
Profiling Metabolites and Biological Activities of Sugarcane (Saccharum officinarum Linn.) Juice and its Product Molasses via a Multiplex Metabolomics Approach

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results and Discussion
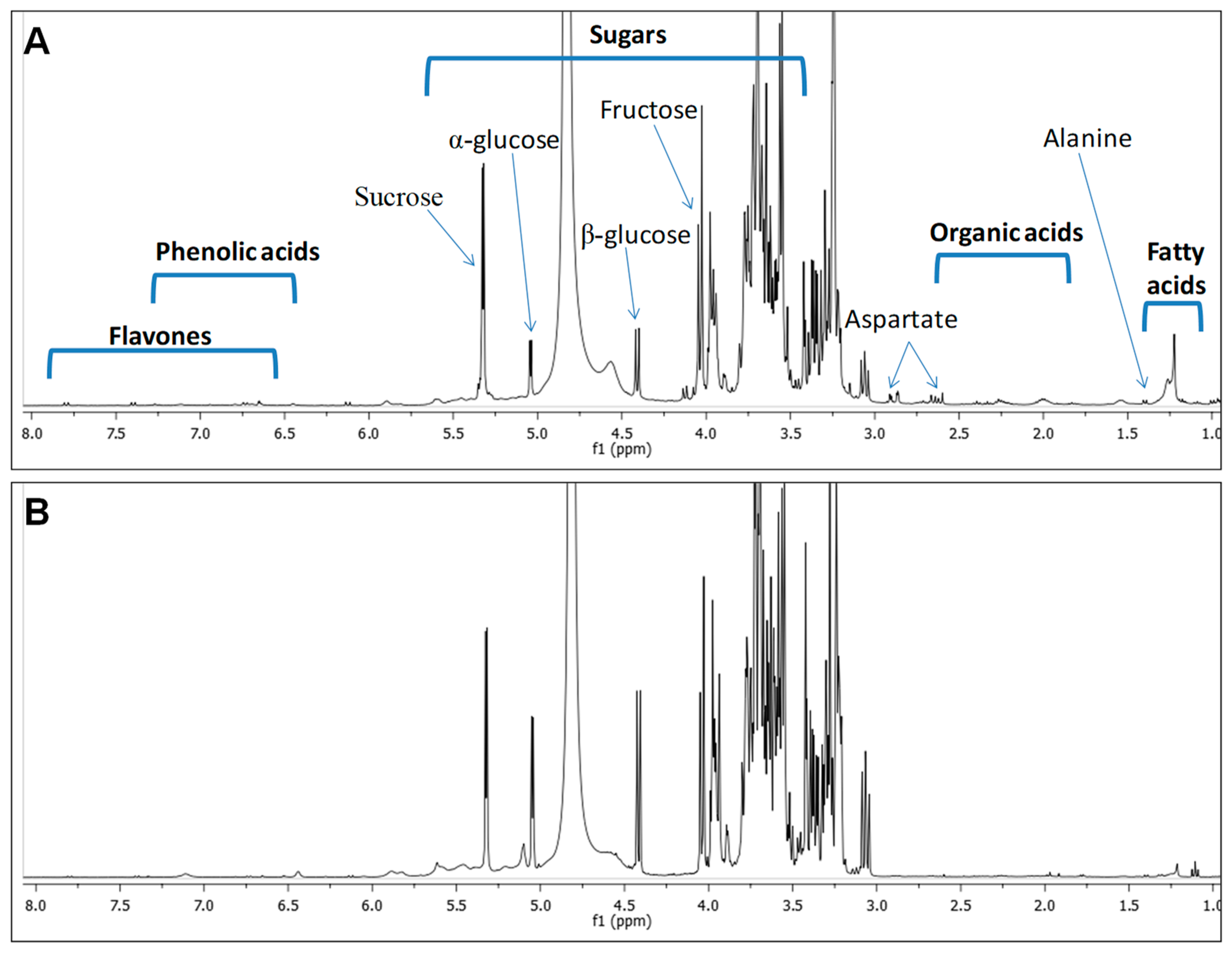
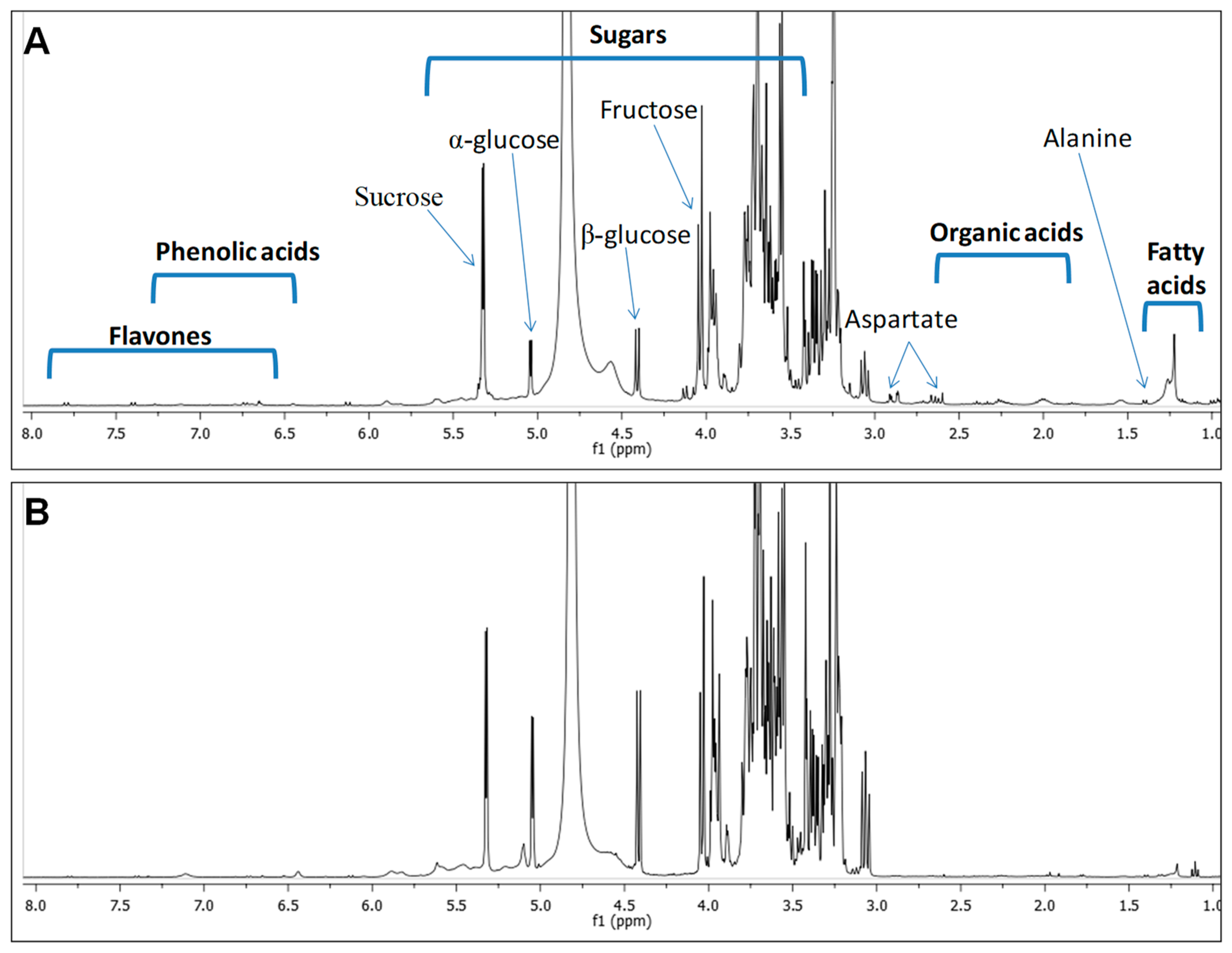
2.1. H-NMR Spectroscopy Metabolite Fingerprinting and Quantification
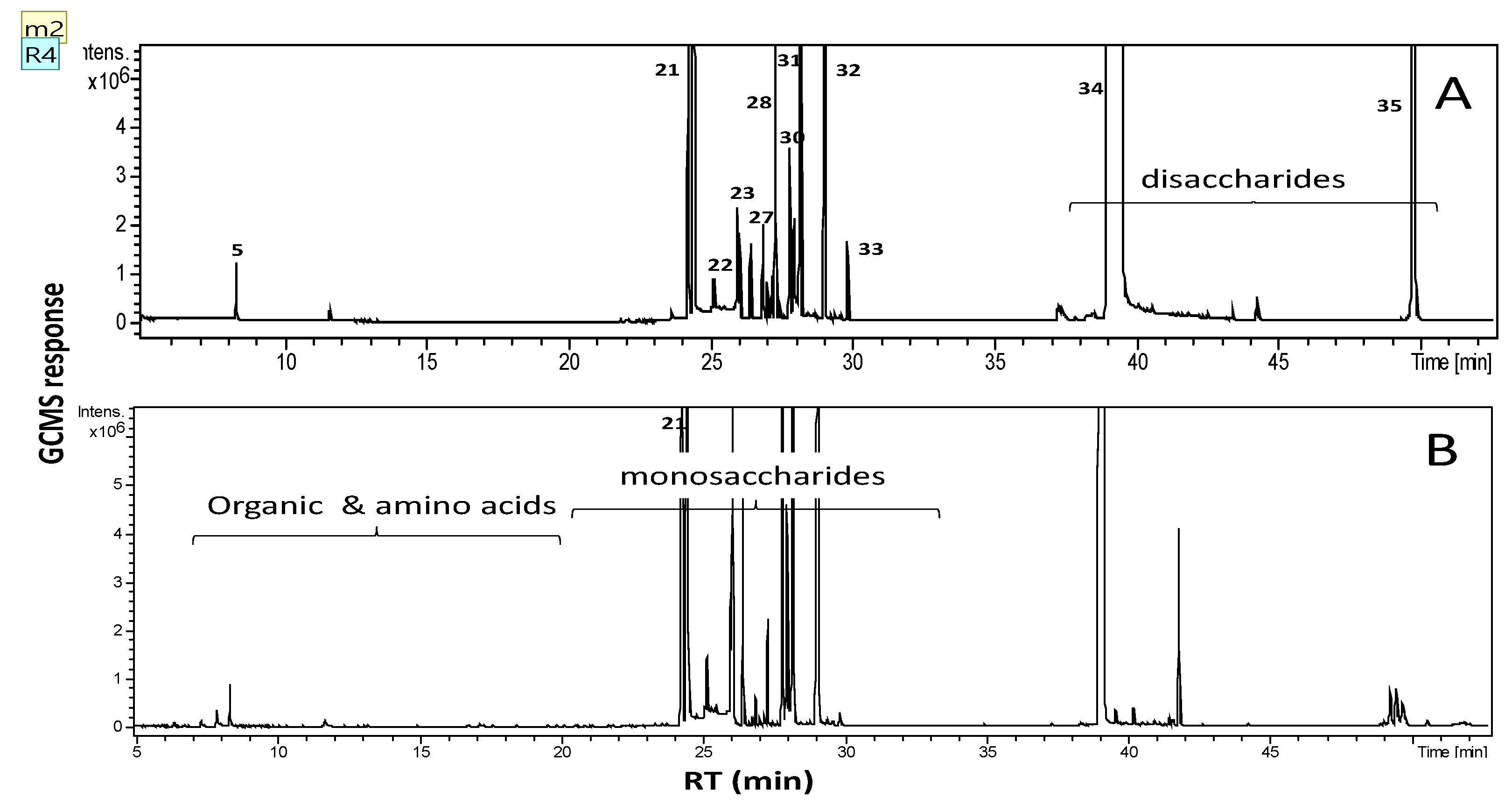
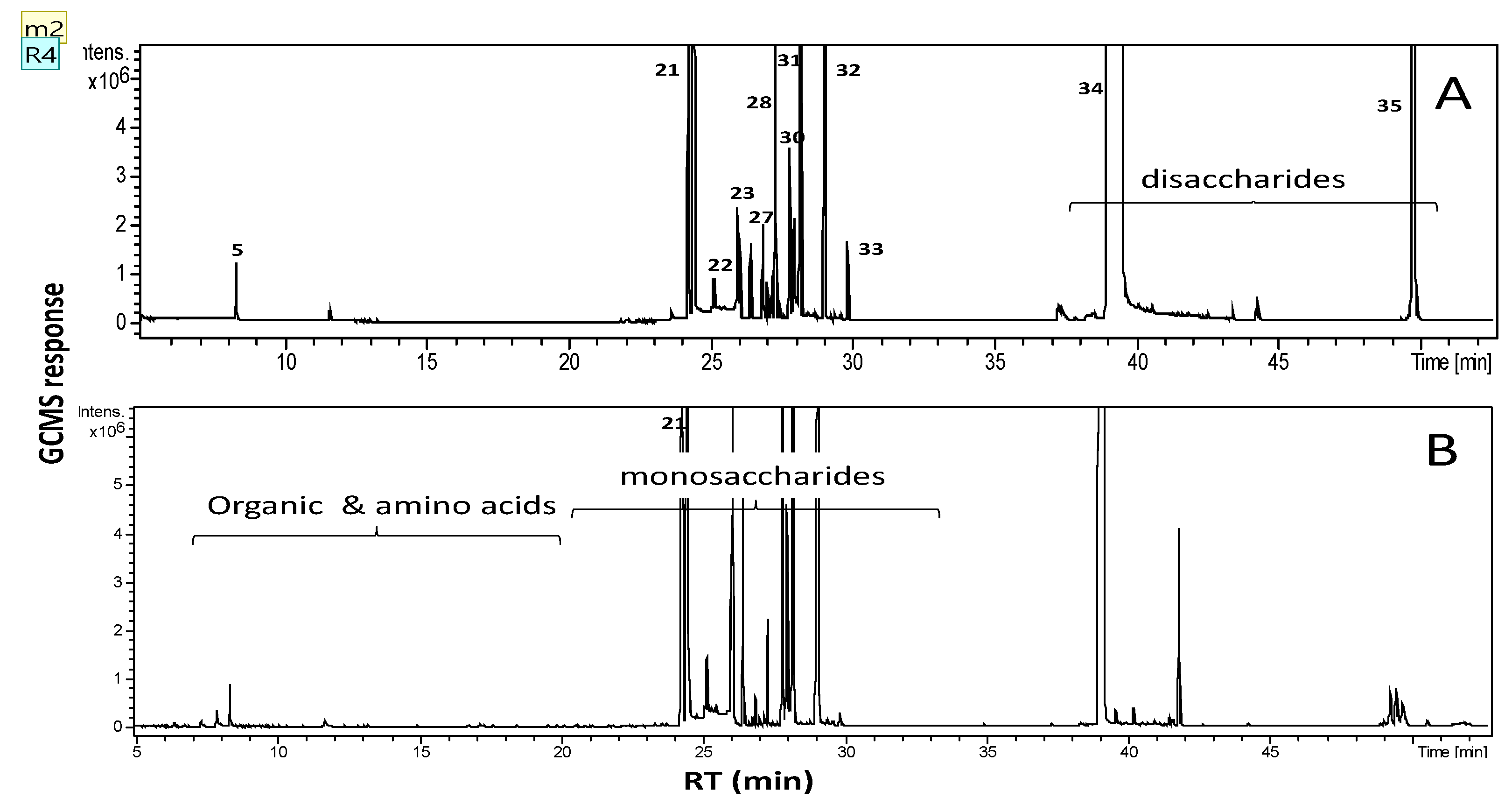
2.2. Primary Metabolite Profiling of Sugars, Amino Acids and Organic Acids via GC/MS
2.3. Secondary Metabolites Profiling via UPLC/MS
2.3.1. Flavonoids
2.3.2. Fatty Acids and Sterols
2.3.3. Miscellaneous
2.4. TPC and TFC in Sugarcane Juice and Molasses
2.5. In vitro DPPH, ABTS and FRAP Antioxidant Assays of Sugarcane and Molasses
2.6. α-Amylase & α-Glucosidase Inhibition Assays
3. Materials and Methods
3.1. Extraction and Sample Preparation
3.2. Chemicals and Reagents
3.3. NMR Analysis of Sugarcane Juice and Molasses
3.4. NMR Quantification
- MT—molecular weight of the target compound [g/mol];
- IT—relative integral value of the 1H-NMR signal of the target compound;
- ISt—relative integral value of the 1H-NMR signal of the standard compound;
- xSt—number of protons belonging to the 1H-NMR signal of the standard compound;
- xT—number of protons belonging to the 1H-NMR signal of the target compound;
- cSt—concentration of the standard compound in the solution used for 1H-NMR analysis [mmol/L];
- vSt—volume of the solution used for 1H NMR analysis [mL].
3.5. GC/MS Analysis of Silylated Primary Metabolites in Molasses and Sugarcane Juice
3.6. RP-UPLC/MS of the Secondary Metabolites in Molasses and Sugarcane
3.7. TPC
3.8. TFC
3.9. DPPH Radical Scavenging Assay
3.10. TEAC Assay
3.11. FRAP Antioxidant Capacity Assay
3.12. α-Amylase Inhibition Assay
3.13. α-Glucosidase Inhibition Assay
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, A.; Lal, U.R.; Mukhtar, H.M.; Singh, P.S.; Shah, G.; Dhawan, R.K. Phytochemical profile of sugarcane and its potential health aspects. Pharmacogn. Rev. 2015, 9, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Luo, Z.; Zhang, Y.; Zhong, Z.; Lu, B. Phytochemical contents and antioxidant capacities of different parts of two sugarcane (Saccharum officinarum L.) cultivars. Food Chem. 2014, 151, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, F.G.C.; Soares, A.K.L.; Santaella, S.T.; Alexandre e Silva, L.M.; Canuto, K.M.; Cáceres, C.A.; Rosa, M.D.F.; Feitosa, J.P.D.A.; Leitão, R.C. Optimization of the acetosolv extraction of lignin from sugarcane bagasse for phenolic resin production. Ind. Crops Prod. 2017, 96, 80–90. [Google Scholar] [CrossRef]
- Costa, M.; Di Stasi, L.C.; Kirizawa, M.; Mendacolli, S.L.; Gomes, C.; Trolin, G. Screening in mice of some medicinal plants used for analgesic purposes in the state of Sao Paulo. Part II. J. Ethnopharmacol. 1989, 27, 25–33. [Google Scholar] [CrossRef]
- Arruzazabala, M.L.; Carbajal, D.; Mas, R.; Molina, V.; Valdes, S.; Laguna, A. Cholesterol-lowering effects of policosanol in rabbits. Biol. Res. 1994, 27, 205–208. [Google Scholar] [PubMed]
- Ribeiro Rde, A.; Fiuza de Melo, M.M.; De Barros, F.; Gomes, C.; Trolin, G. Acute antihypertensive effect in conscious rats produced by some medicinal plants used in the state of Sao Paulo. J. Ethnopharmacol. 1986, 15, 261–269. [Google Scholar] [CrossRef]
- Ledon, N.; Casaco, A.; Rodriguez, V.; Cruz, J.; Gonzalez, R.; Tolon, Z.; Cano, M.; Rojas, E. Anti-inflammatory and analgesic effects of a mixture of fatty acids isolated and purified from sugar cane wax oil. Planta Med. 2003, 69, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Arruzazabala, M.L.; Noa, M.; Menendez, R.; Mas, R.; Carbajal, D.; Valdes, S.; Molina, V. Protective effect of policosanol on atherosclerotic lesions in rabbits with exogenous hypercholesterolemia. Braz. J. Med. Biol. Res. 2000, 33, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Molina, V.; Arruzazabala, M.L.; Carbajal, D.; Mas, R.; Valdes, S. Antiplatelet and antithrombotic effect of D-003. Pharmacol. Res. 2000, 42, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Colombo, R.; Lancas, F.M.; Yariwake, J.H. Determination of flavonoids in cultivated sugarcane leaves, bagasse, juice and in transgenic sugarcane by liquid chromatography-UV detection. J. Chromatogr. A 2006, 1103, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Colombo, R.; Yariwake, J.H.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. On-line identification of sugarcane (Saccharum officinarum L.) methoxyflavones by liquid chromatography-UV detection using post-column derivatization and liquid chromatography-mass spectrometry. J. Chromatogr. A 2005, 1082, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Colombo, R.; Yariwake, J.H.; McCullagh, M. Study of C- and O-glycosylflavones in sugarcane extracts using liquid chromatography: Exact mass measurement mass spectrometry. J. Braz. Chem. Soc. 2008, 19, 483–490. [Google Scholar] [CrossRef]
- Guimaraes, C.M.; Giao, M.S.; Martinez, S.S.; Pintado, A.I.; Pintado, M.E.; Bento, L.S.; Malcata, F.X. Antioxidant activity of sugar molasses, including protective effect against DNA oxidative damage. J. Food Sci. 2007, 72, C039–C043. [Google Scholar] [CrossRef] [PubMed]
- Takara, K.; Otsuka, K.; Wada, K.; Iwasaki, H.; Yamashita, M. 1,1-Diphenyl-2-picrylhydrazyl Radical Scavenging Activity and Tyrosinase Inhibitory Effects of Constituents of Sugarcane Molasses. Biosci. Biotechnol. Biochem. 2007, 71, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Payet, B.; Shum Cheong Sing, A.; Smadja, J. Comparison of the concentrations of phenolic constituents in cane sugar manufacturing products with their antioxidant activities. J. Agric. Food Chem. 2006, 54, 7270–7276. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Tang, Q.; Fu, X.; Yu, S.; Wu, S.; Chen, M. Preparation of antioxidants from sugarcane molasses. Food Chem. 2014, 152, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Cevallos-Cevallos, J.M.; Reyes-De-Corcuera, J.I. Metabolomics in food science. Adv. Food Nutr. Res. 2012, 67, 1–24. [Google Scholar] [PubMed]
- Farag, M.A.; El-Ahmady, S.H.; Elian, F.S.; Wessjohann, L.A. Metabolomics driven analysis of artichoke leaf and its commercial products via UHPLC-q-TOF-MS and chemometrics. Phytochemistry 2013, 95, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Gad, H.A.; Heiss, A.G.; Wessjohann, L.A. Metabolomics driven analysis of six Nigella species seeds via UPLC-qTOF-MS and GC-MS coupled to chemometrics. Food Chem. 2014, 151, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, I.; Thapaliya, M.; Boroujerdi, A.; Chowdhury, K. NMR-based metabolomics study of the biochemical relationship between sugarcane callus tissues and their respective nutrient culture media. Anal. Bioanal. Chem. 2014, 406, 5997–6005. [Google Scholar] [CrossRef] [PubMed]
- Connor, S.C.; Wu, W.; Sweatman, B.C.; Manini, J.; Haselden, J.N.; Crowther, D.J.; Waterfield, C.J. Effects of feeding and body weight loss on the 1H-NMR-based urine metabolic profiles of male Wistar Han rats: Implications for biomarker discovery. Biomarkers 2004, 9, 156–179. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, I.D.; Baker, J.M.; Ward, J.L.; Beale, M.H.; Creste, S.; Cavalheiro, A.J. Metabolite Profiling of Sugarcane Genotypes and Identification of Flavonoid Glycosides and Phenolic Acids. J. Agric. Food Chem. 2016, 64, 4198–4206. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, I.; Shrestha, B.; Boroujerdi, A.; Chowdhury, K. NMR-based metabolomics profile comparisons to distinguish between embryogenic and non-embryogenic callus tissue of sugarcane at the biochemical level. In Vitro Cell. Dev. Biol. Plant 2015, 51, 340–349. [Google Scholar] [CrossRef]
- Aquino, F.W.; Boso, L.M.; Cardoso, D.R.; Franco, D.W. Amino acids profile of sugar cane spirit (cachaca), rum, and whisky. Food Chem. 2008, 108, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [PubMed]
- Alves, V.G.; Souza, A.G.; Chiavelli, L.U.; Ruiz, A.L.; Carvalho, J.E.; Pomini, A.M.; Silva, C.C. Phenolic compounds and anticancer activity of commercial sugarcane cultivated in Brazil. An. Acad. Bras. Cienc. 2016, 88, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Verschooten, L.; Smaers, K.; Van Kelst, S.; Proby, C.; Maes, D.; Declercq, L.; Agostinis, P.; Garmyn, M. The Flavonoid Luteolin Increases the Resistance of Normal, but Not Malignant Keratinocytes, Against UVB-Induced Apoptosis. J. Investig. Dermatol. 2010, 130, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.H. The strengths and weaknesses of NMR spectroscopy and mass spectrometry with particular focus on metabolomics research. Methods Mol. Biol. 2015, 1277, 161–193. [Google Scholar] [PubMed]
- Espinosa, I.; Fogelfeld, L. Tagatose: From a sweetener to a new diabetic medication? Expert Opin. Investig. Drugs 2010, 19, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Yamaguchi, F.; Matsunaga, T.; Hirata, Y.; Kamitori, K.; Dong, Y.; Sui, L.; Tsukamoto, I.; Ueno, M.; Tokuda, M. Rare sugar d-psicose protects pancreas beta-islets and thus improves insulin resistance in OLETF rats. Biochem. Biophys. Res. Commun. 2012, 425, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Kishimoto, Y.; Yoshikawa, Y.; Hayashi, N.; Okuma, K.; Tohi, M.; Yagi, K.; Matsuo, T.; Izumori, K. Acute d-psicose administration decreases the glycemic responses to an oral maltodextrin tolerance test in normal adults. J. Nutr. Sci. Vitaminol. 2008, 54, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Yamaguchi, F.; Hirose, K.; Matsunaga, T.; Sui, L.; Hirata, Y.; Noguchi, C.; Katagi, A.; Kamitori, K.; Dong, Y.; et al. Rare sugar d-psicose prevents progression and development of diabetes in T2DM model Otsuka Long-Evans Tokushima Fatty rats. Drug Des. Dev. Ther. 2015, 9, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Montoya, G.; Londono, J.; Cortes, P.; Izquierdo, O. Quantitation of trans-aconitic acid in different stages of the sugar-manufacturing process. J. Agric. Food Chem. 2014, 62, 8314–8318. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Mendes, P.; Dixon, R.A. Plant metabolomics: Large-scale phytochemistry in the functional genomics era. Phytochemistry 2003, 62, 817–836. [Google Scholar] [CrossRef]
- Bouslimani, A.; Sanchez, L.M.; Garg, N.; Dorrestein, P.C. Mass spectrometry of natural products: Current, emerging and future technologies. Nat. Prod. Rep. 2014, 31, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Colombo, R.; Yariwake, J.H.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. On-line identification of minor flavones from sugarcane juice by LC/UV/MS and post-column derivatization. J. Braz. Chem. Soc. 2009, 20, 1574–1579. [Google Scholar] [CrossRef]
- Colombo, R.; Yariwake, J.H.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. On-line identification of further flavone C- and O-glycosides from sugarcane (Saccharum officinarum L., Gramineae) by HPLC-UV-MS. Phytochem. Anal. 2006, 17, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Takara, K.; Ushijima, K.; Wada, K.; Iwasaki, H.; Yamashita, M. Phenolic Compounds from Sugarcane Molasses Possessing Antibacterial Activity against Cariogenic Bacteria. J. Oleo Sci. 2007, 56, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Indrianingsih, A.W.; Tachibana, S.; Itoh, K. In Vitro Evaluation of Antioxidant and α-Glucosidase Inhibitory Assay of Several Tropical and Subtropical Plants. Procedia Environ. Sci. 2015, 28, 639–648. [Google Scholar] [CrossRef]
- Sun, J.; He, X.M.; Zhao, M.M.; Li, L.; Li, C.B.; Dong, Y. Antioxidant and nitrite-scavenging capacities of phenolic compounds from sugarcane (Saccharum officinarum L.) tops. Molecules 2014, 19, 13147–13160. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.; Afzal Qamar, M.; Bokhari, T.H.; Abbas, M.; Hussain, F.; Masood, N.; Keshavarzi, A.; Qureshi, N.; Nazir, A. Total phenolic, chromium contents and antioxidant activity of raw and processed sugars. Inf. Process. Agric. 2017, 4, 83–89. [Google Scholar] [CrossRef]
- Mabry, T.J.; Liu, Y.-L.; Pearce, J.; Dellamonica, G.; Chopin, J.; Markham, K.R.; Paton, N.H.; Smith, P. New Flavonoids from Sugarcane (Saccharum). J. Nat. Prod. 1984, 47, 127–130. [Google Scholar] [CrossRef]
- Eberhardt, M.V.; Lee, C.Y.; Liu, R.H. Antioxidant activity of fresh apples. Nature 2000, 405, 903–904. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, M.J.C.; Price, K.R. Analytical problems in the study of flavonoid compounds in onions. Food Chem. 1996, 57, 113–117. [Google Scholar] [CrossRef]
- Willcox, J.K.; Catignani, G.L.; Lazarus, S. Tomatoes and Cardiovascular Health. Crit. Rev. Food Sci. Nutr. 2003, 43, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, X.; Tan, J.; Wang, B. Influence of harvest season on antioxidant activity and constituents of rabbiteye blueberry (Vaccinium ashei) leaves. J. Agric. Food Chem. 2013, 61, 11477–11483. [Google Scholar] [CrossRef] [PubMed]
- Cholbi, M.R.; Paya, M.; Alcaraz, M.J. Inhibitory effects of phenolic compounds on CCl4-induced microsomal lipid peroxidation. Experientia 1991, 47, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Said, O.; Fulder, S.; Khalil, K.; Azaizeh, H.; Kassis, E.; Saad, B. Maintaining a physiological blood glucose level with ‘glucolevel’, a combination of four anti-diabetes plants used in the traditional Arab herbal medicine. Evid. Based Complement. Alternat. Med. 2008, 5, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of alpha-glucosidase, alpha-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kwon, C.S.; Son, K.H. Inhibition of alpha-glucosidase and amylase by luteolin, a flavonoid. Biosci. Biotechnol. Biochem. 2000, 64, 2458–2461. [Google Scholar] [CrossRef] [PubMed]
- Custodio, L.; Patarra, J.; Albericio, F.; Neng, N.R.; Nogueira, J.M.; Romano, A. In vitro antioxidant and inhibitory activity of water decoctions of carob tree (Ceratonia siliqua L.) on cholinesterases, alpha-amylase and alpha-glucosidase. Nat. Prod. Res. 2015, 29, 2155–2159. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.; Yu, S.; Zeng, F.; Wu, X. Phenolics Content and Inhibitory Effect of Sugarcane Molasses on α-Glucosidase and α-Amylase In Vitro. Sugar Tech 2016, 18, 333–339. [Google Scholar] [CrossRef]
- Farag, M.A.; Porzel, A.; Mahrous, E.A.; El-Massry, M.M.; Wessjohann, L.A. Integrated comparative metabolite profiling via MS and NMR techniques for Senna drug quality control analysis. Anal. Bioanal. Chem. 2015, 407, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
- Farag, M.A.; Porzel, A.; Wessjohann, L.A. Comparative metabolite profiling and fingerprinting of medicinal licorice roots using a multiplex approach of GC-MS, LC-MS and 1D NMR techniques. Phytochemistry 2012, 76, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Mocan, A.; Zengin, G.; Crisan, G.; Mollica, A. Enzymatic assays and molecular modeling studies of Schisandra chinensis lignans and phenolics from fruit and leaf extracts. J. Enzyme Inhib. Med. Chem. 2016, 31 (Suppl. 4), 200–210. [Google Scholar] [CrossRef] [PubMed]
- Mocan, A.; Vodnar, D.C.; Vlase, L.; Crisan, O.; Gheldiu, A.M.; Crisan, G. Phytochemical Characterization of Veronica officinalis L. V. teucrium L. and V. orchidea Crantz from Romania and Their Antioxidant and Antimicrobial Properties. Int. J. Mol. Sci. 2015, 16, 21109–21127. [Google Scholar] [PubMed]
- Reis, B.; Martins, M.; Barreto, B.; Milhazes, N.; Garrido, E.M.; Silva, P.; Garrido, J.; Borges, F. Structure-property-activity relationship of phenolic acids and derivatives. Protocatechuic acid alkyl esters. J. Agric. Food Chem. 2010, 58, 6986–6993. [Google Scholar] [CrossRef] [PubMed]
- Damiano, S.; Forino, M.; De, A.; Vitali, L.A.; Lupidi, G.; Taglialatela-Scafati, O. Antioxidant and antibiofilm activities of secondary metabolites from Ziziphus jujuba leaves used for infusion preparation. Food Chem. 2017, 230, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Gowri, P.M.; Tiwari, A.K.; Ali, A.Z.; Rao, J.M. Inhibition of alpha-glucosidase and amylase by bartogenic acid isolated from Barringtonia racemosa Roxb. seeds. Phytother. Res. 2007, 21, 796–799. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |

{kind=link}
{kind=link}
{kind=link}
| Class | Metabolite | δ1H (ppm) and Multiplicity | Assignments | Molasses | Sugarcane |
|---|---|---|---|---|---|
| Amino acids | Valine | 1.00 (d, J = 7.1 Hz) | CH3-4,5 | 0.87 | 0.61 |
| Alanine | 1.41 (d, J = 7.2 Hz); 3.23 (d, J = 1.7 Hz) | CH3-3; CH-2 | 1.67 | 1.0 | |
| Aspartate | 2.89 (dd, J = 16.9, 3.4 Hz); 2.63 (dd, J = 17.1, 9.5 Hz) | CH2-3 | - | 6.44 | |
| Organic acids | Lactic acid | 1.33 (d, J = 6.9 Hz) | CH3-3 | 1.54 | - |
| Malic acid | 2.28 (dd, J = 13.5, 1.7 Hz) | CH2-3 | - | 3.45 | |
| Formic acid | 8.31 (s) | HCOOH | 0.94 | 0.4 | |
| Sugars | β-Glucose | 4.42 (d, J = 7.8 Hz); 3.11 (d, J = 1.8 Hz); 3.36 (m); 3.22 (d, J = 2.0 Hz); 3.24 (m) | CH-1; CH-2; CH-3; CH-4; CH-5 | 57.1 | 11.2 |
| α-Glucose | 5.12 (d, J = 3.7 Hz); 3.61 (d, J = 2.9 Hz) | CH-1; CH-5 | 51.1 | 10.2 | |
| Fructose | 4.04 (d, J = 8.3 Hz); 3.56 (m) | CH-5 (α-Fructofuranose); CH2-6 (β-Fructofuranose) | + | + | |
| Sucrose | 5.39 (d, J = 3.8 Hz); 4.11 (d, J = 8.3 Hz) | CH-1 (glucose)CH-3 (fructose) | 126 | 67 | |
| Phenolic acids | Gentisic acid | 7.35 (d, J = 3.6 Hz); 6.75 (dd, J = 8.5, 2.3 Hz); 6.67 (d, J = 2.3 Hz) | CH-3; CH-5; CH-6 | 10.6 | 6.5 |
| Fatty acids | Fatty acids | 2.27 (m); 1.60 (m); 1.29 (m); 1.32 (m) | H-2; H-3; (CH2-)n ω-2; (CH2-)n ω-1 | + | + |
| Flavones | Luteolin | 6.73 (d, J = 2.3 Hz); 6.75 (d, J = 2.3 Hz) | CH-6; CH-8 | 8.5 | 6.2 |
| Apigenin | 7.41 (d, J = 8.5 Hz); 7.80 (d, J = 9.5 Hz); 6.66 (d, J = 2.3 Hz) | CH-3’ & 5’; CH-2’ & 6’; CH-6 | 11.2 | - |
| Name | Class | Rt (min) | RI | Sugarcane (%) | Molasses (%) |
|---|---|---|---|---|---|
| Ethylene glycol (2TMS) | Alcohol | 7.883 | 1122 | 0.34 | 0.67 |
| Phosphate (3TMS) | Inorganic acid | 11.569 | 1269 | 0.22 | 0.51 |
| Pipecolic acid (N,O-2 TMS) | Nitrogenous compound | 17.857 | 1517 | 0.03 | 0.08 |
| Phenylethanolamine (3TMS) | 20.473 | 1629 | 0.01 | 0.01 | |
| N,O-hydroxylamine (2TMS) | 8.346 | 1141 | 0.55 | 1.23 | |
| Aminoethanol, O,N,N-tris-TMS | 18.022 | 1525 | 0.02 | - | |
| Lactic acid (2TMS) | Organic acid | 6.394 | 1055 | 0.01 | 0.04 |
| Acetic acid (TMS) | 6.782 | 1073 | - | 0.01 | |
| Glyceric acid (3TMS) | 13.103 | 1329 | 0.02 | 0.03 | |
| Glyceric acid isomer (3TMS) | 13.738 | 1353 | 0.02 | 0.04 | |
| Acetoacetic acid (2TMS) | 16.671 | 1470 | 0.01 | 0.1 | |
| Malic acid (3TMS) | 17.078 | 1486 | 0.09 | 0.21 | |
| Itaconic acid (2TMS) | 19.971 | 1607 | 0.01 | 0.03 | |
| trans-Aconitic acid (3TMS) | 23.098 | 1747 | 0.07 | 0.12 | |
| Citric acid (4TMS) | 24.523 | 1813 | 0.07 | 0.01 | |
| α-d-Ribofuranoside, methyl (3TMS) | Sugars | 17.549 | 1505 | - | 0.01 |
| Threonic acid (4TMS) | 18.368 | 1539 | 0.01 | 0.01 | |
| Gluconic acid, 2-methoxime (4TMS) | 20.804 | 1644 | - | 0.01 | |
| Ribitol (5TMS) | 22.381 | 1714 | 0.01 | 0.02 | |
| Arabinofuranose (4TMS) | 23.605 | 1771 | 0.06 | 0.04 | |
| Tagatofuranose (5TMS) (isomer 2) | 24.216 | 1798 | 5.58 | 7.92 | |
| Mannopyranose, 6-deoxy-(4TMS) | 25.093 | 1841 | 0.20 | 1.05 | |
| Psicose (5TMS) | 25.927 | 1881 | 0.52 | 3.14 | |
| Mannopyranose (5TMS) | 26.069 | 1888 | 2.74 | 9.19 | |
| Glucitol (6TMS) | 26.721 | 1921 | 0.02 | 0.13 | |
| Fructose oxime (6TMS) | 26.966 | 1933 | 0.06 | 0.09 | |
| Fructose (5TMS) | 27.123 | 1941 | 0.72 | 0.89 | |
| Talose (5TMS) | 27.772 | 1975 | 4.28 | 5.94 | |
| Allopyranose (5TMS) | 27.786 | 1975 | 4.28 | 5.94 | |
| Galactofuranose (5 TMS) | 27.901 | 1981 | 1.44 | 1.65 | |
| Gluconic acid δ-lactone (4TMS) | 28.143 | 1993 | 1.49 | - | |
| Glucopyranose (5TMS) | 28.97 | 2035 | 6.30 | - | |
| Inositol (6TMS) myo- | 29.77 | 2076 | 0.26 | - | |
| Sucrose (8TMS) | 38.866 | 2539 | 60.69 | 58.8 | |
| Sucrose (8TMS) | 49.563 | 3083 | 5.25 | - | |
| Unknown | unknown | 24.402 | 1808 | 4.62 | 2.120 |
| Total | 100.00 | 100.00 |
| Peak No. | M + H | Rt. (sec.) | UV (nm) | Fragments MS/MS+ | Formula | Er. ppm | Identification | Class | Scj. | M |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 188.0705 | 144 | 220 | 163 | C11H10NO2+ | 0.3 | 1H-Indole-3-propenoic acid | A | + | - |
| 2 | 185.0801 | 186 | 221 | 163, 158, 141 | C9H13O4+ | 8.1 | Hydroxyphenyl)glycerol | GD | + | - |
| 3 | 257.1013 | 198 | 221 | 222, 195, 185, 163, 145 | C12H17O6+ | 2.7 | Unknown | - | + | |
| 4 | 227.0914 | 204 | 221,301 | 223, 215, 195, 185 | C11H15O5+ | 0.1 | Hydroxyphenyl)glyceryl acetate | GD | + | - |
| 5 | 249.0737 | 222 | 221 | 227, 209, 185, 163, 141 | C13H13O5+ | 8.4 | Unknown | + | - | |
| 6 | 565.1544 | 246 | 223,272,334 | 547, 475, 445, 409, 379 | C26H29O14+ | 1.5 | Schaftoside/ Isoschaftoside | Fl | + | + |
| 7 | 463.1234 | 252 | 223,271,345 | 445, 427, 409, 397, 343, 313 | C22H23O11+ | 0.1 | Diosmetin-8-C-glucoside | Fl | + | + |
| 8 | 433.1133 | 270 | 223,271,336 | 415, 397, 379, 367, 337, 313 | C21H21O10+ | 0.8 | Vitexin | Fl | + | + |
| 9 | 447.1294 | 276 | 222,272,332 | 297, 327 | C22H23O10+ | 1.8 | Swertisin | Fl | + | + |
| 10 | 477.1395 | 288 | 262,345 | 459, 441, 423, 411, 381, 357, 327, 315 | C23H25O11+ | 0.7 | 4’, 5’Dimethylluteolin-8-C-glucoside. | Fl | + | + |
| 11 | 377.1431 | 294 | 280 | C16H25O10+ | 3.0 | Guaiacylglycerol-O-hexoside | GD | - | + | |
| 12 | 653.1694 | 306 | 224,351 | 507, 331 | C29H33O17+ | 2.7 | Tricin7-O-deoxyhexosyl-glucuronide | Fl | + | + |
| 13 | 639.1903 | 312 | 222,346 | 331 | C29H35O16+ | 2.6 | Tricin-7-O-neohesperidoside | Fl | + | + |
| 14 | 353.23 | 444 | 223 | 325, 313, 303, 295, 277, 233, 203, 194 | C20H33O5+ | 6.4 | Unknown diterpene | DT | + | + |
| 15 | 311.2219 | 492 | 223 | 295, 254, 213, 202 | C18H31O4+ | 0.8 | Epoxy-hydroxy-octadecadienoic acid | HFA | + | - |
| 16 | 257.0792 | 504 | 223 | 230, 175 | C15H13O4+ | 6.5 | Liquiritigenin/ isoliquiritigenin | Fl | + | + |
| 17 | 271.0605 | 522 | 223 | 257, 236, 226 | C15H11O5+ | 1.4 | Apigenin | Fl | - | + |
| 18 | 272.2583 | 528 | 223 | 251, 226,209 | C16H34NO2+ | 0.4 | Hydroxy palmitic acyl amide | HFAA | + | - |
| 19 | 381.2608 | 570 | 224 | 365, 341, 325, 303 | C22H37O5+ | 7.2 | Unknown diterpene | DT | + | - |
| 20 | 295.2261 | 576 | 224 | 285, 277, 259, 249 | C18H31O3 | 2.3 | Hydroxy-octadecatrienoic acid | HFA | + | - |
| 21 | 345.2429 | 594 | 224 | 337, 325, 297, 285, 251, 226 | C22H33O3+ | 1.3 | Unknown diterpene | DT | - | + |
| 22 | 297.2424 | 600 | 224 | 277, 197 | C18H33O3+ | 0.1 | Hydroxy-octadecadienoic acid | HFA | - | + |
| 23 | 335.2197 | 630 | 224 | 325, 303, 295, 285 | C20H31O4+ | 5.8 | Unknown diterpene | DT | + | - |
| 24 | 335.219 | 678 | 224 | 325, 319, 313, 303, 295, 285, 249 | C20H31O4+ | 8.0 | Unknown diterpene | DT | - | + |
| 25 | 279.231 | 684 | 224 | 269, 249, 235, 226, 197 | C18H31O2+ | 3.1 | Octadecatrienoic acid | FA | + | - |
| 26 | 411.0939 | 738 | 224 | 389, 371, 365, 345, 331, 325, 317, 301 | C29H15O3+ | 8.6 | Unknown | - | + | |
| 27 | 341.2662 | 792 | 225 | 319, 297, 275, 253 | C20H37O4+ | 7.0 | Unknown diterpene | DT | - | + |
| 28 | 297.2398 | 792 | 225 | 285, 275, 267, 253 | C18H33O3+ | 9.0 | Hydroxy Octadecatrienoic acid | HFA | + | - |
| 29 | 355.2829 | 822 | 225 | 365, 355, 337, 325, 301, 279, 263, 245, 197 | C21H39O4+ | 3.9 | Octadecadienoyl glycerol | GD | - | + |
| 30 | 439.3556 | 834 | 225 | 399, 385, 355, 341, 325, 311, 301, 285 | C30H47O2+ | 3.4 | Unknown triterpene | TT | - | + |
| 31 | 277.2141 | 864 | 225 | 267, 261, 255, 243, 237 | C18H29O2+ | 7.6 | Octadecatetraenoic acid | FA | + | - |
| 32 | 353.2661 | 870 | 225 | 331,325,313, 301 | C21H37O4+ | 7.1 | Unknown | + | - | |
| 33 | 281.2471 | 876 | 225 | 281, 263, 245, 226 | C18H33O2+ | 1.5 | Linoleic acid | FA | + | + |
| 34 | 425.2868 | 888 | 226 | 365, 301, 285, 267, 241, 226. | C24H41O6+ | 6.9 | Tetrahydroxycholanoic acid | St | ||
| 35 | 408.3082 | 912 | 226 | 365, 325, 301, 282, 266, 250 | C24H42NO4+ | 6.4 | Cholamide | ST | + | + |
| 36 | 257.2464 | 918 | 226 | 239, 226 | C16H33O2+ | 4.3 | Palmitic acid | FA | + | + |
| 37 | 283.2638 | 936 | 226 | 265,257, 226 | C18H35O2+ | 2.3 | Oleic acid | FA | + | - |
| 38 | 397.3292 | 1008 | 227 | 375, 365, 355, 325, 312, 301 | C24H45O4+ | 5.2 | Unknown fatty acid | FA | + | + |
| 39 | 338.342 | 1014 | 227 | 321, 301, 259, 226 | C22H44NO+ | 0.6 | Docosenoic acyl amide | NAA | + | + |
| 40 | 413.2665 | 1020 | 227 | 393, 365, 338 | C26H37O4+ | 5.2 | Unknown fatty acid | FA | + | + |
| 41 | 353.3029 | 1026 | 331, 315, 301 | C22H41O3+ | 6.1 | Unknown | + | + | ||
| 42 | 469.3861 | 1068 | 430, 413, 365, 301 | C28H53O5+ | 5.6 | Unknown | - | + |
| Total Phenolics and Flavonoids Content | Antioxidant Capacity Assays | Enzyme Inhibitory Capacities | |||||
|---|---|---|---|---|---|---|---|
| TPC (mg GAE/100g Extract) | TFC (mg QE/100g Extract) | DPPH Scavenging (mgTE/g Extract) | ABTS Scavenging (mgTE/g Extract) | FRAP (mgTE/g Extract) | α-Amylase Inhibition (mmolACAE/g Extract) | α-Glucosidase Inhibiton (mmolACAE/g Extract) | |
| Sugarcane | 93 ± 2.9 | 259 ± 1.00 | 2.34 ± 0.66 | 9.65 ± 0.26 | 13.46 ± 0.72 | 0.5 ± 0.03 | 3.61 ± 0.70 |
| Molasses | 196 ± 2.1 | 297 ± 6.00 | 1.91 ± 0.19 | 11.82 ± 3.16 | 18.46 ± 0.90 | 0.43 ± 0.04 | 6.20 ± 0.31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, S.E.; El Gedaily, R.A.; Mocan, A.; Farag, M.A.; El-Seedi, H.R. Profiling Metabolites and Biological Activities of Sugarcane (Saccharum officinarum Linn.) Juice and its Product Molasses via a Multiplex Metabolomics Approach. Molecules 2019, 24, 934. https://doi.org/10.3390/molecules24050934
Ali SE, El Gedaily RA, Mocan A, Farag MA, El-Seedi HR. Profiling Metabolites and Biological Activities of Sugarcane (Saccharum officinarum Linn.) Juice and its Product Molasses via a Multiplex Metabolomics Approach. Molecules. 2019; 24(5):934. https://doi.org/10.3390/molecules24050934
Chicago/Turabian StyleAli, Sara E., Rania A. El Gedaily, Andrei Mocan, Mohamed A. Farag, and Hesham R. El-Seedi. 2019. "Profiling Metabolites and Biological Activities of Sugarcane (Saccharum officinarum Linn.) Juice and its Product Molasses via a Multiplex Metabolomics Approach" Molecules 24, no. 5: 934. https://doi.org/10.3390/molecules24050934
APA StyleAli, S. E., El Gedaily, R. A., Mocan, A., Farag, M. A., & El-Seedi, H. R. (2019). Profiling Metabolites and Biological Activities of Sugarcane (Saccharum officinarum Linn.) Juice and its Product Molasses via a Multiplex Metabolomics Approach. Molecules, 24(5), 934. https://doi.org/10.3390/molecules24050934