Tropane Alkaloids: Chemistry, Pharmacology, Biosynthesis and Production


Abstract
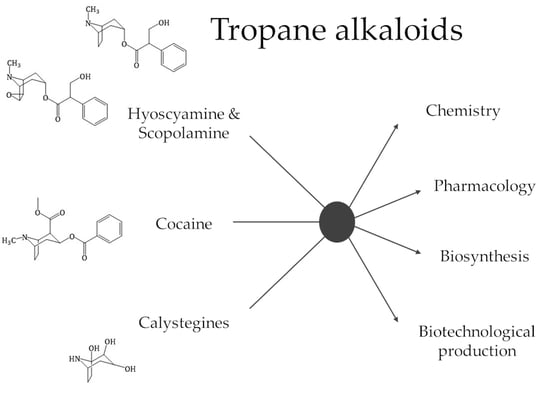
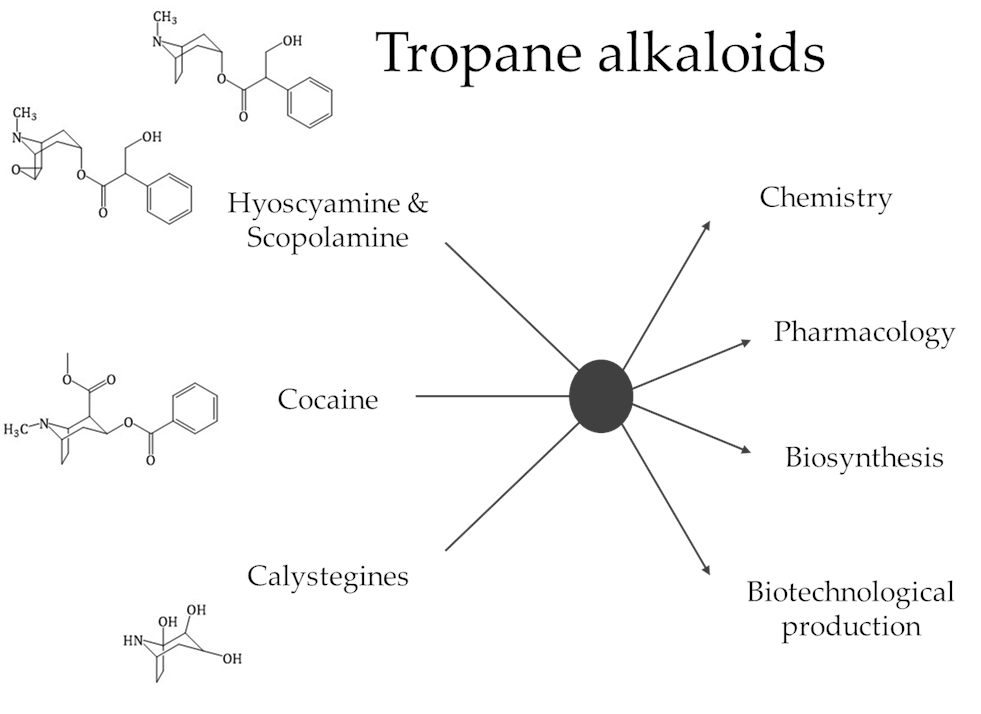
1. Introduction
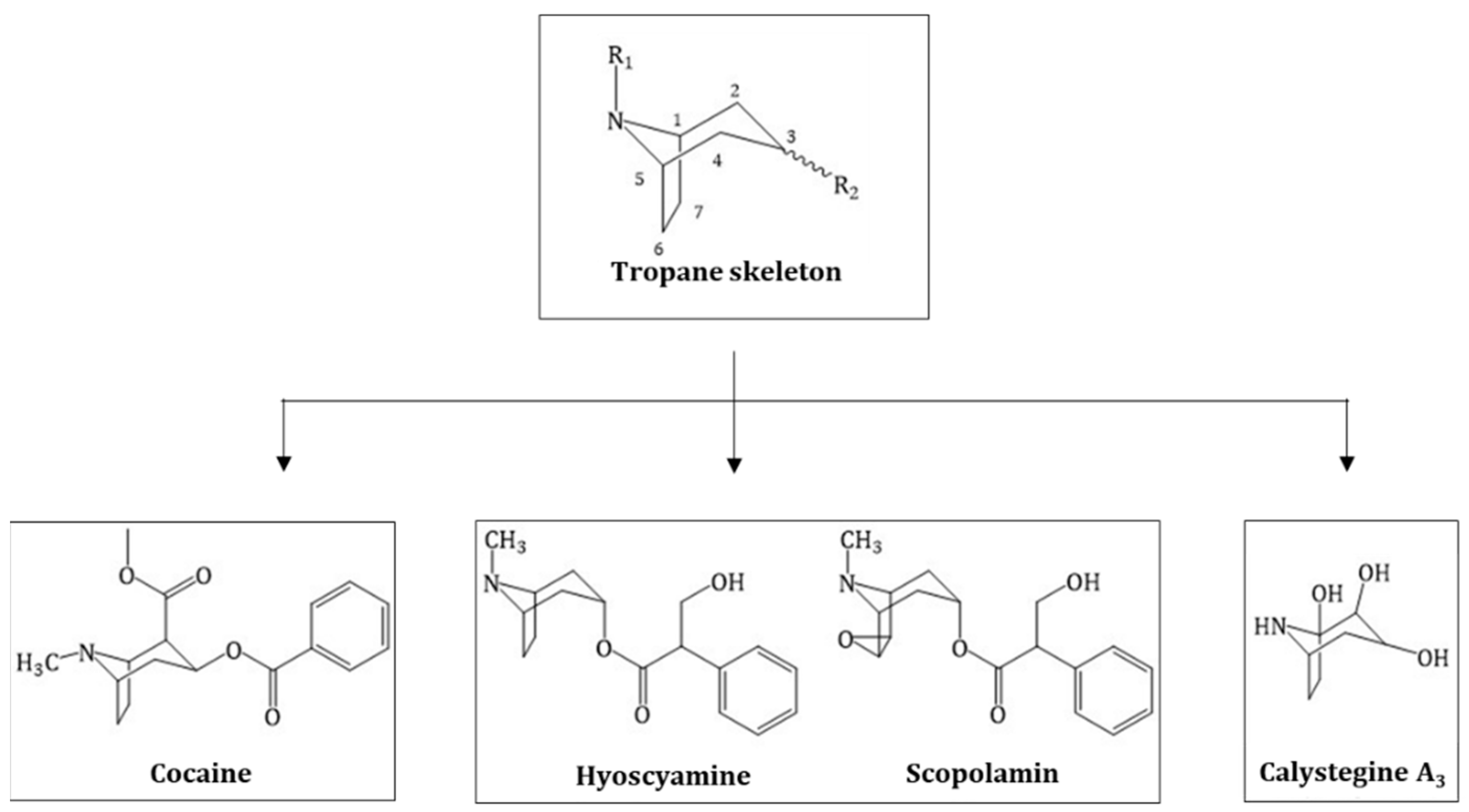
2. History and Chemical Elucidation of Tropane Alkaloids
2.1. Hyoscyamine and Scopolamine
2.2. Cocaine
2.3. Calystegines
3. Pharmacology of TAs and Their Role as Drug Lead Substances
3.1. Scopolamine, Hyoscyamine and Anisodamine and Their Derived Drugs
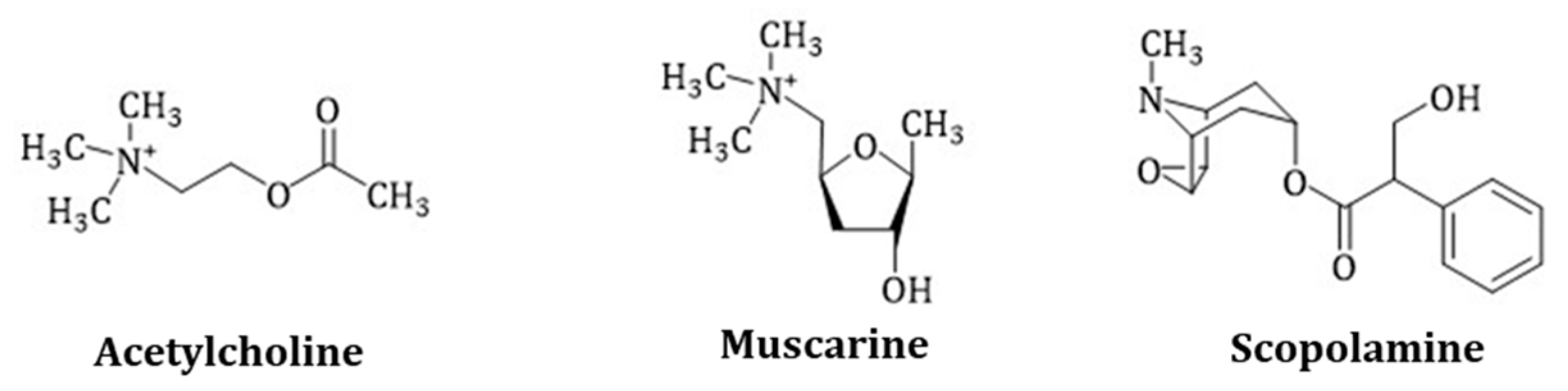
3.1.1. Scopolamine
3.1.2. Hyoscyamine and Atropine
3.1.3. Anisodamine
3.1.4. Homatropine, Cyclopentolate and Tropicamide
3.1.5. Trospium Chloride
3.1.6. Tropisetron
3.1.7. N-butylscopolamine
3.1.8. Tiotropium Bromide, Ipratropium Bromide and Oxitropium Bromide
3.1.9. Benzatropine
3.1.10. Scopolamine and Its Use as an Antidepressant
3.2. Cocaine Derived Drugs
3.3. Calystegine Derived Drugs
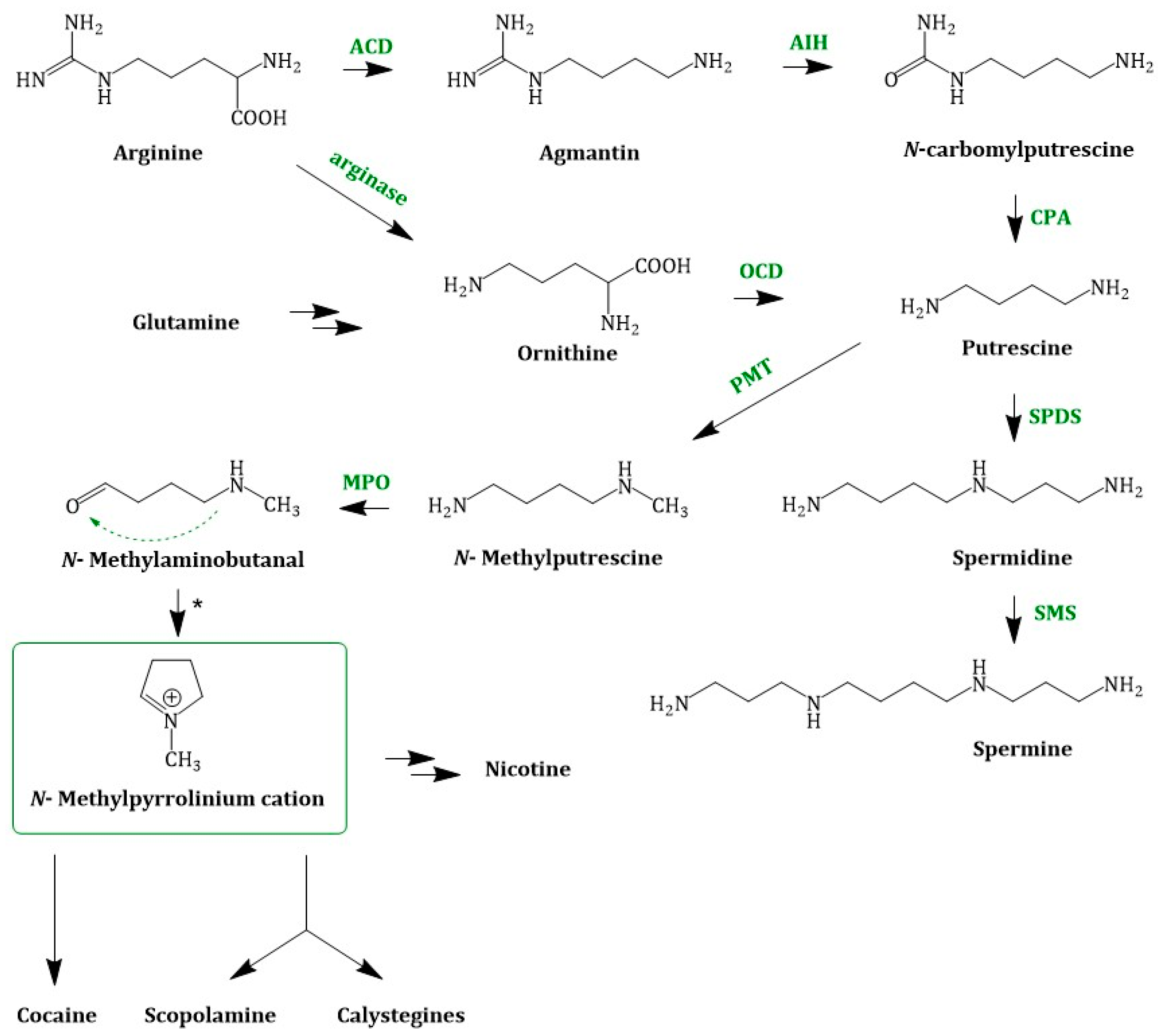
4. TA Biosynthesis In Planta
4.1. Early Steps in TA Biosynthesis—A United Way
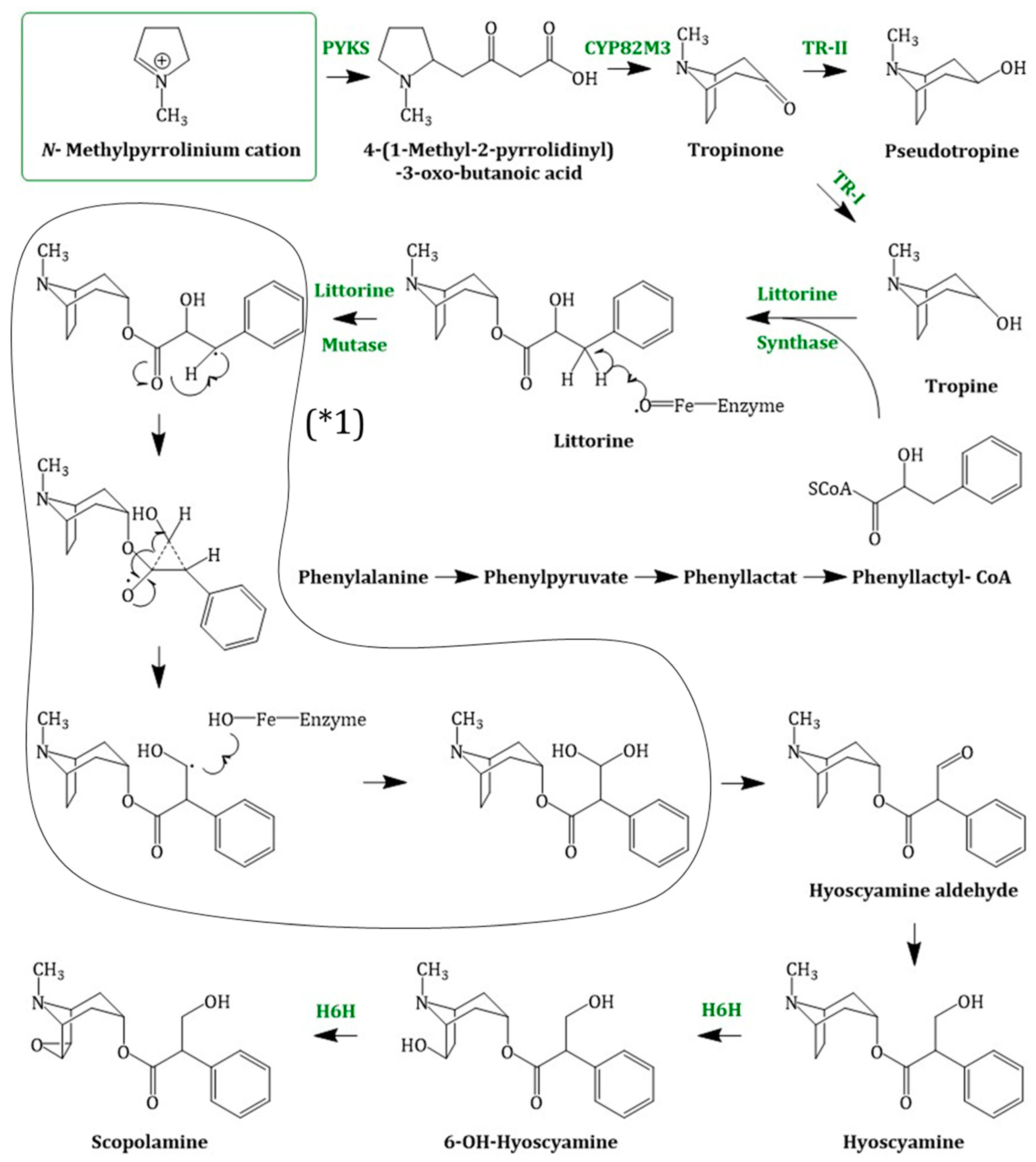
4.2. Hyoscyamine and Scopolamine Biosynthesis
4.2.1. Enzymes Involved in Scopolamine Formation and Their Regulation
Putrescine Methyltransferase
Tropinone Reductase I and Tropinone Reductase II
Putative Littorine Synthase
Littorine Mutase/Monooxygenase//CYP80F1
Hyoscyamine 6β-hydroxylase
4.2.2. Localization and Organization of Scopolamine Biosynthesis in Plants
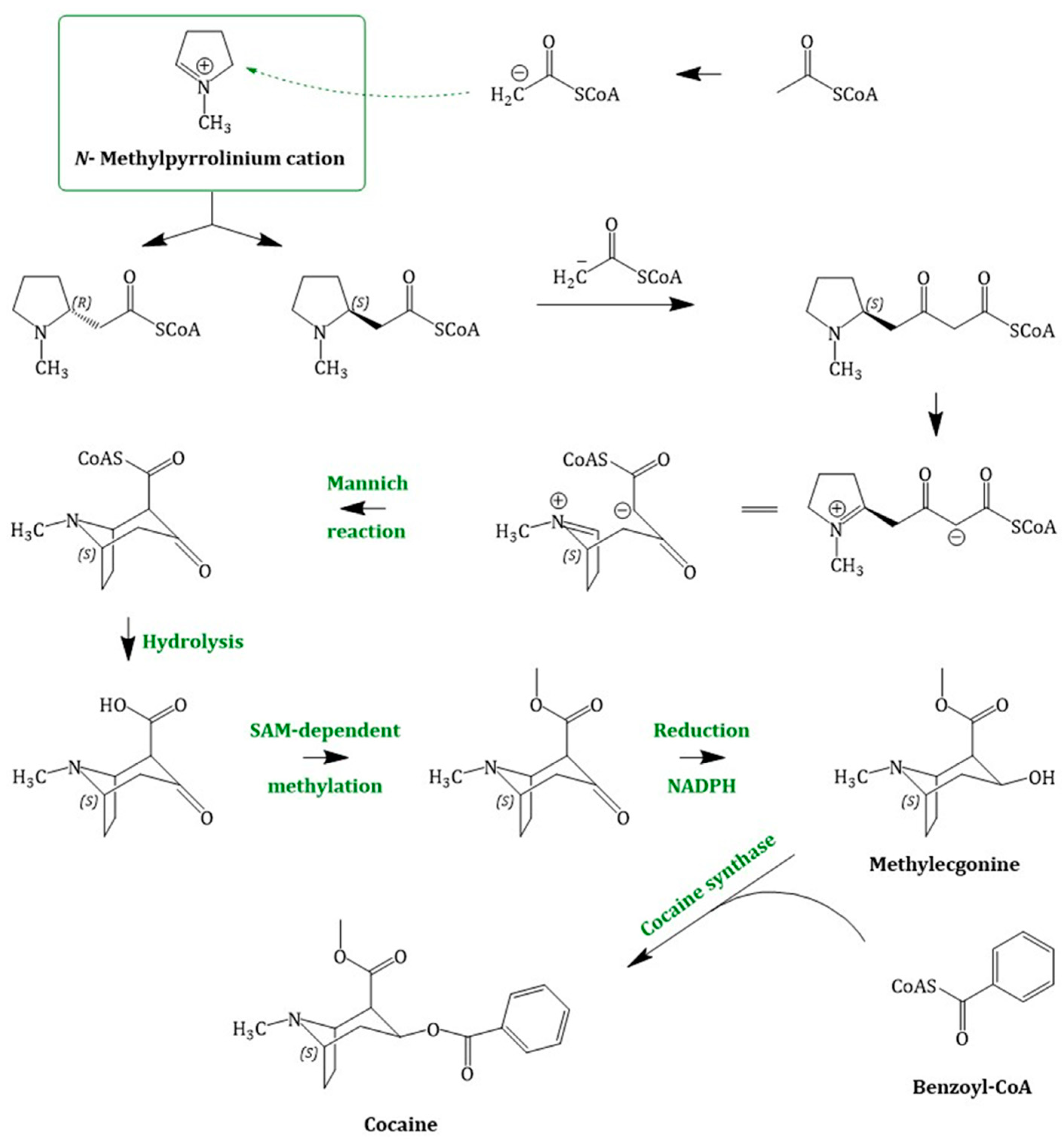
4.3. Cocaine Biosynthesis
4.4. Calystegine Biosynthesis
5. Biotechnological Approaches of Scopolamine Production and Alternative Methods of Raw Material Supply
5.1. Scopolamine Production in Cell Suspension and Hairy Root Cultures
5.2. Microbial Production of Scopolamine and Enzyme Engineering Approaches
5.3. Additional Methods of Scopolamine Production
6. Big Data—The Use of “Omics” in Plant Science and Pathway Elucidation
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grynkiewicz, G.; Gadzikowska, M. Tropane alkaloids as medicinally useful natural products and their synthetic derivatives as new drugs. Pharmacol. Rep. 2008, 60, 439–463. [Google Scholar] [PubMed]
- Dräger, B. Chemistry and Biology of calystegines. Nat. Prod. Rep. 2003, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Gadzikowska, M.; Grynkiewicz, G. Tropane alkaloids in pharmaceutical and phytochemical analysis. Acta Pol. Pharm. 2001, 58, 481–492. [Google Scholar]
- Ziegler, J.; Facchini, P.J. Alkaloid biosynthesis: Metabolism and trafficking. Annu. Rev. Plant Biol. 2008, 59, 735–769. [Google Scholar] [CrossRef] [PubMed]
- Schultze-Kraft, M. Evolution of Estimated Coca Cultivation and Cocaine Production in South America (Bolivia, Colombia and Peru) and of the Actors, Modalities and Routes of Cocaine Trafficking to Europe; Background Paper Commissioned by the EMCDDA for the 2016 EU Drug Marke; European Monitoring Centre for Drugs and Drug Addiction: Lisbon, Portugal, 2016; pp. 1–15.
- WHO. Annex 1 19th WHO Model List of Essential Medicines; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Langmead, C.J.; Watson, J.; Reavill, C. Muscarinic acetylcholine receptors as CNS drug targets. Pharmacol. Ther. 2008, 117, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Rothman, R.B.; Baumann, M.H.; Dersch, C.M.; Romero, D.V.; Rice, K.C.; Carroll, F.I.V.Y.; Partilla, J.S. Amphetamine-Type Central Nervous Norepinephrine More Potently Than They Release Dopamine and Serotonin. Synapse 2001, 39, 32–41. [Google Scholar] [CrossRef]
- Dräger, B. Identification and Quantification of Calystegines, Polyhydroxyl Nortropane Alkaloids. Phytochem. Anal. 1994, 6, 31–37. [Google Scholar] [CrossRef]
- Griffin, W.J.; Lin, G.D. Chemotaxonomy and geographical distribution of tropane alkaloids. Phytochemistry 2000, 53, 623–637. [Google Scholar] [CrossRef]
- Müller, J. Hexensalben und Liebestränke: Ein Beitrag zur Kulturgeschichte der Nachtschattengewächse Hexensalben und Liebestränke. Gesnerus Swiss J. Hist. Med. Sci. 1998, 55, 205–220. [Google Scholar]
- Ulbricht, C.; Basch, E.; Hammerness, P.; Vora, M.; Wylie, J.; Woods, J. An Evidence-Based Systematic Review of Belladonna by the Natural Standard Research Collaboration. J. Herb. Pharmacother. 2005, 4, 61–90. [Google Scholar] [CrossRef]
- Heinrich, M.; Jäger, A.K. Ethnopharmacology; Wiley: West Sussex, UK, 2015; ISBN 9781118930748. [Google Scholar]
- Endo, T.; Yamada, Y. Alkaloid production in cultured roots of three species of Duboisia. Phytochemistry 1985, 24, 1233–1236. [Google Scholar] [CrossRef]
- Naudé, T.W. Datura spp. and Other Related Plants; Elsevier Inc.: Atlanta, GA, USA, 2007; ISBN 9780123704672. [Google Scholar]
- Bobick, J.E.; Balaban, N.E. The Handy Science Answer Book; Science and Technology Department of the Carnegie Library of Pittsburgh: Pittsburgh, PA, USA, 2011; ISBN 1578593212. [Google Scholar]
- Wiart, C. Ethnopharmacology of Medicinal Plants; Humana Press: Totowa, NJ, USA, 2006; ISBN 1588297489. [Google Scholar]
- Pearn, J.; Thearle, J. The history of hyoscine. Hist. Sci. Med. 1982, 17, 257–261. [Google Scholar]
- Kim, N.; Estrada, O.; Chavez, B.; Stewart, C.; D’Auria, J. Tropane and Granatane Alkaloid Biosynthesis: A Systematic Analysis. Molecules 2016, 21, 1510. [Google Scholar] [CrossRef] [PubMed]
- Döbereiner, J.W. Deutsches Apothekerbuch; zum Gebrauche bei Vorlesungen und zum Selbstunterrichte für Apotheker, Droguisten, Aerzte und Medicin-Studirende; Walz: Stuttgart, Germany, 1847. [Google Scholar]
- Geiger, P.L.; Hesse, O. Über das Atropin. Pharm. Cent. 1833, 49, 768. [Google Scholar]
- Lossen, W. Ueber das Atropin. Ann. der Chemie und Pharm. 1864. [Google Scholar] [CrossRef]
- Wolffenstein, R. Die Pflanzenalkaloide; Julius Springer Verlag: Heidelberg, Germany, 1922. [Google Scholar]
- Ladenburg, A. Künstliches Atropin. Berichte der Dtsch. Chem. Gesellschaft 1879, 12, 941–944. [Google Scholar] [CrossRef]
- Gaedcke, F. Ueber das Erythroxylin. Arch. der Pharm. 1855, 132, 141–150. [Google Scholar] [CrossRef]
- Niemann, A. Ueber eine neue organische Base in den Cocablättern. Arch. Der Pharm. 1860, 153, 129–155. [Google Scholar] [CrossRef]
- Hardegger, E.; Ott, H. Konfiguration des Cocains und Derivate der Ecgoninsäure. Helv. Chim. Acta 1954, 331, 312–320. [Google Scholar] [CrossRef]
- Freud, S. Über Coca; Moritz Perles Verlag: Wien, Austria, 1885; ISBN 978-3226009682. [Google Scholar]
- Jocković, N.; Fischer, W.; Brandsch, M.; Brandt, W.; Dräger, B. Inhibition of human intestinal b-glucosidases by calystegines. J. Agric. Food Chem. 2013, 61, 5550–5557. [Google Scholar] [CrossRef] [PubMed]
- Wess, J.; Duttaroy, A.; Zhang, W.; Gomeza, J.; Cui, Y.; Miyakawa, T.; Bymaster, F.P.; Mckinzie, L.; Felder, C.C.; Lamping, K.G.; et al. M 1 -M 5 Muscarinic Receptor Knockout Mice as Novel Tools to Study the Physiological Roles of the Muscarinic Cholinergic System. Recept. Channels 2003, 9, 279–290. [Google Scholar] [CrossRef]
- Watts, S.W.; Kanagy, N.L.; Lombard, J.H. Receptor-Mediated Events in the Microcirculation. Microcirculation 2008, 285–348. [Google Scholar] [CrossRef]
- EFSA Scientific Opinion on Tropane alkaloids in food and feed. EFSA J. 2013, 9, 1–134. [CrossRef]
- Renner, U.D.; Oertel, R.; Kirch, W. Pharmacokinetics and pharmacodynamics in clinical use of scopolamine. Ther. Drug Monit. 2005, 27, 655–665. [Google Scholar] [CrossRef]
- Kanto, J.; Kentala, E.; Kaila, T.; Pihlajamäki, K. Pharmacokinetics of scopolamine during caesarean section: Relationship between serum concentration and effect. Acta Anaesthesiol. Scand. 1989, 33, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Boffa, J.-M.; Yarnéogo, G.; Nikiéma, P.; Knudson, D.M. Domestication and Commercialization of Non-Timber Forest Products in Agroforestry Systems; Food and Agriculture Organization of the United Nations: Rome, Italy, 1996; ISBN 9251039356. [Google Scholar]
- Robenshtok, E.; Luria, S.; Tashma, Z.; Hourvitz, A. Adverse reaction to atropine and the treatment of organophosphate intoxication. Isr. Med. Assoc. J. 2002, 4, 535–539. [Google Scholar] [PubMed]
- Gyermek, L. Structure-activity relationships among derivatives of dicarboxylic acid esters of tropine. Pharmacol. Ther. 2002, 96, 1–21. [Google Scholar] [CrossRef]
- Sneader, W. Drug Discovery: A History; Wiley: West Sussex, UK, 2005; ISBN 0471899798. [Google Scholar]
- Rudy, D.; Cline, K.; Harris, R.; Goldberg, K.; Dmochowski, R. Multicenter phase III trial studying trospium chloride in patients with overactive bladder. Urology 2006, 67, 275–280. [Google Scholar] [CrossRef]
- Sorbe, B.; Andersson, H.; Schmidt, M.; Söderberg, M.; Högberg, T.; Wernstedt, L.; Janson, E.T.; Ehrnström, B.; Kjaer, M.; Havsteen, H.; et al. Tropisetron (Navoban) in the prevention of chemotherapy-induced nausea and vomiting—the Nordic experience. Support. Care Cancer 1994, 2, 393–399. [Google Scholar] [CrossRef]
- Mutschler, E.; Geisslinger, G.; Kroemer, H.; Ruth, P.; Schäfer-Korting, M. Mutschler Arzneimittelwirkungen, 9th ed.; WVG: Stuttgart, Germany, 2008; ISBN 978-3-8047-1952-1. [Google Scholar]
- Barnes, P.J. The Pharmacological Properties of Tiotropium. Chest 2000, 117, 63S–66S. [Google Scholar] [CrossRef]
- Schlagmann, C.; Remien, J. Klinische Schrift Zur Behandlung der Parkinson-Krankheit. Klin. Wochenschr. 1986, 64, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Lv, D.; Shen, M.; Zhang, Y.; Zhou, D.; Chen, Z.; Wang, C. BDNF mediates the protective effects of scopolamine in reserpine-induced depression-like behaviors via up-regulation of 5-HTT and TPH1. Psychiatry Res. 2019, 271, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Park, L.; Furey, M.; Nugent, A.; Farmer, C.; Ellis, J.; Szczepanik, J.; Lener, M.; Zarate, C. F173. Negative Trial of Scopolamine in Major Depressive Disorder Does Not Demonstrate Neurophysiological Changes Seen With the Antidepressant Response of Ketamine. Biol. Psychiatry 2018, 83, S305–S306. [Google Scholar] [CrossRef]
- Williams, N. Cocaine and Metabolites: Relationship between pharmacological activity and inhibitory action on dopamine uptake into struatal synaptosomes. Prog. Neuropsychopharmacol. 1977, 1, 265–269. [Google Scholar] [CrossRef]
- Berger, H. Archiv Für Psychiatrie Und Nervenkrankheiten. Clin. Neurophysiol. 1931, 28, 95–132. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products. Pharm. Sci. 2002, 0471496405, 514. [Google Scholar]
- Sinatra, R.S.; Jahr, J.S.; Watkins-Pitchford, M. The Essence of Analgesia and Analgesics; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780511841378. [Google Scholar]
- Agostinelli, E. Polyamines and transglutaminases: Biological, clinical and biotechnological perspectives. Amino Acids 2014, 46, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Robins, R.J.; Parr, A.J.; Walton, N.J. Studies on the biosynthesis of tropane alkaloids in Datura stramonium L. transformed root cultures. Planta 1991, 183, 196–201. [Google Scholar] [CrossRef]
- Biastoff, S.; Brandt, W.; Dräger, B. Putrescine N-methyltransferase—The start for alkaloids. Phytochemistry 2009, 70, 1708–1718. [Google Scholar] [CrossRef]
- Mizusaki, S.; Tanabe, Y.; Noguchi, M.; Tamaki, E. N-methylputrescine oxidase from tobacco roots. Phytochemistry 1972, 11, 2757–2762. [Google Scholar] [CrossRef]
- Courdavault, V. Biosynthesis and Regulation of Alkaloids. In Plant Developmental Biology; Humana Press: Totowa, NJ, USA, 2010; Volume 655, pp. 139–160. ISBN 978-1-60761-764-8. [Google Scholar]
- Bedewitz, M.A.; Jones, A.D.; D’Auria, J.C.; Barry, C.S. Tropinone synthesis via an atypical polyketide synthase and P450-mediated cyclization. Nat. Commun. 2018, 9, 5281. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Nakajima, K.; Ongena, G.; Yasuyuki, Y. Two Tropinone Reductases with Distinct Stereospecificities from Cultured Roots of Hyoscyamus niger. Plant Physiol. 1992, 100, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Bedewitz, M.A.; Gongora-Castillo, E.; Uebler, J.B.; Gonzales-Vigil, E.; Wiegert-Rininger, K.E.; Childs, K.L.; Hamilton, J.P.; Vaillancourt, B.; Yeo, Y.-S.; Chappell, J.; et al. A Root-Expressed l-Phenylalanine:4-Hydroxyphenylpyruvate Aminotransferase Is Required for Tropane Alkaloid Biosynthesis in Atropa belladonna. Plant Cell 2014, 26, 3745–3762. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Yang, C.; Yuan, L.; Xiang, D.; Lan, X.; Chen, M.; Liao, Z. A Phenylpyruvic Acid Reductase Is Required for Biosynthesis of Tropane Alkaloids. Org. Lett. 2018, 20, 7807–7810. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, G.W.; Jirschitzka, J.; Porta, T.; Reichelt, M.; Luck, K.; Torre, J.C.P.; Dolke, F.; Varesio, E.; Hopfgartner, G.; Gershenzon, J.; et al. The Last Step in Cocaine Biosynthesis Is Catalyzed by a BAHD Acyltransferase. Plant Physiol. 2015, 167, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Robins, R.J.; Bachmann, P.; Robinson, T.; Rhodes, M.J.; Yamada, Y. The formation of 3 alpha- and 3 beta-acetoxytropanes by Datura stramonium transformed root cultures involves two acetyl-CoA-dependent acyltransferases. FEBS Lett. 1991, 292, 293–297. [Google Scholar] [PubMed]
- Li, R.; Reed, D.W.; Liu, E.; Nowak, J.; Pelcher, L.E.; Page, J.E.; Covello, P.S. Functional genomic analysis of alkaloid biosynthesis in Hyoscyamus niger reveals a cytochrome P450 involved in littorine rearrangement. Chem. Biol. 2006, 13, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Yamada, Y. Hyoscyamine 6b-Hydroxylase, a 2-Oxoglutarate-Dependent Dioxygenase, in Alkaloid Producing Root Cultures. Plant Physiol. 1985, 81, 619–625. [Google Scholar] [CrossRef]
- Hashimoto, T.; Matsuda, J.; Yamada, Y. Two-step epoxidation of hyoscyamine to scopolamine is catalyzed by bifunctional hyoscyamine-6β-hydroxylase. FEBS Lett. 1993, 329, 35–39. [Google Scholar] [CrossRef]
- Korolev, S.; Ikeguchi, Y.; Skarina, T.; Beasley, S.; Arrowsmith, C.; Edwards, A.; Joachimiak, A.; Pegg, A.E.; Savchenko, A. The crystal structure of spermidine synthase with a multisubstrate adduct inhibitor. Nat. Struct. Biol. 2002, 9, 27–31. [Google Scholar] [CrossRef]
- Biastoff, S.; Reinhardt, N.; Reva, V.; Brandt, W.; Draeger, B. Evolution of putrescine N-methyltransferase from spermidine synthase demanded alterations in substrate binding. FEBS Lett. 2009, 583, 3367–3374. [Google Scholar] [CrossRef] [PubMed]
- Teuber, M.; Azemi, M.E.; Namjoyan, F.; Meier, A.C.; Wodak, A.; Brandt, W.; Draeger, B. Putrescine N-methyltransferases—A structure-function analysis. Plant Mol. Biol. 2007, 63, 787–801. [Google Scholar] [CrossRef]
- Oppermann, U.; Filling, C.; Hult, M.; Shafqat, N.; Wu, X.; Lindh, M.; Shafqat, J.; Nordling, E.; Kallberg, Y.; Persson, B.; et al. Short-chain dehydrogenases/reductases (SDR): The 2002 update. Chem. Biol. Interact. 2003, 143–144, 247–253. [Google Scholar] [CrossRef]
- Kushwaha, A.K.; Sangwan, N.S.; Tripathi, S.; Sangwan, R.S. Molecular cloning and catalytic characterization of a recombinant tropine biosynthetic tropinone reductase from Withania coagulans leaf. Gene 2013, 516, 238–247. [Google Scholar] [CrossRef]
- Qiang, W.; Xia, K.; Zhang, Q.; Zeng, J.; Huang, Y.; Yang, C.; Chen, M.; Liu, X.; Lan, X.; Liao, Z. Functional characterisation of a tropine-forming reductase gene from Brugmansia arborea, a woody plant species producing tropane alkaloids. Phytochemistry 2016, 127, 12–22. [Google Scholar] [CrossRef]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Chesters, N.C.J.E.; O’Hagan, D.; Robins, R.J. The biosynthesis of tropic acid: The (R)-d-phenyllactyl moiety is processed by the mutase involved in hyoscyamine biosynthesis in Datura stramonium. J. Chem. Soc. Chem. Commun. 1995, 0, 127–128. [Google Scholar] [CrossRef]
- Ollagnier, S.; Kervio, E.; Rétey, J. The role and source of 5′-deoxyadenosyl radical in a carbon skeleton rearrangement catalyzed by a plant enzyme. FEBS Lett. 1998, 437, 309–312. [Google Scholar] [CrossRef]
- Pramod, K.K.; Singh, S.; Jayabaskaran, C. Biochemical and structural characterization of recombinant hyoscyamine 6β-hydroxylase from Datura metel. Plant Physiol. Biochem. 2010, 48, 966–970. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yamada, Y. Purification and characterization of hyoscyamine 6 beta-hydroxylase from root cultures of Hyoscyamus niger L. Hydroxylase and epoxidase activities in the enzyme preparation. Eur. J. Biochem. 1987, 164, 277–285. [Google Scholar] [CrossRef]
- Suzuki, K.; Yamada, Y.; Hashimoto, T. Expression of Atropa belladonna putrescine N-methyltransferase gene in root pericycle. Plant Cell Physiol. 1999, 40, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Kanegae, T.; Kajiya, H.; Amano, Y.; Hashimoto, T.; Yamada, Y. Species-dependent expression of the hyoscyamine 6 beta-hydroxylase gene in the pericycle. Plant Physiol. 1994, 105, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Hashimoto, T. Two tropinone reductases, that catalyze opposite stereospecific reductions in tropane alkaloid biosynthesis, are localized in plant root with different cell-specific patterns. Plant Cell Physiol. 1999, 40, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, H.; Richter, U.; Keiner, R.; Brabant, A.; Hause, B.; Draeger, B. Immunolocalisation of two tropinone reductases in potato (Solanum tuberosum L.) root, stolon and tuber sprouts. Planta 2006, 225, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Yamada, Y.; Hashimoto, T. Jasmonate induction of putrescine N-methyltransferase genes in the root of Nicotiana sylvestris. Plant Cell Physiol. 2000, 41, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Lauersen, K.J. Eukaryotic microalgae as hosts for light-driven heterologous isoprenoid production. Planta 2018, 249, 155–180. [Google Scholar] [CrossRef] [PubMed]
- Leete, E.; Endo, T.; Yamada, Y. Biosynthesis of nicotine and scopolamine in a root culture of Duboisia leichhardtii. Phytochemistry 1990, 29, 1847–1851. [Google Scholar] [CrossRef]
- Hemscheidt, T.; Spenser, I.D. Biosynthesis of 6β.-hydroxytropine in Datura stramonium: Nonregiospecific incorporation of [1,2-13C2]acetate. J. Am. Chem. Soc. 1992, 114, 5472–5473. [Google Scholar] [CrossRef]
- Hoye, T.R.; Bjorklund, J.A.; Koltun, D.O.; Renner, M.K. N-methylputrescine oxidation during cocaine biosynthesis: Study of prochiral methylene hydrogen discrimination using the remote isotope method. Org. Lett. 2000, 2, 3–5. [Google Scholar] [CrossRef]
- Leete, E.; Bjorklund, J.A.; Sung, H.K. The biosynthesis of the benzoyl moiety of cocaine. Phytochemistry 1988, 27, 2553–2556. [Google Scholar] [CrossRef]
- Scholl, Y.; Höke, D.; Dräger, B. Calystegines in Calystegia sepium derive from the tropane alkaloid pathway. Phytochemistry 2001, 58, 883–889. [Google Scholar] [CrossRef]
- Goldmann, A.; Message, B.; Tepfer, D.; Molyneux, R.J.; Duclos, O. Biological Activities of the Nortropane Alkaloid, Calystegine B 2 and Analogs: Structure—Function Relationships. J. Nat. Prod. 1996, 59, 1137–1142. [Google Scholar] [CrossRef]
- Luanratana, O.; Griffin, W.J. Cultivation of a Duboisia Hybrid—Part A. J. Nat. Prod. 1980, 43, 477–491. [Google Scholar] [CrossRef]
- Ullrich, S.F.; Hagels, H.; Kayser, O. Scopolamine: a journey from the field to clinics. Phytochem. Rev. 2017, 16, 333–353. [Google Scholar] [CrossRef]
- Ullrich, S.F.; Rothauer, A.; Hagels, H.; Kayser, O. Influence of Light, Temperature and Macronutrients on Growth and Scopolamine Biosynthesis in Duboisia species. Planta Med. 2017, 83, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Oksman-Caldentey, K.-M.; Strauss, A. Somaclonal Variation of Scopolamine Content in Protoplast-Derived Cell Culture Clones of Hyoscyamus muticus. Planta Med. 1986, 52, 6–12. [Google Scholar] [CrossRef]
- Wang, K. Agrobacterium Protocols; Humana Press: Totowa, NJ, USA, 2007; ISBN 1588298434. [Google Scholar]
- Georgiev, M.I.; Agostini, E.; Ludwig-Müller, J.; Xu, J. Genetically transformed roots: From plant disease to biotechnological resource. Trends Biotechnol. 2012, 30, 528–537. [Google Scholar] [CrossRef]
- Maldonado-Mendoza, I.E.; Ayora-Talavera, T.; Loyola-Vargas, V.M. Establishment of hairy root cultures of Datura stramonium. Characterization and stability of tropane alkaloid production during long periods of subculturing. Plant Cell. Tissue Organ Cult. 1993, 33, 321–329. [Google Scholar] [CrossRef]
- Kamada, H.; Okamura, N.; Satake, M.; Harada, H.; Shimomura, K. Alkaloid production by hairy root cultures in Atropa belladonna. Plant Cell Rep. 1986, 5, 239–242. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yun, D.J.; Yamada, Y. Production of tropane alkaloids in genetically engineered root cultures. Phytochemistry 1993, 32, 713–718. [Google Scholar] [CrossRef]
- Falk, L.R.; Doran, P.M. Influence of inoculum morphology on growth of Atropa belladonna hairy roots and production of tropane alkaloids. Biotechnol. Lett. 1996, 18, 1099–1104. [Google Scholar] [CrossRef]
- Vakili, B.; Karimi, F.; Sharifi, M.; Behmanesh, M. Chromium-induced tropane alkaloid production and H6H gene expression in Atropa belladonna L. (Solanaceae) invitro-propagated plantlets. Plant Physiol. Biochem. 2012, 52, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Jaremicz, Z.; Luczkiewicz, M.; Kokotkiewicz, A.; Krolicka, A.; Sowinski, P. Production of tropane alkaloids in Hyoscyamus niger (black henbane) hairy roots grown in bubble-column and spray bioreactors. Biotechnol. Lett. 2014, 36, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ding, R.; Chai, Y.; Bonfill, M.; Moyano, E.; Oksman-Caldentey, K.-M.; Xu, T.; Pi, Y.; Wang, Z.; Zhang, H.; et al. Engineering tropane biosynthetic pathway in Hyoscyamus niger hairy root cultures. Proc. Natl. Acad. Sci. USA 2004, 101, 6786–6791. [Google Scholar] [CrossRef] [PubMed]
- Kai, G.; Zhang, A.; Guo, Y.; Li, L.; Cui, L.; Luo, X.; Lin, C.; Xiao, J. Enhancing the production of tropane alkaloids in transgenic Anisodus acutangulus hairy root cultures by over-expressing tropinone reductase I and hyoscyamine-6β-hydroxylase. Mol. Biosyst. 2012, 8, 2883–2890. [Google Scholar] [CrossRef] [PubMed]
- Cardillo, A.B.; Otálvaro, A.Á.M.; Busto, V.D.; Talou, J.R.; Velásquez, L.M.E.; Giulietti, A.M. Scopolamine, anisodamine and hyoscyamine production by Brugmansia candida hairy root cultures in bioreactors. Process Biochem. 2010, 45, 1577–1581. [Google Scholar] [CrossRef]
- Jouhikainen, K.; Lindgren, L.; Jokelainen, T.; Hiltunen, R.; Teeri, T.H.; Oksman-Caldentey, K.-M. Enhancement of scopolamine production in Hyoscyamus muticus L. hairy root cultures by genetic engineering. Planta 1999, 208, 545–551. [Google Scholar] [CrossRef]
- Moyano, E.; Fornalé, S.; Palazón, J.; Cusidó, R.M.; Bagni, N.; Piñol, M.T. Alkaloid production in Duboisia hybrid hairy root cultures overexpressing the pmt gene. Phytochemistry 2002, 59, 697–702. [Google Scholar] [CrossRef]
- Palazón, J.; Moyano, E.; Cusidó, R.M.; Bonfill, M.; Oksman-Caldentey, K.M.; Piñol, M.T. Alkaloid production in Duboisia hybrid hairy roots and plants overexpressing the h6h gene. Plant Sci. 2003, 165, 1289–1295. [Google Scholar] [CrossRef]
- Bourgaud, F.; Benoit, M.; De Picardie, U.; Verne, J. Développement D’ un Nouveau Procédé de Production D’actifs Pharmaceutiques à Partir de Plantes Médicinales: La Technologie des Plantes à Traire. Conference Paper: Les Rencontres du Végétal. Angers, France, 2013. Available online: http://agris.fao.org/agris-search/search.do?recordID=FR2016219199 (accessed on 28 December 2018).
- Xia, K.; Liu, X.; Zhang, Q.; Qiang, W.; Guo, J.; Lan, X.; Chen, M.; Liao, Z. Promoting scopolamine biosynthesis in transgenic Atropa belladonna plants with pmt and h6h overexpression under field conditions. Plant Physiol. Biochem. 2016, 106, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, M.; Yang, C.; Liu, X.; Zhang, L.; Lan, X.; Tang, K.; Liao, Z. Enhancing the scopolamine production in transgenic plants of Atropa belladonna by overexpressing pmt and h6h genes. Physiol. Plant. 2011, 143, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Cardillo, A.B.; Talou, J.R.; Giulietti, A.M. Expression of Brugmansia candida Hyoscyamine 6beta-Hydroxylase gene in Saccharomyces cerevisiae and its potential use as biocatalyst. Microb. Cell Fact. 2008, 7, 17. [Google Scholar] [CrossRef] [PubMed]
- Kai, G.; Liu, Y.; Wang, X.; Yang, S.; Fu, X.; Luo, X.; Liao, P. Functional identification of hyoscyamine 6β-hydroxylase from Anisodus acutangulus and overproduction of scopolamine in genetically-engineered Escherichia coli. Biotechnol. Lett. 2011, 33, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Van Belkum, M.J.; Vederas, J.C. Functional characterization of recombinant hyoscyamine 6β-hydroxylase from Atropa belladonna. Bioorganic Med. Chem. 2012, 20, 4356–4363. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhu, P.; Meng, C.; Cheng, K.D.; He, H.X. Molecular cloning, expression and characterization of hyoscyamine 6beta-hydroxylase from hairy roots of Anisodus tanguticus. Planta 2005, 71, 249–253. [Google Scholar] [CrossRef]
- Fischer, C.; Kwon, M.; Ro, D.; Van Belkum, M.J.; Vederas, J.C. Isolation, expression and biochemical characterization of recombinant hyoscyamine-6β- hydroxylase from Brugmansia sanguinea—Tuning the scopolamine production. Med. Chem. Commun. 2018, 9, 888–892. [Google Scholar] [CrossRef]
- Cao, Y.; He, Y.; Li, H.; Kai, G.; Xu, J.; Yu, H. Efficient biosynthesis of rare natural product scopolamine using E. coli cells expressing a S14P/K97A mutant of hyoscyamine 6b -hydroxylase. J. Biotechnol. 2015, 211, 123–129. [Google Scholar] [CrossRef]
- Dehghan, E.; Reed, D.W.; Covello, P.S.; Hasanpour, Z.; Palazon, J.; Oksman-Caldentey, K.M.; Ahmadi, F.S. Genetically engineered hairy root cultures of Hyoscyamus senecionis and H. muticus: Ploidy as a promising parameter in the metabolic engineering of tropane alkaloids. Plant Cell Rep. 2017, 36, 1615–1626. [Google Scholar] [CrossRef]
- Dehghan, E.; Shahriari Ahmadi, F.; Ghotbi Ravandi, E.; Reed, D.W.; Covello, P.S.; Bahrami, A.R. An atypical pattern of accumulation of scopolamine and other tropane alkaloids and expression of alkaloid pathway genes in Hyoscyamus senecionis. Plant Physiol. Biochem. 2013, 70, 188–194. [Google Scholar] [CrossRef]
- Naik, T.; Vanitha, S.C.; Rajvanshi, P.K.; Chandrika, M.; Kamalraj, S.; Jayabaskaran, C. Novel Microbial Sources of Tropane Alkaloids: First Report of Production by Endophytic Fungi Isolated from Datura metel L. Curr. Microbiol. 2017, 75, 1–7. [Google Scholar] [CrossRef]
- De Luca, V.; Salim, V.; Thamm, A.; Masada, S.A.; Yu, F. Making iridoids/secoiridoids and monoterpenoid indole alkaloids: Progress on pathway elucidation. Curr. Opin. Plant Biol. 2014, 19, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Polturak, G.; Heinig, U.; Grossman, N.; Battat, M.; Leshkowitz, D.; Malitsky, S.; Rogachev, I.; Aharoni, A. Transcriptome and Metabolic Profiling Provides Insights into Betalain Biosynthesis and Evolution in Mirabilis jalapa. Mol. Plant 2018, 11, 189–204. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Overexpression of | Amount | Citation | |
|---|---|---|---|---|
| Hyoscyamine | Scopolamine | |||
| Atropa belladonna | - | 0.371 ± 0.013% DW | 0.024 ± 0.010% DW | Kamada et al., 1986 [94] |
| H. niger h6h | 0.02% *) | 0.45% *) | Hashimoto et al., 1993b [95] | |
| - | 2.1 + 0.2 mg g−1 DW | n.d. | Falk and Doran, 1996 [96] | |
| **) | 0.31 mg g−1 DW | 0.27 mg g−1 DW | Vakili et al., 2012 [97] | |
| Hyoscyamus niger | - | 1.6 mg g−1 DW | 5.3 mg g−1 DW | Jaremicz et al., 2014 [98] |
| pmt, h6h | n.d. | 411 mg L−1 | Zhang et al., 2004 [99] | |
| Anisodus acutangulus | h6h | 0.789 ± 0.078 mg g−1 DW | 0.070 ± 0.003 mg g−1 DW | Kai et al., 2012 [100] |
| tr-I | 2.479 ± 0.432 mg g−1 DW | 0.023 ± 0.004 mg g−1 DW | ||
| tr-I, h6h | 2.286 ± 0.46 mg g−1 DW | 0.072 ± 0.018 mg g−1 DW | ||
| Brugmansia candida **) | - | 0.35 ± 0.07 mg g−1 DW | 1.05 mg g−1 DW | Cardillo et al., 2010 [101] |
| Hyoscyamus muticus | h6h | 287.7 mg L−1 | 14.41 mg L−1 | Jouhikainen et al., 1999 [102] |
| Duboisia myoporoides | pmt | no increase observed | Moyano et al., 2002 [103] | |
| h6h | n.d. | 24.93 mg g−1 DW | Palazón et al., 2003 [104] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohnen-Johannsen, K.L.; Kayser, O. Tropane Alkaloids: Chemistry, Pharmacology, Biosynthesis and Production. Molecules 2019, 24, 796. https://doi.org/10.3390/molecules24040796
Kohnen-Johannsen KL, Kayser O. Tropane Alkaloids: Chemistry, Pharmacology, Biosynthesis and Production. Molecules. 2019; 24(4):796. https://doi.org/10.3390/molecules24040796
Chicago/Turabian StyleKohnen-Johannsen, Kathrin Laura, and Oliver Kayser. 2019. "Tropane Alkaloids: Chemistry, Pharmacology, Biosynthesis and Production" Molecules 24, no. 4: 796. https://doi.org/10.3390/molecules24040796
APA StyleKohnen-Johannsen, K. L., & Kayser, O. (2019). Tropane Alkaloids: Chemistry, Pharmacology, Biosynthesis and Production. Molecules, 24(4), 796. https://doi.org/10.3390/molecules24040796