
Identification and Characterization of the Caspase-Mediated Apoptotic Activity of Teucrium mascatense and an Isolated Compound in Human Cancer Cells




Abstract
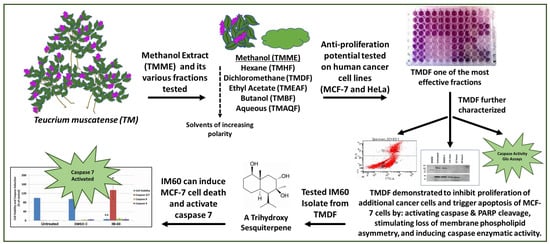
1. Introduction
2. Results
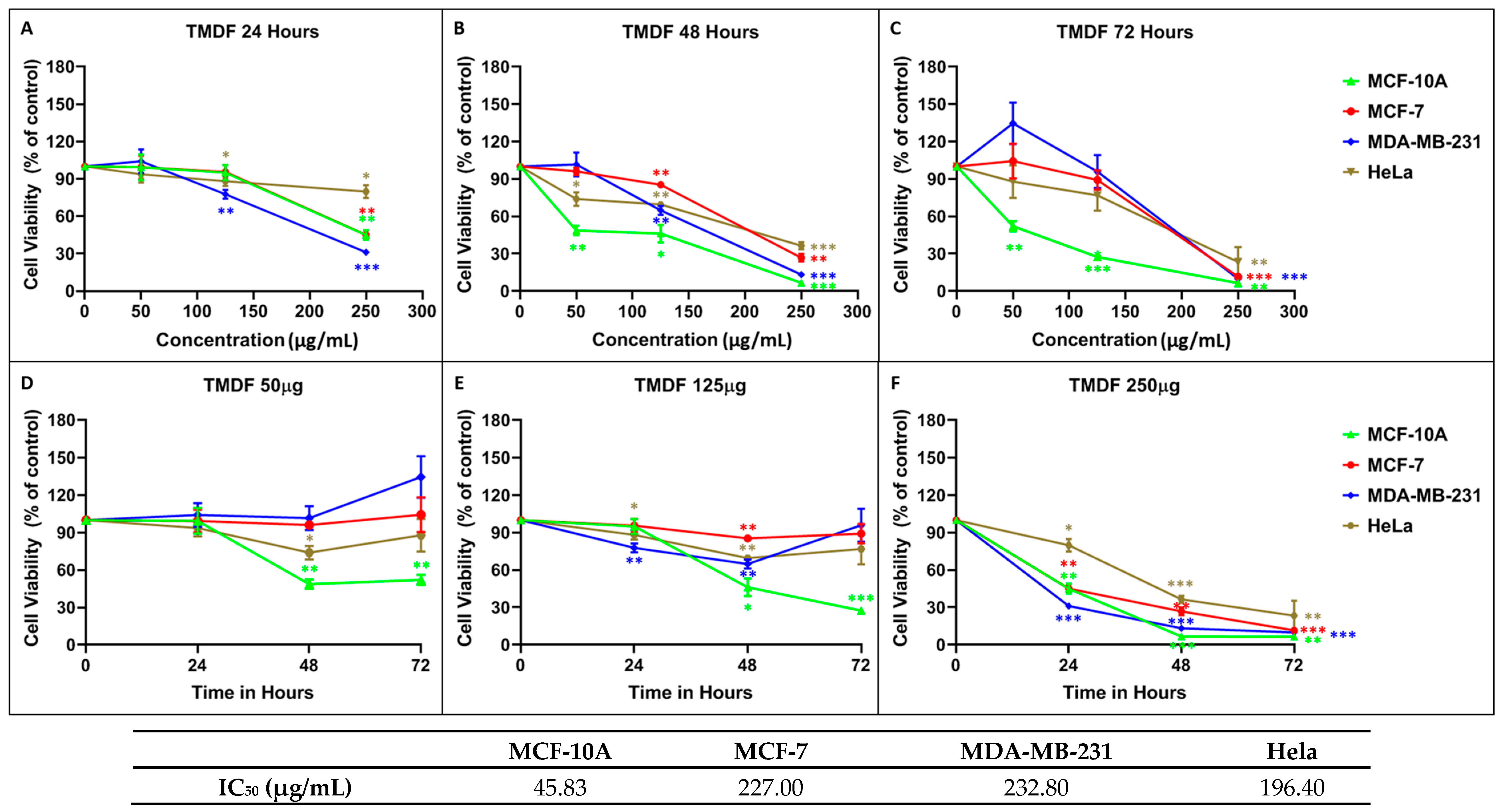
2.1. Crude Methanolic Extract of T. mascatense and Some of Its Fractions Induce Cytotoxic Effects in Human Normal and Cancer Cell Lines
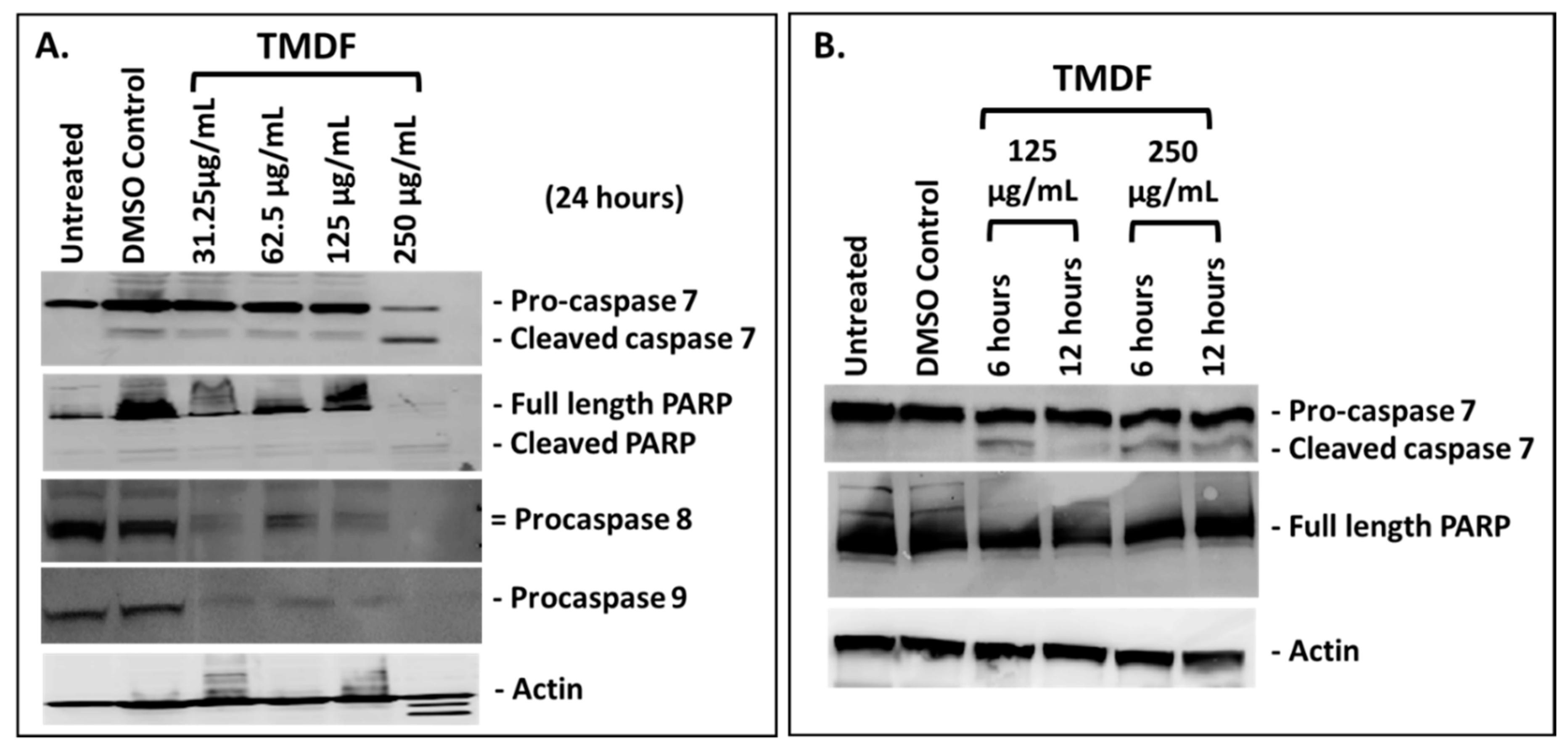
2.2. TMDF Activates Key Apoptotic Proteins in Breast Cancer Cells
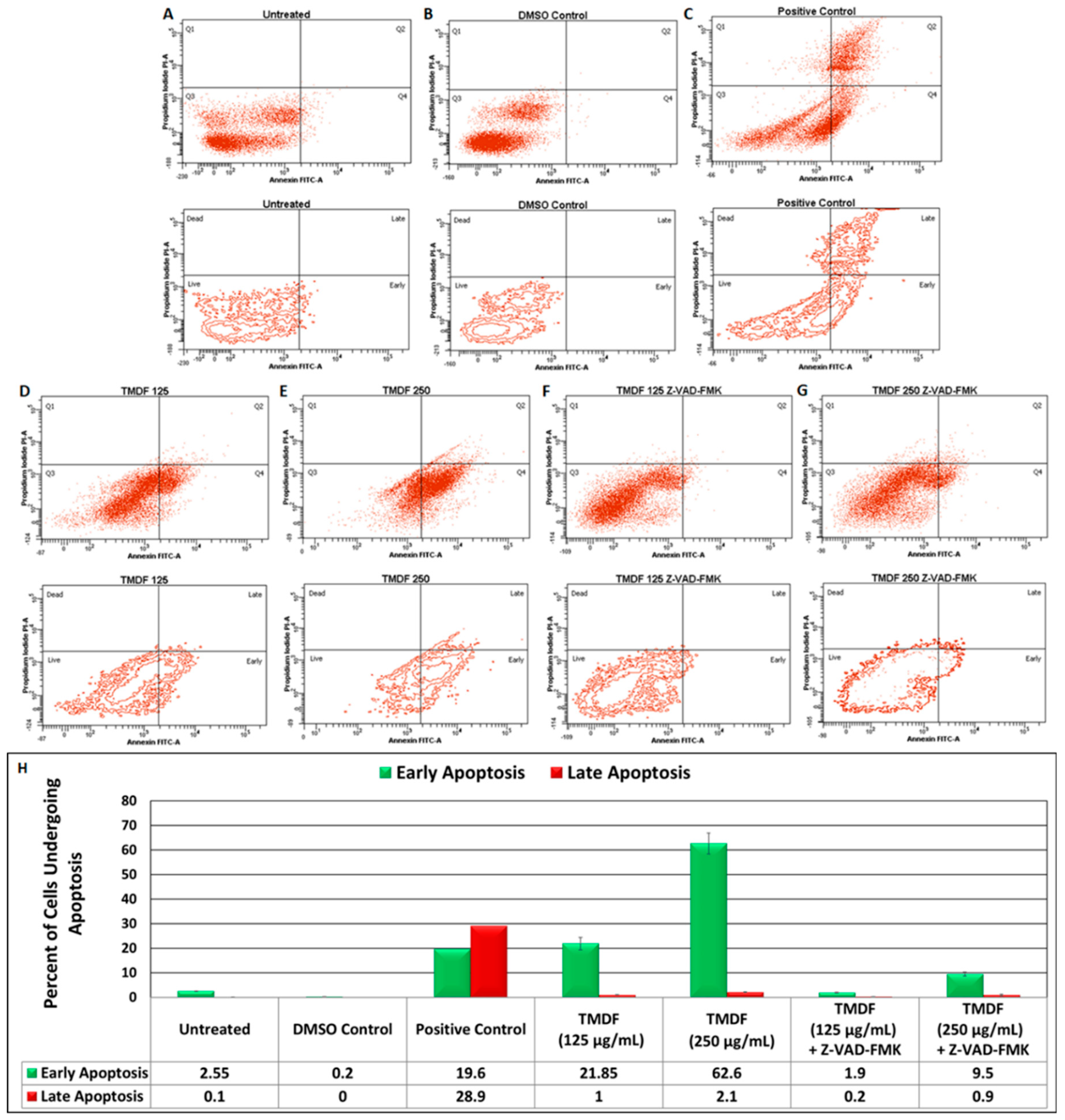
2.3. TMDF Can Induce Apoptosis in MCF-7 Cells
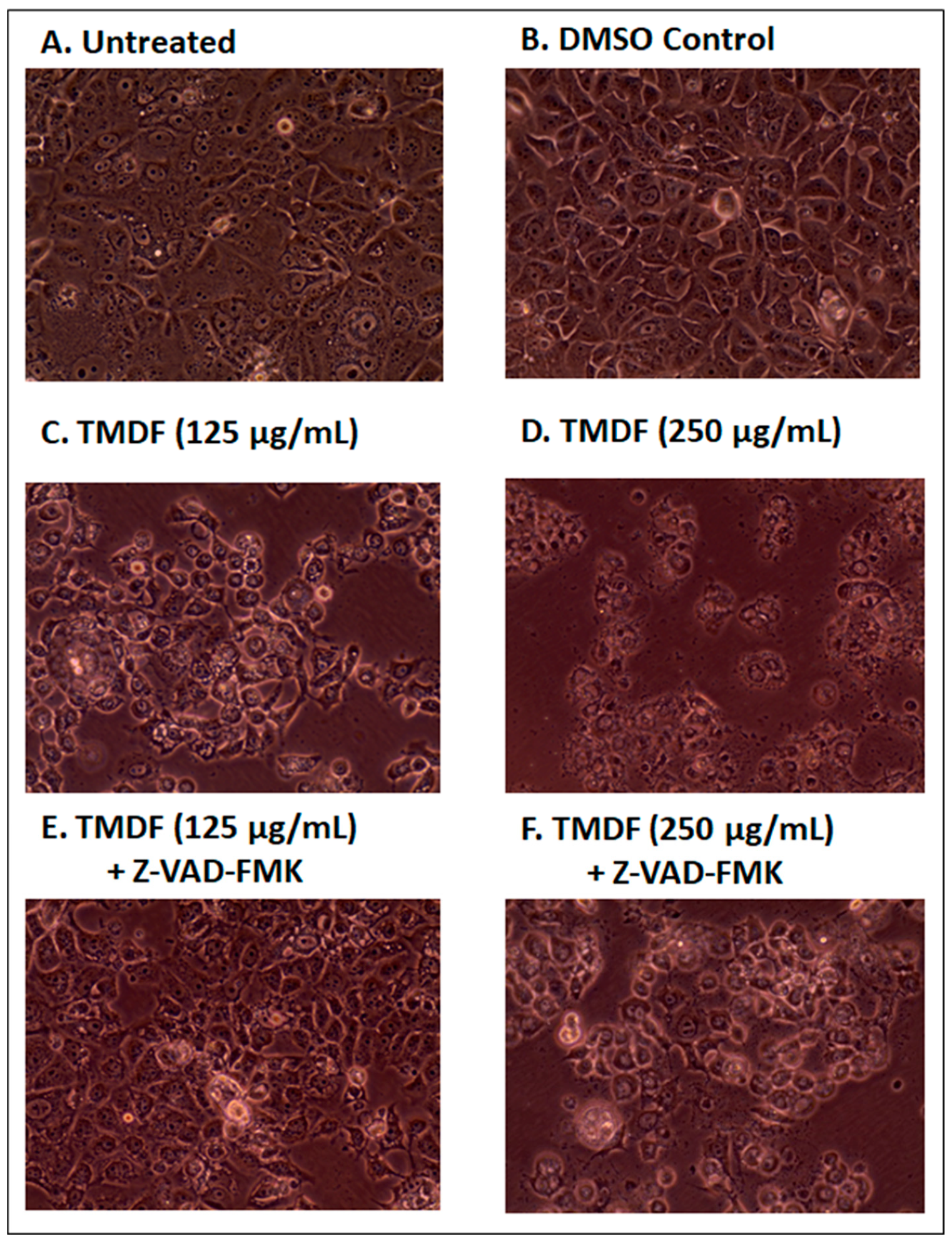
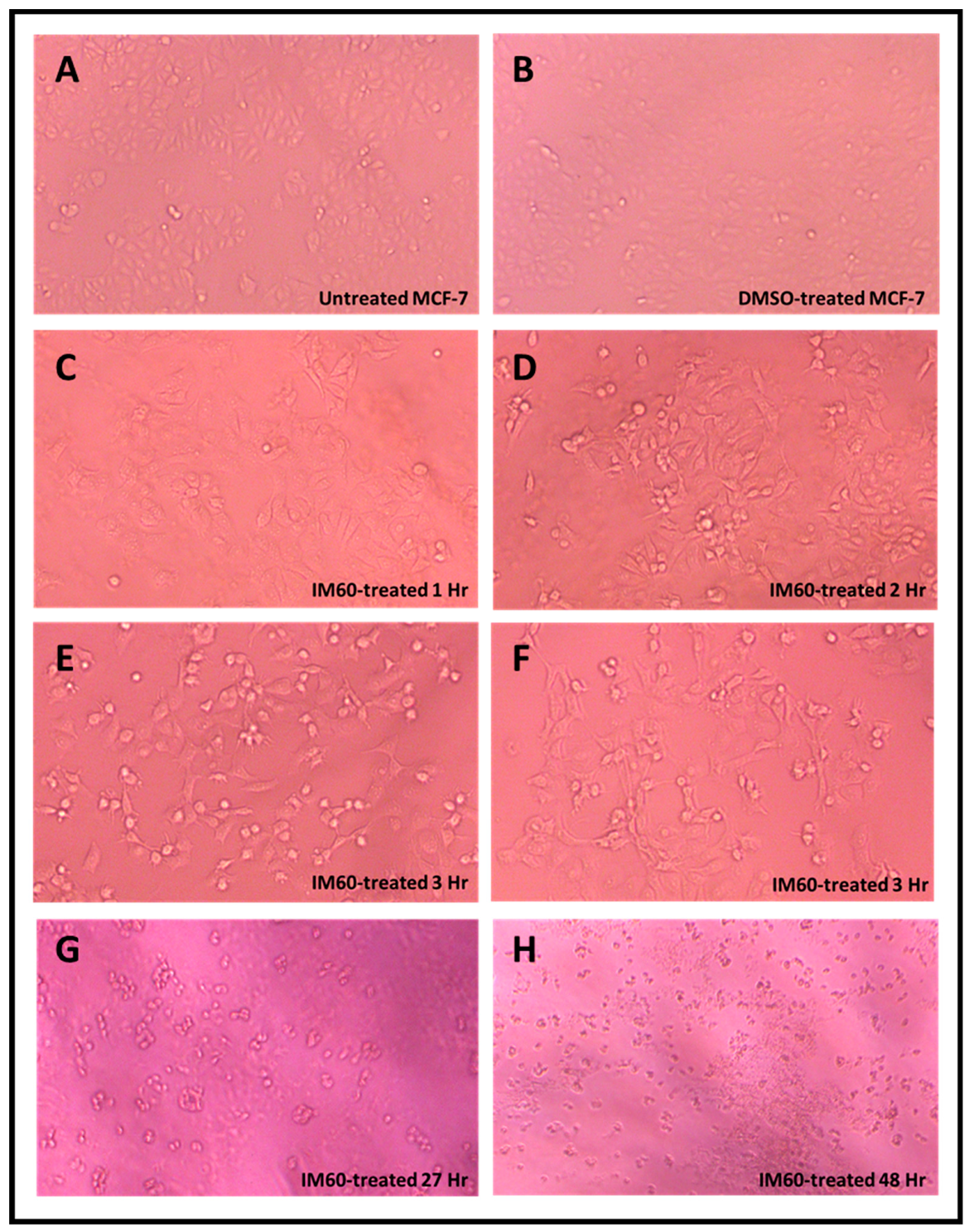
2.4. Effects of TMDF on the Morphological Properties of Treated Cells
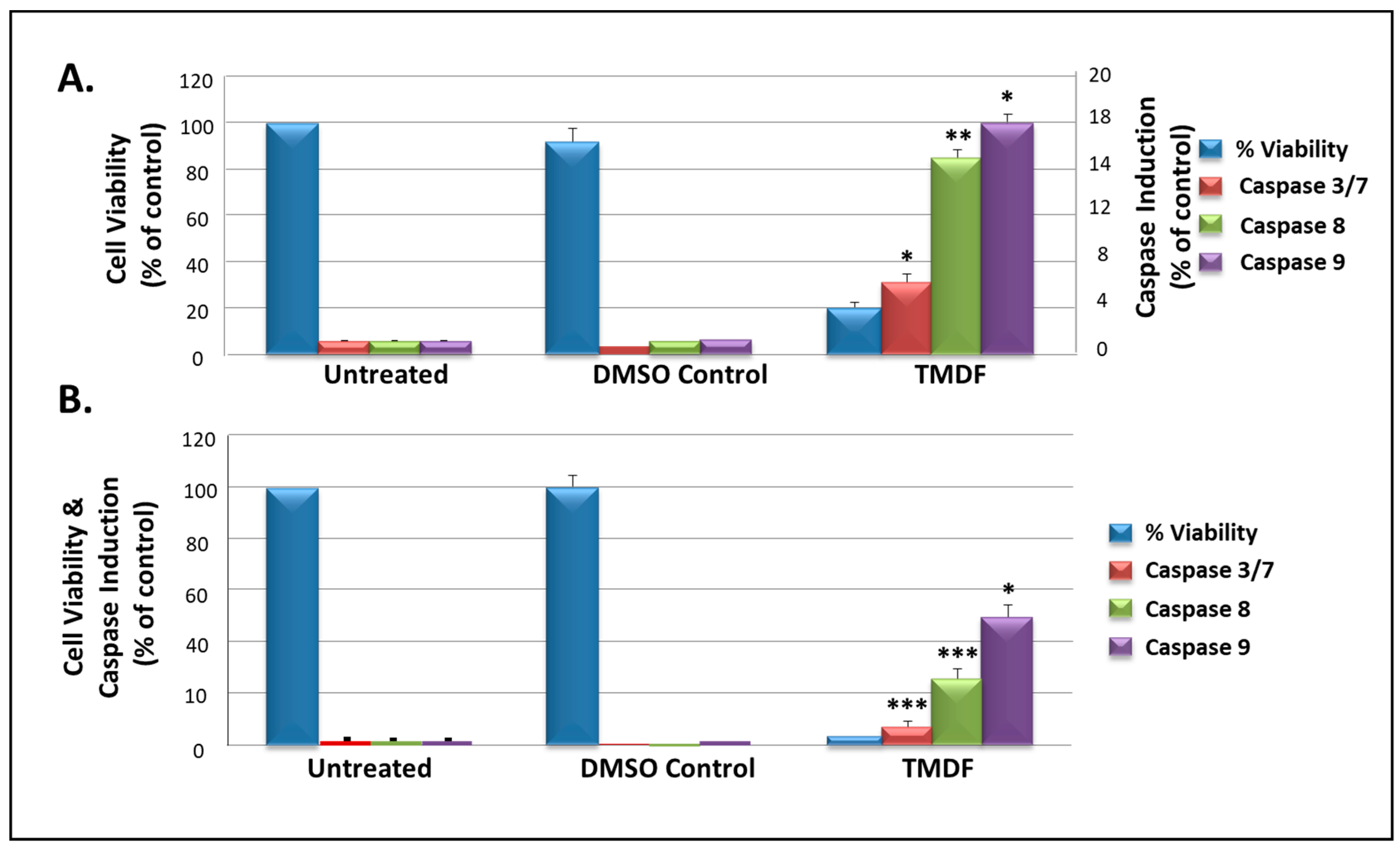
2.5. TMDF Activates Caspase Activity in Human Breast Cancer Cells
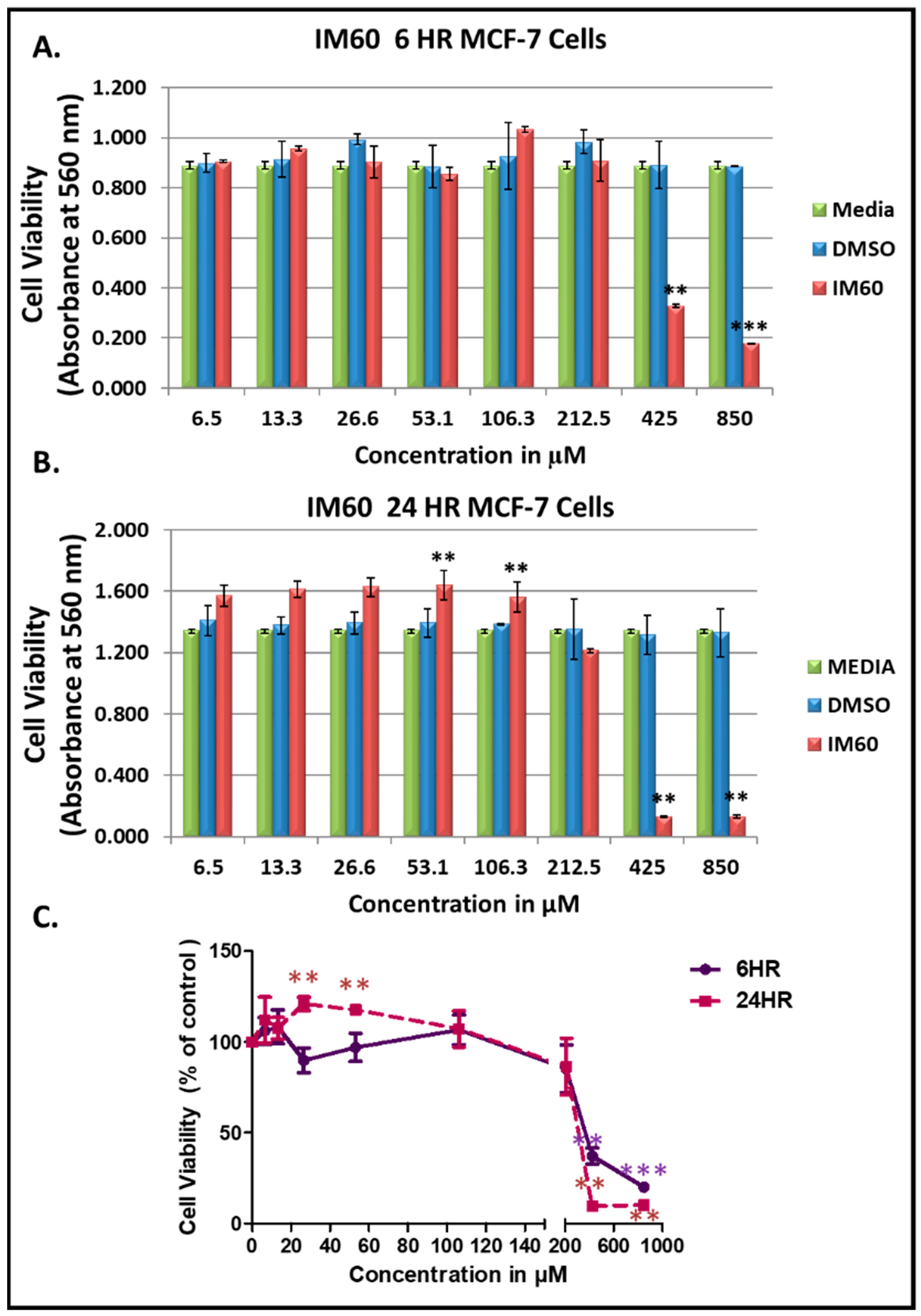
2.6. Test of the Anti-Proliferation Effect of IM60 on Breast Cancer Cells
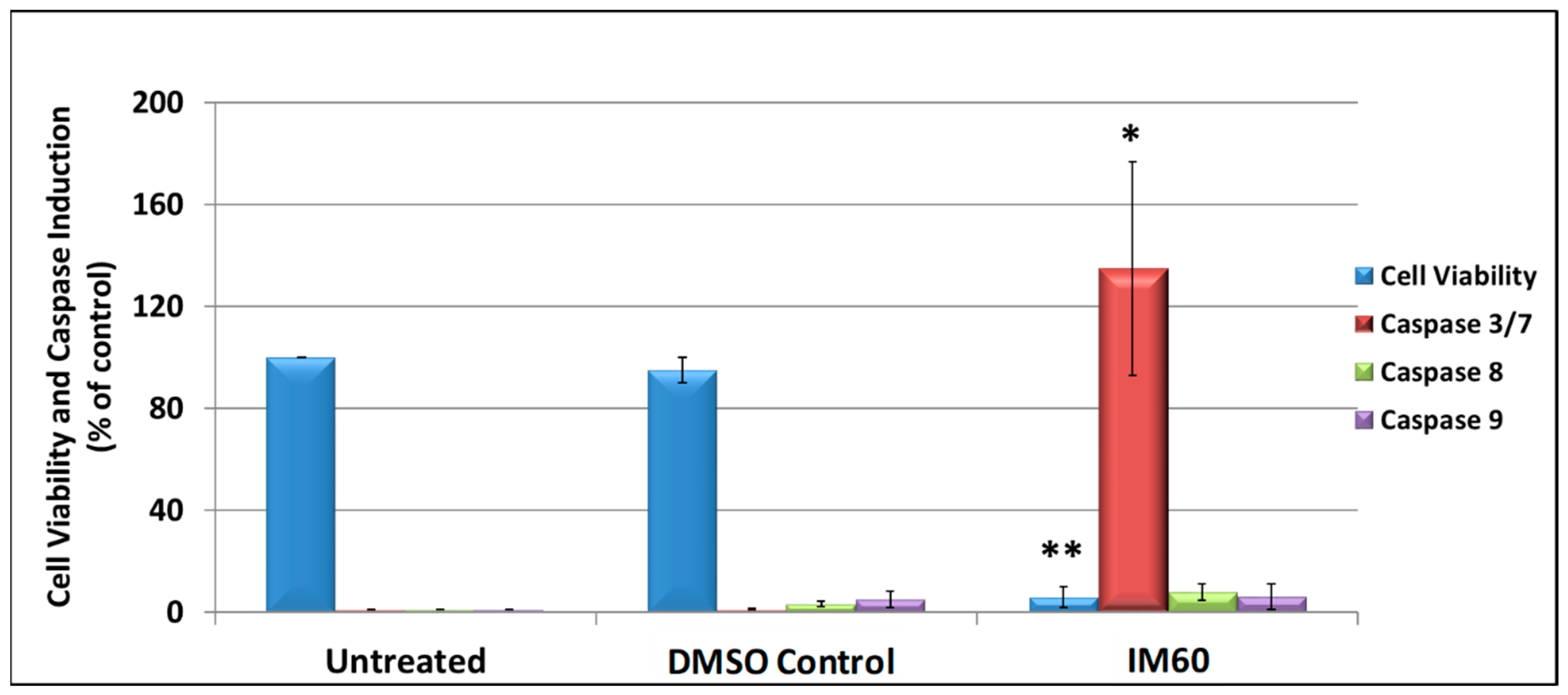
2.7. IM60 Can Induce Caspase Activity in MCF-7 Cells
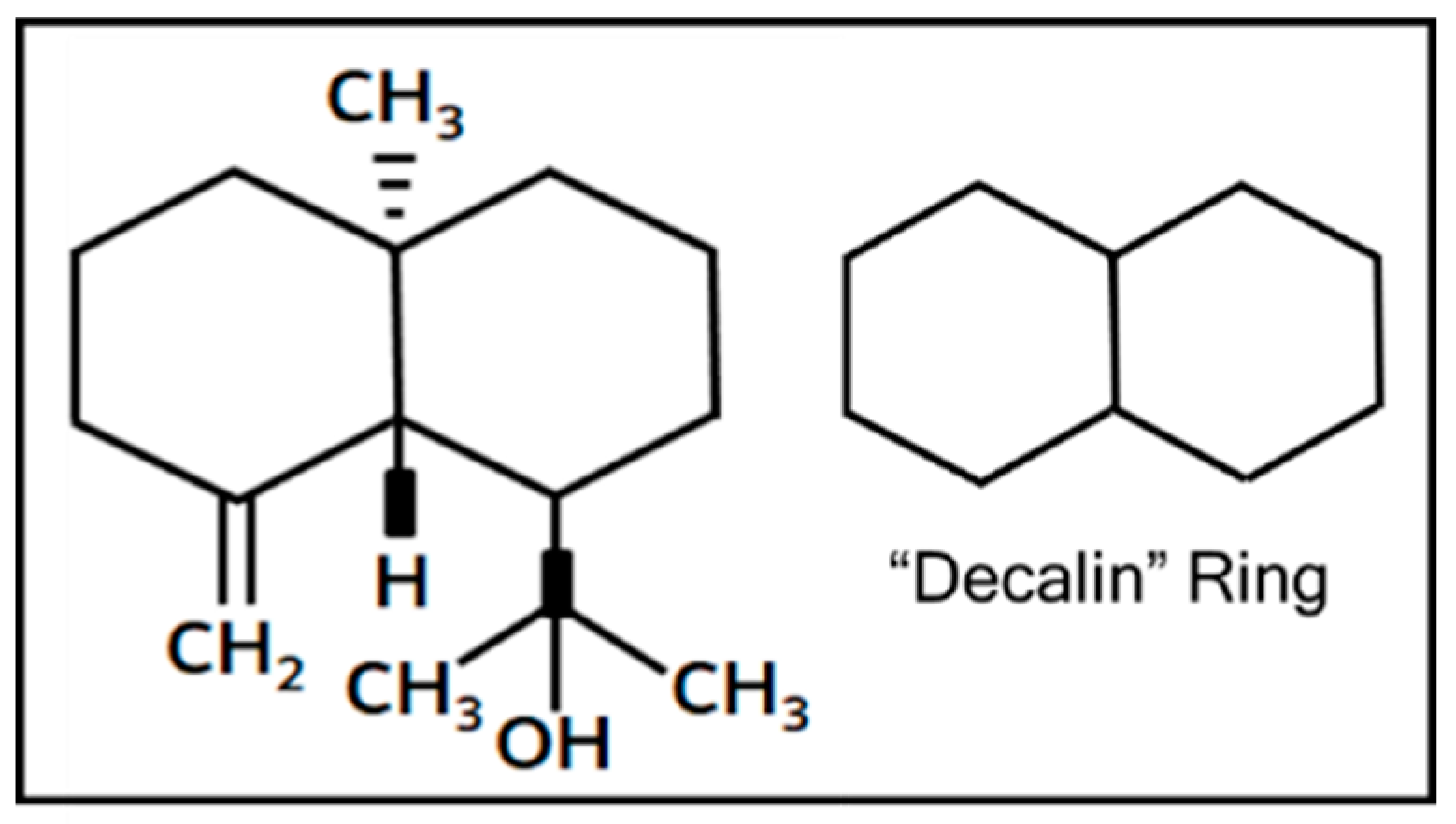
3. Discussion
4. Materials and Methods
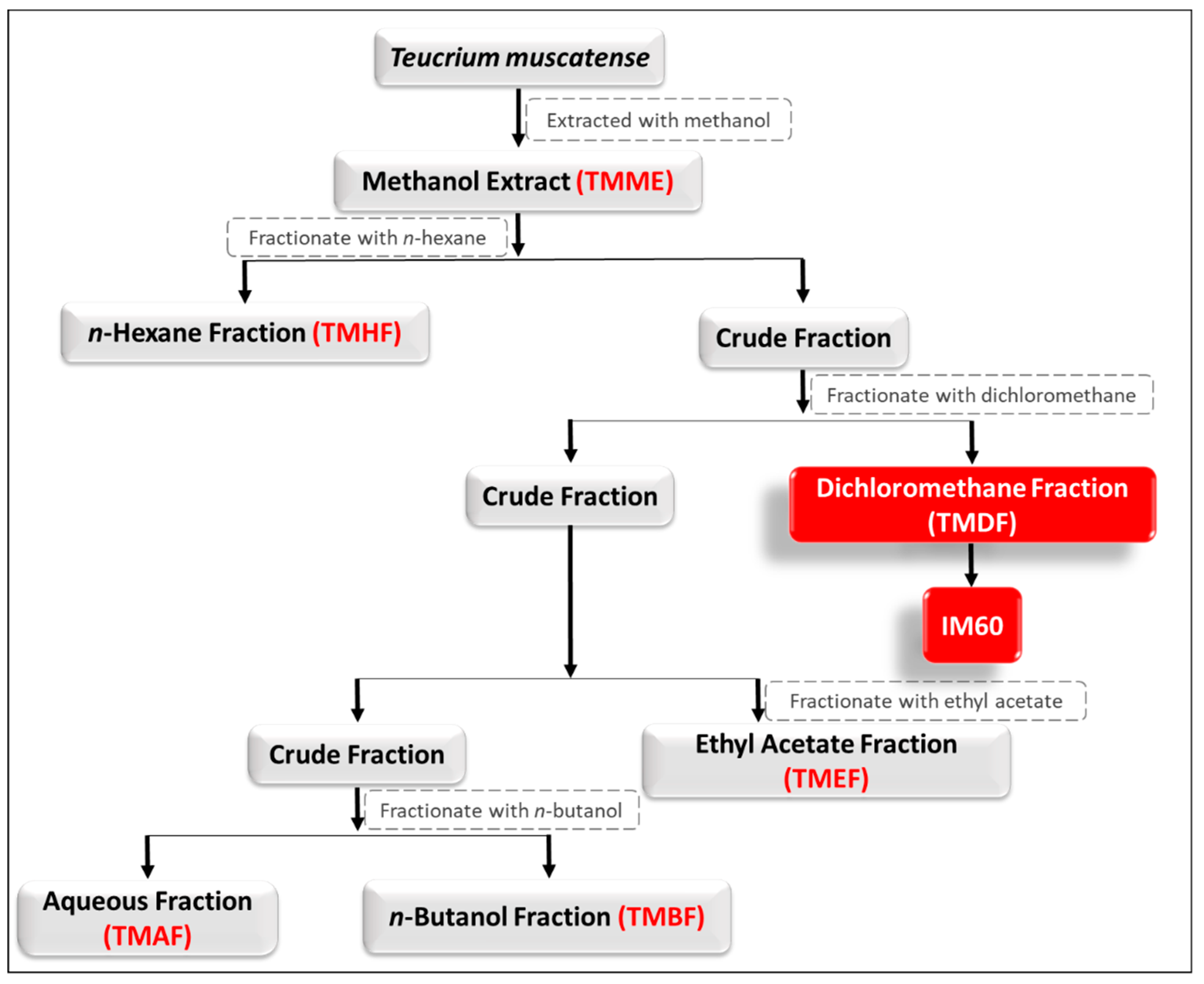
4.1. Extraction and Isolation of Plant Material
4.2. Cell Lines and Culture Medium
4.3. MTT Cell Proliferation Assay
4.4. Cell Titer-GLO Cell Viability and Caspase-GLO 3/7, 8 & 9 Luminescent Assays
4.5. Morphological Studies
4.6. Western Blot Analysis
4.7. Annexin V/Propidium Iodide Staining
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Cancer: Key Facts. Geneva, Switzerland. Available online: http://www.who.int/mediacentre/factsheets/fs297/en/ (accessed on 1 February 2018).
- GLOBOCAN 2018: Estimated Cancer Incidence, Mortality and Prevalence Worldwide in 2018. 2018. Available online: http://gco.iarc.fr/ (accessed in 1 September 2018).
- Stewart, B.W.; Wild, C.P. (Eds.) World Cancer Report 2014; International Agency for Research on Cancer: Lyon, France, 2014. [Google Scholar]
- Cragg, G.M.; Newman, D.J.; Yang, S.S. Natural product extracts of plant and marine origin having antileukemia potential. The NCI experience. J. Nat. Prod. 2006, 69, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Khan, H. Medicinal Plants in Light of History: Recognized Therapeutic Modality. J. Evid. Based Complement. Altern. Med. 2014, 19, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Verpoorte, R. Pharmacognosy in the New Millennium: Lead finding and Biotechnology. J. Pharm. Pharmacol. 2000, 52, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Levitsky, D.O.; Dembitsky, V.M. Anti-breast Cancer Agents Derived from Plants. Nat. Prod. Bioprospect. 2014, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tariq, A.; Sadia, S.; Pan, K.; Ullah, I.; Mussarat, S.; Sun, F.; Abiodun, O.O.; Batbaatar, A.; Li, Z.; Song, D.; et al. A systematic review on ethnomedicines of anti-cancer plants. Phytother. Res. 2017, 31, 202–264. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, J.M.; Romane, A.; Efferth, T.; Salgueiro, L. North African Medicinal Plants Traditionally Used in Cancer Therapy. Front. Pharmacol. 2017, 8, 383. [Google Scholar] [CrossRef] [PubMed]
- Richards, T.S.; Kamdje, A.H.N.; Mukhtar, F. Medicinal Plants in Breast Cancer Therapy. J. Dis. Med. Plants 2015, 1, 19–23. [Google Scholar]
- Western, A.R. The Flora of United Arab Emirates, an Introduction; Publication of the UAE University: Sharjah, UAE, 1986. [Google Scholar]
- El-Ghonemy, A.A. Encyclopedia of Medicinal Plants of the United Emirates, 1st ed.; University of UAE: Sharjah, UAE, 1993. [Google Scholar]
- Ghazanfar, S.A. Handbook of Arabian Medicinal Plants; Library of Congress: Washington, DC, USA, 1994. [Google Scholar]
- Jongbloed, M.V. The Comprehensive Guide to the Wild Flowers of the United Arab Emirates, ERWDA; Emirates Printing Press: Dubai, UAE, 2003. [Google Scholar]
- Miller, A.G.; Morris, M. Plants of Dhofar, The southern Region of Oman: Traditional, Economic and Medicinal Uses; Office of the Advisor for Conservation of the Environment: Muscat, Oman, 1987.
- Zaragari, A. Medicinal Plants, 6th ed.; Tehran University Publications: Tehran, Iran, 1994. [Google Scholar]
- Tutin, T.G.; Heywood, V.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea III. Genus teucrium L.; Cambridge Press: Cambridge, UK, 1972. [Google Scholar]
- Gharaibeh, M.N.; Elayan, H.H.; Salhab, A.S. Hypoglycemic effects of Teucrium polium. J. Ethnopharmacol. 1988, 24, 93–99. [Google Scholar] [CrossRef]
- Rudolf-Pfaendler, H.; Piozzi, F.; Rodriguez, B.; Savona, G. Advances in the chemistry of the Furanoditerphenoids from Teucrium species. Heterocycles 1987, 25, 807–841. [Google Scholar] [CrossRef]
- Suleiman, M.S.; Abdul-Ghani, A.S.; Al-Khalil, S.; Amin, R. Effect of Teucrium polium boiled leaf extract on intestinal motility and blood pressure. J. Ethnopharmacol. 1988, 22, 111–116. [Google Scholar] [CrossRef]
- Stankovic, M.; Topuzovic, M.; Solujic, S.; Mihailovic, V. Antioxidant activity and concentration of phenols and flavonoids in the whole plant and plant parts of Teucrium chamaerdys L. var. glanduliferum Haussk. J. Med. Plant Res. 2010, 4, 2092–2098. [Google Scholar]
- Shahraki, M.R.; Arab, M.R.; Mirimokaddam, E.; Palan, M.J. The effect of Teucrium polium (Calpoureh) on liver function, serum lipids and glucose in diabetic male rats. Iran Biomed. J. 2007, 11, 65–68. [Google Scholar] [PubMed]
- Rasekh, H.R.; Khoshnood-Mansourkhani, M.J.; Kamalinejad, M. Hypolipidemic, Effects of Teucrium polium in rats. Fitoterapia 2001, 72, 937–939. [Google Scholar] [CrossRef]
- Esmaeili, M.A.; Yazdanparast, R. Hypoglycaemic effect of Teucrium polium: Studies with rat pancreatic islets. J. Ethnopharmacol. 2004, 95, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Panovska, T.K.; Kulevanova, S.; Gjorgoski, I.; Bogdanova, M.; Petrushevska, G. Hepatoprotective effect of the ethyl acetate extract of Teucrium polium L. against carbon tetrachloride-induced hepatic injury in rats. Acta Pharm. 2007, 57, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Tariq, M.; Ageel, A.M.; al-Yahya, M.A.; Mossa, J.S.; al-Said, M.S. Anti-inflammatory activity of Teucrium polium. Int. J. Tissue React. 1989, 11, 185–188. [Google Scholar] [PubMed]
- Heidari, M.; Karaminezhad, R.; Davvand, E.; Jalali, S. Evaluation of the analgesic effect of Teucrium polium extract in mice. DARU J. Pharm. Sci. 2001, 24, 277–285. [Google Scholar]
- Kadifkova Panovska, T.; Kulevanova, S.; Stefova, M. In vitro antioxidant activity of some Teucrium species (Lamiaceae). Acta Pharm. 2005, 55, 207–214. [Google Scholar] [PubMed]
- Hasani, P.; Yasa, N.; Vosough-Ghanbari, S.; Mohammadirad, A.; Dehghan, G.; Abdollahi, M. In vivo antioxidant potential of Teucrium polium, as compared to alpha-tocopherol. Acta Pharm. 2007, 57, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Jurisic, R.; Vladimir-Knezevic, S.; Kalodera, Z.; Grgic, J. Determination of selenium in Teucrium species by hydride generation atomic absorption spectrometry. Z. Naturforsch. C 2003, 58, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Stanković, M.; Topuzović, M.; Marković, A.; Pavlovic, D.; Solujic, S.; Niciforovic, N.; Mihailovic, V. Antioxidant Activity, Phenol and flavonoid contents of different Teucrium chamaedrys L. extracts. Biotechnol Equip 2010, 24, 82–86. [Google Scholar] [CrossRef]
- Camarda, I. Ricerche etnobotaniche nel comune di Dorgali (Sardegna centro-orientale). Bollettino della Società Sarda di Scienze Naturali 1990, 27, 147–204. [Google Scholar]
- Bahramikia, S.; Yazdanparast, R. Phytochemistry and medicinal properties of Teucrium polium L. (Lamiaceae). Phytother. Res. 2012, 26, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Rajabalian, S. Methanolic extract of Teucrium polium L. potentiates the cytotoxic and apoptotic effects of anticancer drugs of vincristine, vinblastine and doxorubicin against a panel of cancerous cell lines. Exp. Oncol. 2008, 30, 133–138. [Google Scholar] [PubMed]
- Bedir, E.; Tasdemir, D.; Çalis, I.; Zerbe, O.; Sticher, O. Neo-clerodane diterpenoids from Teucrium polium. Phytochemistry 1999, 51, 921–925. [Google Scholar] [CrossRef]
- Piozzi, F. Further researches on the furoclerodanes from Teucrium species. Heterocycles 1994, 37, 603–626. [Google Scholar] [CrossRef]
- Piozzi, F.; Bruno, M.; Rosselli, S. Further furoclerodanes from Teucrium genus. Heterocycles 1998, 48, 2185–2203. [Google Scholar] [CrossRef]
- Acamovic, T.; Brooker, J.D. Biochemistry of plant secondary metabolites and their effects in animals. Proc. Nutr. Soc. 2005, 64, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Kandouz, M.; Alachkar, A.; Zhang, L.; Dekhil, H.; Chehna, F.; Yasmeen, A.; Al Moustafa, A.E. Teucrium polium plant extract inhibits cell invasion and motility of human prostate cancer cells via the restoration of the E-cadherin/catenin complex. J. Ethnopharmacol. 2010, 129, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Movahedi, A.; Basir, R.; Rahmat, A.; Charaffedine, M.; Othman, F. Remarkable Anticancer Activity of Teucrium polium on Hepatocellular Carcinogenic Rats. Evid. Based Complement. Altern. Med. 2014, 2014, 726724. [Google Scholar] [CrossRef] [PubMed]
- Rehman, N.; Al-Sahai, J.M.S.; Hussain, J.; Khan, A.L.; Gilani, S.A.; Abbas, G.; Hussain, J.; Sabahi, J.N.; Al-Harassi, A. Phytochemical and pharmacological investigation of Teucrium muscatense. Int. J. Phytomed. 2016, 8, 567–579. [Google Scholar]
- Eloff, J.N. Which extractant should be used for the screening and isolation of antimicrobial components from plants? J. Ethnopharmacol. 1998, 60, 1–8. [Google Scholar] [CrossRef]
- Soule, H.D.; Vazguez, J.; Long, A.; Albert, S.; Brennan, M. A human cell line from a pleural effusion derived from a breast carcinoma. J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Scherer, W.F.; Syverton, J.T.; Gey, G.O. Studies on the propagation in vitro of poliomyelitis viruses. IV. Viral multiplication in a stable strain of human malignant epithelial cells (strain HeLa) derived from an epidermoid carcinoma of the cervix. J. Exp. Med. 1953, 97, 695–710. [Google Scholar] [CrossRef] [PubMed]
- Soule, H.D.; Maloney, T.M.; Wolman, S.R.; Peterson, W.D., Jr.; Brenz, R.; McGrath, C.M.; Russo, J.; Pauley, R.J.; Jones, R.F.; Brooks, S.C. Isolation and characterization of a spontaneously immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990, 50, 6075–6086. [Google Scholar] [PubMed]
- Cailleau, R.; Young, R.; Olive, M.; Reeves, W.J., Jr. Breast tumor cell lines from pleural effusions. J. Natl. Cancer Inst. 1974, 53, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Wahba, H.A.; El-Hadaad, H.A. Current approaches in treatment of triple-negative breast cancer. Cancer Biol. Med. 2015, 12, 106–116. [Google Scholar] [PubMed]
- Iqbal, J.; Abbasi, B.A.; Mahmood, T.; Kanwal, S.; Ali, B.; Shah, S.A.; Khalil, A.T. Plant-derived anticancer agents: A green anticancer approach. Asian Pac. J. Trop. Biomed. 2017, 7, 1129–1150. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Desouza, M.; Gunning, P.W.; Stehn, J.R. The actin cytoskeleton as a sensor and mediator of apoptosis. Bioarchitecture 2012, 2, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Stennicke, H.R.; Salvesen, G.S. Biochemical characteristics of caspases-3, -6, -7, and -8. J. Biol. Chem. 1997, 272, 25719–25723. [Google Scholar] [CrossRef] [PubMed]
- Suhail, M.M.; Wu, W.; Cao, A.; Mondalek, F.G.; Fung, K.M.; Shih, P.T.; Fang, Y.T.; Woolley, C.; Young, G.; Lin, H.K. Boswellia sacra essential oil induces tumor cell-specific apoptosis and suppresses tumor aggressiveness in cultured human breast cancer cells. BMC Complement. Altern. Med. 2011, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Janicke, R.U. MCF-7 breast carcinoma cells do not express caspase-3. Breast Cancer Res. Treat. 2009, 117, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Gali-Muhtasib, H.; Hmadi, R.; Kareh, M.; Tohme, R.; Darwiche, N. Cell death mechanisms of plant-derived anticancer drugs: Beyond apoptosis. Apoptosis 2015, 20, 1531–1562. [Google Scholar] [CrossRef] [PubMed]
- Wink, M.; Alfermann, A.W.; Frank, R.; Wetterauer, B.; Disti, M.; Windhovel, J.; Krohn, O.; Fuss, E.; garden, H.; Mohagheghzadeh, A.; et al. Sustainable bioproduction of phytochemicals by plant in vitro cultures: Anticancer agents. Plant Genet. Resour. 2005, 3, 90–100. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Ali, L.; Hussain, I.; Rizvi, T.S.; Khan, A.L.; Shaukat, S.; Al-Sahi, J.M.S.; Al-Harrasi, A.; Hussain, J. A New Irregular Trihydroxy Sesquiterpene from Teucrium mascatense. Helv. Chim. Acta 2015, 98, 1462–1465. [Google Scholar] [CrossRef]
- Stankovic, M.S.; Curcic, M.G.; Zizic, J.B.; Topuzovic, M.D.; Solujic, S.R.; Markovic, S.D. Teucrium plant species as natural sources of novel anticancer compounds: Antiproliferative, proapoptotic and antioxidant properties. Int. J. Mol. Sci. 2011, 12, 4190–4205. [Google Scholar] [CrossRef] [PubMed]
- Tafrihi, M.; Toosi, S.; Minaei, T.; Gohari, A.R.; Niknam, V.; Arab Najafi, S.M. Anticancer properties of Teucrium persicum in PC-3 prostate cancer cells. Asian Pac. J. Cancer Prev. 2014, 15, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Jaradat, N.; Al-Lahham, S.; Abualhasan, M.N.; Bakri, A.; Zaide, H.; Hammad, J.; Hussein, F.; Issa, L.; Mousa, A.; Speih, R. Chemical Constituents, Antioxidant, Cyclooxygenase Inhibitor, and Cytotoxic Activities of Teucrium pruinosum Boiss. Essential Oil. BioMed Res. Int. 2018, 2018, 4034689. [Google Scholar] [CrossRef] [PubMed]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Ncube, N.; Afolayan, A.; Okoh, A. Assessment techniques of antimicrobial properties of natural compounds of plant origin: Current methods and future trends. Afr. J. Biotechnol. 2008, 7, 1797–1806. [Google Scholar] [CrossRef]
- Chaves, A.L.; Vergara, C.E.; Mayer, J.E. Dichloromethane as an economic alternative to chloroform in the extraction of DNA from plant tissues. Plant Mol. Biol. Rep. 1995, 13, 18–25. [Google Scholar] [CrossRef]
- Al-Oqail, M.M.; Al-Sheddi, E.S.; Siddiqui, M.A.; Musarrat, J.; Al-Khedhairy, A.A.; Farshori, N.N. Anticancer Activity of Chloroform Extract and Sub-fractions of Nepeta deflersiana on Human Breast and Lung Cancer Cells: An In vitro Cytotoxicity Assessment. Pharmacogn. Mag. 2015, 11 (Suppl. 4), S598–S605. [Google Scholar] [PubMed]
- Kh, A.H.; Kh, S.; Yaseen, N. Evaluation the activity of chloroform extract of Solanum nigrum on non-Hodgkin lymphoma cell line (SR), rat embryo fibroblast cell line (REF) and human lymphocytes in vitro. World J. Pharm. Sci. 2015, 4, 14–22. [Google Scholar]
- Modzelewska, A.; Sur, S.; Kumar, S.K.; Khan, S.R. Sesquiterpenes: Natural products that decrease cancer growth. Curr. Med. Chem. Anticancer Agents 2005, 5, 477–499. [Google Scholar] [CrossRef] [PubMed]
- Bartikova, H.; Hanusova, V.; Skalova, L.; Ambroz, M.; Bousova, I. Antioxidant, pro-oxidant and other biological activities of sesquiterpenes. Curr. Top. Med. Chem. 2014, 14, 2478–2494. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Spiteller, M. Natural products containing ‘decalin’ motif in microorganisms. Nat. Prod. Rep. 2014, 31, 1175–1201. [Google Scholar] [CrossRef] [PubMed]
- Menichini, F.; Conforti, F.; Rigano, D.; Formisano, C.; Piozzi, F.; Senatore, F. Phytochemical composition, anti-inflammatory and antitumour activities of four Teucrium essential oils from Greece. Food Chem. 2009, 115, 679–686. [Google Scholar] [CrossRef]
- Guaman-Ortiz, L.M.; Orellana, M.I.; Ratovitski, E.A. Natural Compounds as Modulators of Non-apoptotic Cell Death in Cancer Cells. Curr. Genom. 2017, 18, 132–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Karantza, V. Autophagy as a therapeutic target in cancer. Cancer Biol. Ther. 2011, 11, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, A.; Wang, L. Bridging the divide: Preclinical research discrepancies between triple-negative breast cancer cell lines and patient tumors. Oncotarget 2017, 8, 113269–113281. [Google Scholar] [CrossRef] [PubMed]
- Suffredini, I.B.; Paciencia, M.L.; Frana, S.A.; Varella, A.D.; Younes, R.N. In vitro breast cancer cell lethality of Brazilian plant extracts. Pharmazie 2007, 62, 798–800. [Google Scholar] [PubMed]
- Fouche, G.; Cragg, G.M.; Pillay, P.; Kolesnikova, N.; Maharaj, V.J.; Senabe, J. In vitro anticancer screening of South African plants. J. Ethnopharmacol. 2008, 119, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. In Assay Guidance Manual; Sittampalam, G.S., Coussens, N.P., Brimacombe, K., Grossman, A., Arkin, M., Auld, D., Austin, C., Baell, J., Bejcek, B., Caaveiro, J.M.M., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
Sample Availability: Samples of TMDF may be available from the authors, if not exhausted. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract/Fractions | MCF-7 (24/72 HRS) Breast Cancer Cells (µg/mL) * | HeLa (24/72 HRS) Cervical Cancer Cells (µg/mL) * |
|---|---|---|
| TMHF | 24HR: 250 72HR: 125/250 | 24HR: X 72HR: 250 |
| TMDF | 24HR: 250 72HR: 125/250 | 24HR: X 72HR: 250 |
| TMEF | X | X |
| TMBF | X | X |
| TMME | 24HR: 250 72HR: 125/250 | 24HR: X 72HR: 250 |
| TMAF | X | X |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panicker, N.G.; Balhamar, S.O.M.S.; Akhlaq, S.; Qureshi, M.M.; Rizvi, T.S.; Al-Harrasi, A.; Hussain, J.; Mustafa, F. Identification and Characterization of the Caspase-Mediated Apoptotic Activity of Teucrium mascatense and an Isolated Compound in Human Cancer Cells. Molecules 2019, 24, 977. https://doi.org/10.3390/molecules24050977
Panicker NG, Balhamar SOMS, Akhlaq S, Qureshi MM, Rizvi TS, Al-Harrasi A, Hussain J, Mustafa F. Identification and Characterization of the Caspase-Mediated Apoptotic Activity of Teucrium mascatense and an Isolated Compound in Human Cancer Cells. Molecules. 2019; 24(5):977. https://doi.org/10.3390/molecules24050977
Chicago/Turabian StylePanicker, Neena Gopinathan, Sameera Omar Mohammed Saeed Balhamar, Shaima Akhlaq, Mohammed Mansour Qureshi, Tania Shamim Rizvi, Ahmed Al-Harrasi, Javid Hussain, and Farah Mustafa. 2019. "Identification and Characterization of the Caspase-Mediated Apoptotic Activity of Teucrium mascatense and an Isolated Compound in Human Cancer Cells" Molecules 24, no. 5: 977. https://doi.org/10.3390/molecules24050977
APA StylePanicker, N. G., Balhamar, S. O. M. S., Akhlaq, S., Qureshi, M. M., Rizvi, T. S., Al-Harrasi, A., Hussain, J., & Mustafa, F. (2019). Identification and Characterization of the Caspase-Mediated Apoptotic Activity of Teucrium mascatense and an Isolated Compound in Human Cancer Cells. Molecules, 24(5), 977. https://doi.org/10.3390/molecules24050977