H. Nanoethosomal formulation of gammaoryzanol for skin-aging protection and wrinkle improvement: A histopathological study. Drug Dev. Ind. Pharm. 2017, 43, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Baba, A.; Kawakami, Y.; Saito, K.; Murashima, Y.L.; Itoh, Y. Effects of edaravone on hippocampal antioxidants in EL mice. J. Nippon Med. Sch. 2016, 83, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Sunanaga, T. Senescence-associated superoxide dismutase influences mitochondrial gene expression in budding tunicates. Dev. Growth Differ. 2013, 55, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, A.; Ferreri, C.; Filipovic, M.; Ivanovic-Burmazovic, I.; Stancic, A.; Otasevic, V.; Korac, A.; Buzadzic, B.; Korac, B. Targeting the superoxide/nitric oxide ratio by L-arginine and SOD mimic in diabetic rat skin. Free Radic. Res. 2016, 50, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Ben Khedir, S.; Moalla, D.; Jardak, N.; Mzid, M.; Sahnoun, Z.; Rebai, T. Pistacia lentiscus fruit oil reduces oxidative stress in human skin explants caused by hydrogen peroxide. Biotech. Histochem. 2016, 91, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.; Haldar, C. UVB irradiation severely induces systemic tissue injury by augmenting oxidative load in a tropical rodent: Efficacy of melatonin as an antioxidant. J. Photochem. Photobiol. B 2014, 141, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Chen, H.; Niu, C.; Hu, J.; Cao, B. Protective effect of Schizandrin B against damage of UVB irradiated skin cells depend on inhibition of inflammatory pathways. Bioengineered 2017, 8, 36–44. [Google Scholar] [CrossRef]
- Robinson, E.S.; Werth, V.P. The role of cytokines in the pathogenesis of cutaneous lupus erythematosus. Cytokine 2015, 73, 326–334. [Google Scholar] [CrossRef]
- Chen, C.C.; Chiang, A.N.; Liu, H.N.; Chang, Y.T. EGb-761 prevents ultraviolet B-induced photoaging via inactivation of mitogen-activated protein kinases and proinflammatory cytokine expression. J. Dermatol. Sci. 2014, 75, 55–62. [Google Scholar] [CrossRef]
- Pacholczyk, M.; Czernicki, J.; Ferenc, T. The effect of solar ultraviolet radiation (UVR) on induction of skin cancers. Med. PR 2016, 67, 255–266. [Google Scholar] [CrossRef]
- Hwang, E.; Ngo, H.T.T.; Park, B.; Seo, S.A.; Yang, J.E.; Yi, T.H. Myrcene, an aromatic volatile compound, ameliorates human skin extrinsic aging via regulation of MMPs production. Am. J. Chin. Med. 2017, 45, 1113–1124. [Google Scholar] [CrossRef]
- Choi, E.M.; Lee, Y.S. Luteolin suppresses IL-1beta-induced cytokines and MMPs production via p38 MAPK, JNK, NF-kappaB and AP-1 activation in human synovial sarcoma cell line, SW982. Food Chem. Toxicol. 2010, 48, 2607–2611. [Google Scholar] [CrossRef]
- Hasegawa, T.; Nakashima, M.; Suzuki, Y. Nuclear DNA damage-triggered NLRP3 inflammasome activation promotes UVB-induced inflammatory responses in human keratinocytes. Biochem. Biophys. Res. Commun. 2016, 477, 329–335. [Google Scholar] [CrossRef]
- Lee, D.H.; Oh, J.H.; Chung, J.H. Glycosaminoglycan and proteoglycan in skin aging. J. Dermatol. Sci. 2016, 83, 174–181. [Google Scholar] [CrossRef]
- Makpol, S.; Azura Jam, F.; Anum Mohd Yusof, Y.; Zurinah Wan Ngah, W. Modulation of collagen synthesis and its gene expression in human skin fibroblasts by tocotrienol-rich fraction. Arch. Med. Sci. 2011, 7, 889–895. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Z.; Cai, X.; Sun, Y.; Zhao, C.; Liu, F.; Liu, D. Effects of dimethylaminoethanol and compound amino acid on D-galactose induced skin aging model of rat. Sci. World J. 2014, 2014, 507351. [Google Scholar] [CrossRef]
- Song, H.; Zhang, S.; Zhang, L.; Li, B. Effect of orally administered collagen peptides from bovine bone on skin aging in chronologically aged mice. Nutrients 2017, 9, 1209. [Google Scholar] [CrossRef]
- Devi, S.; Kumar, N.; Kapila, S.; Mada, S.B.; Reddi, S.; Vij, R.; Kapila, R. Buffalo casein derived peptide can alleviates H2O2 induced cellular damage and necrosis in fibroblast cells. Exp. Toxicol. Pathol. 2017, 69, 485–495. [Google Scholar] [CrossRef]
- Van Doren, S.R. Matrix metalloproteinase interactions with collagen and elastin. Matrix Biol. 2015, 44–46, 224–231. [Google Scholar] [CrossRef]
- Mali, A.V.; Joshi, A.A.; Hegde, M.V.; Kadam, Sh.S. Enterolactone suppresses proliferation, migration and metastasis of MDA-MB-231 breast cancer cells through inhibition of uPA induced plasmin activation and MMPs-mediated ECM remodeling. Asian Pac. J. Cancer Prev. 2017, 18, 905–915. [Google Scholar]
- Shih, M.F.; Pan, K.H.; Cherng, J.Y. Possible mechanisms of Di(2-ethylhexyl) phthalate-induced MMP-2 and MMP-9 expression in A7r5 rat vascular smooth muscle cells. Int. J. Mol. Sci. 2015, 16, 28800–28811. [Google Scholar] [CrossRef]
- Huang, M.; Yang, H.; Zhu, L.; Li, H.; Zhou, J.; Zhou, Z. Inhibition of connective tissue growth factor attenuates paraquat-induced lung fibrosis in a human MRC-5 cell line. Environ. Toxicol. 2016, 31, 1620–1626. [Google Scholar] [CrossRef]
- Kim, J.A.; Ahn, B.N.; Kong, C.S.; Kim, S.K. Protective effect of chromene isolated from Sargassum horneri against UV-A-induced damage in skin dermal fibroblasts. Exp. Dermatol. 2012, 21, 630–631. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Nishimura, R.; Nozaki, K.; Sasaki, N.; Kadosawa, T.; Goto, N.; Date, M.; Takeuchi, A. Collagen production and maturation at the experimental ligament defect stimulated by pulsing electromagnetic fields in rabbits. J. Vet. Med. Sci. 1993, 55, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Czekalla, C.; Schönborn, K.H.; Döge, N.; Jung, S.; Darvin, M.E.; Lademann, J.; Meinke, M.C. Impact of body site, age, and gender on the collagen/elastin index by noninvasive in vivo vertical two-photon microscopy. Skin Pharmacol. Physiol. 2017, 30, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, F.T.; Fonseca, Y.M.; Pitol, D.L.; Iyomasa, M.M.; Bentley, M.V.; Fonseca, M.J. Evaluation of protective effect of a water-in-oil microemulsion incorporating quercetin against UVB-induced damage in hairless mice skin. J. Pharm. Pharm. Sci. 2010, 13, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Bagdas, D.; Gul, N.Y.; Topal, A.; Tas, S.; Ozyigit, M.O.; Cinkilic, N.; Gul, Z.; Etoz, B.C.; Ziyanok, S.; Inan, S.; et al. Pharmacologic overview of systemic chlorogenic acid therapy on experimental wound healing. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 1101–1116. [Google Scholar] [CrossRef] [PubMed]
- Henry-Vitrac, C.; Ibarra, A.; Roller, M.; Mérillon, J.M.; Vitrac, X. Contribution of chlorogenic acids to the inhibition of human hepatic glucose-6-phosphatase activity in vitro by Svetol, a standardized decaffeinated green coffee extract. J. Agric. Food Chem. 2010, 58, 4141–4144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.W.; Ji, T.; Su, S.L.; Shang, E.X.; Guo, S.; Guo, J.M.; Qian, D.W.; Duan, J.A. Pharmacokinetics of Mori Folium flavones and alkaloids in normal and diabetic rats. Zhongguo Zhong Yao Za Zhi 2017, 42, 4218–4225. [Google Scholar] [PubMed]
Sample Availability: Samples of the Ilex kudingcha C. J. Tseng polyphenols are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Constituent | Retention Time (min) | Content (mg/g) | Theoretical Plate Number | Resolution |
|---|---|---|---|---|---|
| 1 | Chlorogenic acid | 12.623 | 79.71 | 22586 | 2.56 |
| 2 | Cryptochlorogenic acid | 13.453 | 16.02 | 29063 | 34.37 |
| 3 | Isochlorogenic acid B | 22.220 | 52.53 | 206572 | 1.88 |
| 4 | Isochlorogenic acid A | 22.613 | 120.03 | 164025 | 11.05 |
| 5 | Isochlorogenic acid C | 25.210 | 96.42 | 165480 | 5.33 |
| Group | T-SOD (U/mL) | CAT (U/mL) | MDA (nmol/mL) |
|---|---|---|---|
| Healthy | 107.36 ± 5.71 a | 13.36 ± 0.31 a | 10.62 ± 0.25 e |
| UVB-treated | 68.71 ± 3.44 e | 7.85 ± 0.25 e | 21.03 ± 0.65 a |
| VC | 84.34 ± 3.02 c | 9.76 ± 0.25 c | 15.21 ± 0.42 c |
| LKTP | 77.62 ± 2.61 d | 8.99 ± 0.18 d | 18.33 ± 0.61 b |
| HKTP | 93.56 ± 3.26 b | 11.89 ± 0.32 b | 13.29 ± 0.46 d |
| Group | IL-6 (pg/mL) | IL-1β (pg/mL) | TNF-α (pg/mL) |
|---|---|---|---|
| Healthy | 10.77 ± 0.35 e | 37.52 ± 1.59 e | 44.36 ± 3.78 e |
| UVB-treated | 18.33 ± 0.61 a | 64.39 ± 3.15 a | 71.26 ± 4.58 a |
| VC | 14.37 ± 0.36 c | 50.32 ± 2.87 c | 52.34 ± 2.30 c |
| LKTP | 16.11 ± 0.28 b | 57.17 ± 1.88 b | 64.59 ± 3.51 b |
| HKTP | 12.58 ± 0.32 d | 44.02 ± 2.03 d | 47.33 ± 1.89 d |
| Group | Col I (mg/g) | Col III (mg/g) | Hydroxyproline (mg/g) | Hyaluronic Acid (μg/g) |
|---|---|---|---|---|
| Healthy | 21.33 ± 2.77 a | 5.42 ± 0.82 e | 81.36 ± 5.69 a | 536.72 ± 27.86 a |
| UVB-treated | 7.35 ± 1.21 e | 19.37 ± 1.67 a | 32.02 ± 3.35 e | 148.36 ± 13.53 e |
| VC | 12.36 ± 1.58 c | 10.35 ± 1.28 c | 52.36 ± 2.92 c | 312.05 ± 22.38 c |
| LKTP | 9.78 ± 0.68 d | 13.14 ± 2.10 b | 46.20 ± 2.43 d | 277.06 ± 18.75 d |
| HKTP | 16.39 ± 2.28 b | 7.79 ± 0.63 d | 66.71 ± 4.86 b | 441.25 ± 25.16 b |
| Time/min | Mobile Phase A% | Mobile Phase B% |
|---|---|---|
| 0 | 12 | 88 |
| 0–20 | 12→44 | 88→56 |
| 20–23 | 44→50 | 56→50 |
| 23–30 | 50→55 | 50→45 |
| Forward Sequence | Reverse Sequence | |
|---|---|---|
| TIMP-1 | 5′-ATGCGGCCGCATGATGGCCCCCTTTGCATC-3′ | 5′-ATCCCGGGTCATCGGGCCCCAAGGGATC-3′ |
| TIMP-2 | 5′-GTAGTGATCAGGGCCAAAG-3′ | 5′-TTCTCTGTGACCCAGTCCAT-3′ |
| MMP-2 | 5′-ACCGAGGACTATGACCGGGATAA-3′ | 5′-GTCCTCATACTTGTTGCCCAGGA-3′ |
| MMP-9 | 5′-GCCCTGGAACTCACACGACA-3′ | 5′-TTGGAAATCCACACGCCAGAAG-3 |
| iNOS | 5′-GTTCTCAGCCCAACAATACAAGA-3′ | 5′-GTGGACGGGTCGATGTCAC-3 |
| Cu/Zn-SOD | 5′-AACCAGTTGTGTTGTCAGGAC-3′ | 5′-CCACCATGTTTCTTAGAGTGAGG-3′ |
| Mn-SOD | 5′-CAGACCTGCCTTACGACTATGG-3′ | 5′-CTCGGTGGCGTTGAGATTGTT-3′ |
| CAT | 5′-GGAGGCGGGAACCCAATAG-3′ | 5′-GTGTGCCATCTCGTCAGTGAA-3 |
| GSH-Px | 5′-CCACCGTGTATGCCTTCTCC-3′ | 5′-AGAGAGACGCGACATTCTCAAT-3′ |
| GAPDH | 5′-AGGTCGGTGTGAACGGATTTG-3′ | 5′-GGGGTCGTTGATGGCAACA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, R.; Zhang, J.; Sun, P.; Qian, Y.; Zhao, X. Protective Effects of Kuding Tea (Ilex kudingcha C. J. Tseng) Polyphenols on UVB-Induced Skin Aging in SKH1 Hairless Mice. Molecules 2019, 24, 1016. https://doi.org/10.3390/molecules24061016
Yi R, Zhang J, Sun P, Qian Y, Zhao X. Protective Effects of Kuding Tea (Ilex kudingcha C. J. Tseng) Polyphenols on UVB-Induced Skin Aging in SKH1 Hairless Mice. Molecules. 2019; 24(6):1016. https://doi.org/10.3390/molecules24061016
Chicago/Turabian StyleYi, Ruokun, Jing Zhang, Peng Sun, Yu Qian, and Xin Zhao. 2019. "Protective Effects of Kuding Tea (Ilex kudingcha C. J. Tseng) Polyphenols on UVB-Induced Skin Aging in SKH1 Hairless Mice" Molecules 24, no. 6: 1016. https://doi.org/10.3390/molecules24061016
APA StyleYi, R., Zhang, J., Sun, P., Qian, Y., & Zhao, X. (2019). Protective Effects of Kuding Tea (Ilex kudingcha C. J. Tseng) Polyphenols on UVB-Induced Skin Aging in SKH1 Hairless Mice. Molecules, 24(6), 1016. https://doi.org/10.3390/molecules24061016