GC-MS-Based Metabolomics to Reveal the Protective Effect of Gross Saponins of Tribulus terrestris Fruit against Ischemic Stroke in Rat

 ,
,
Abstract
:1. Introduction
2. Results
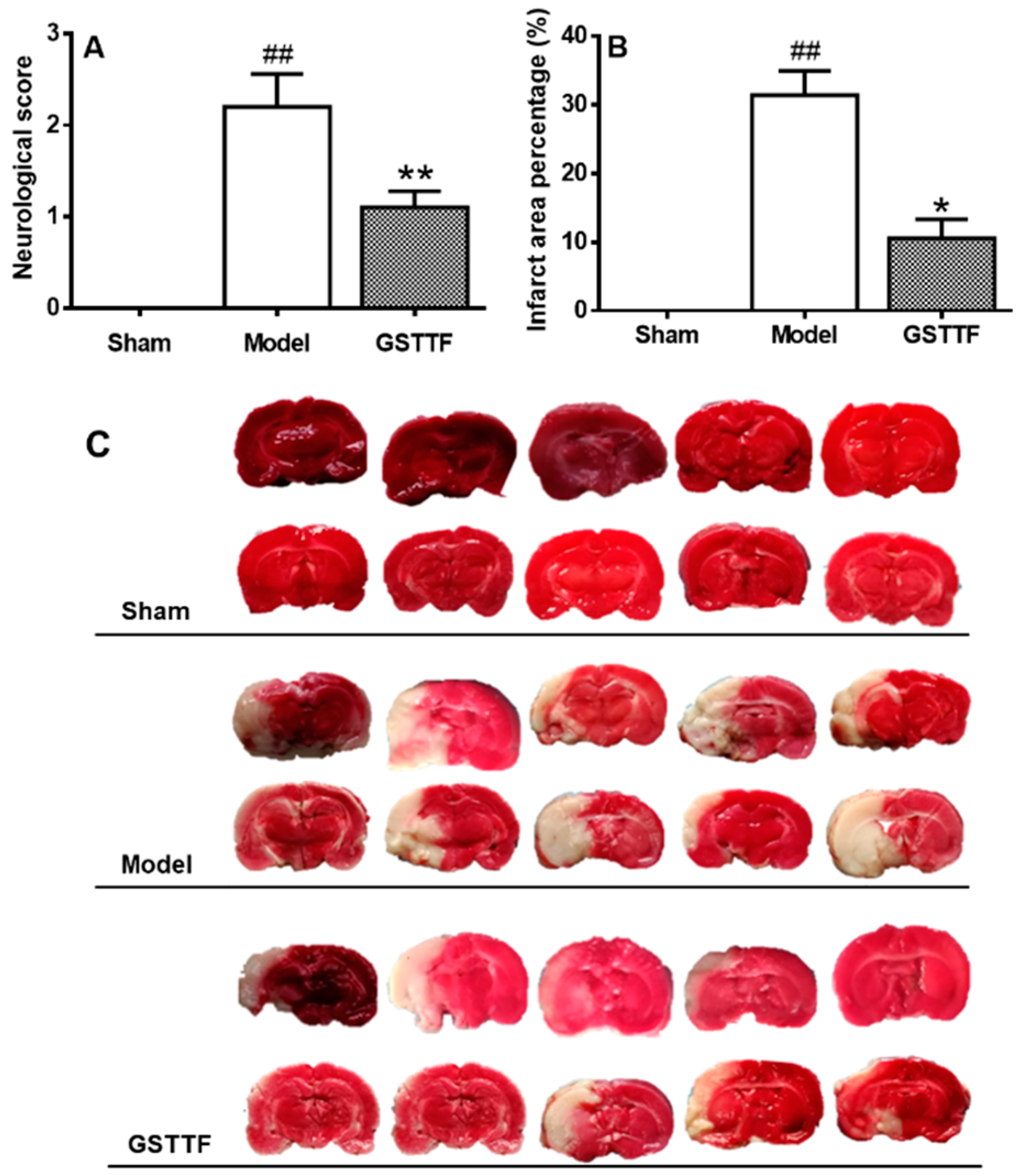
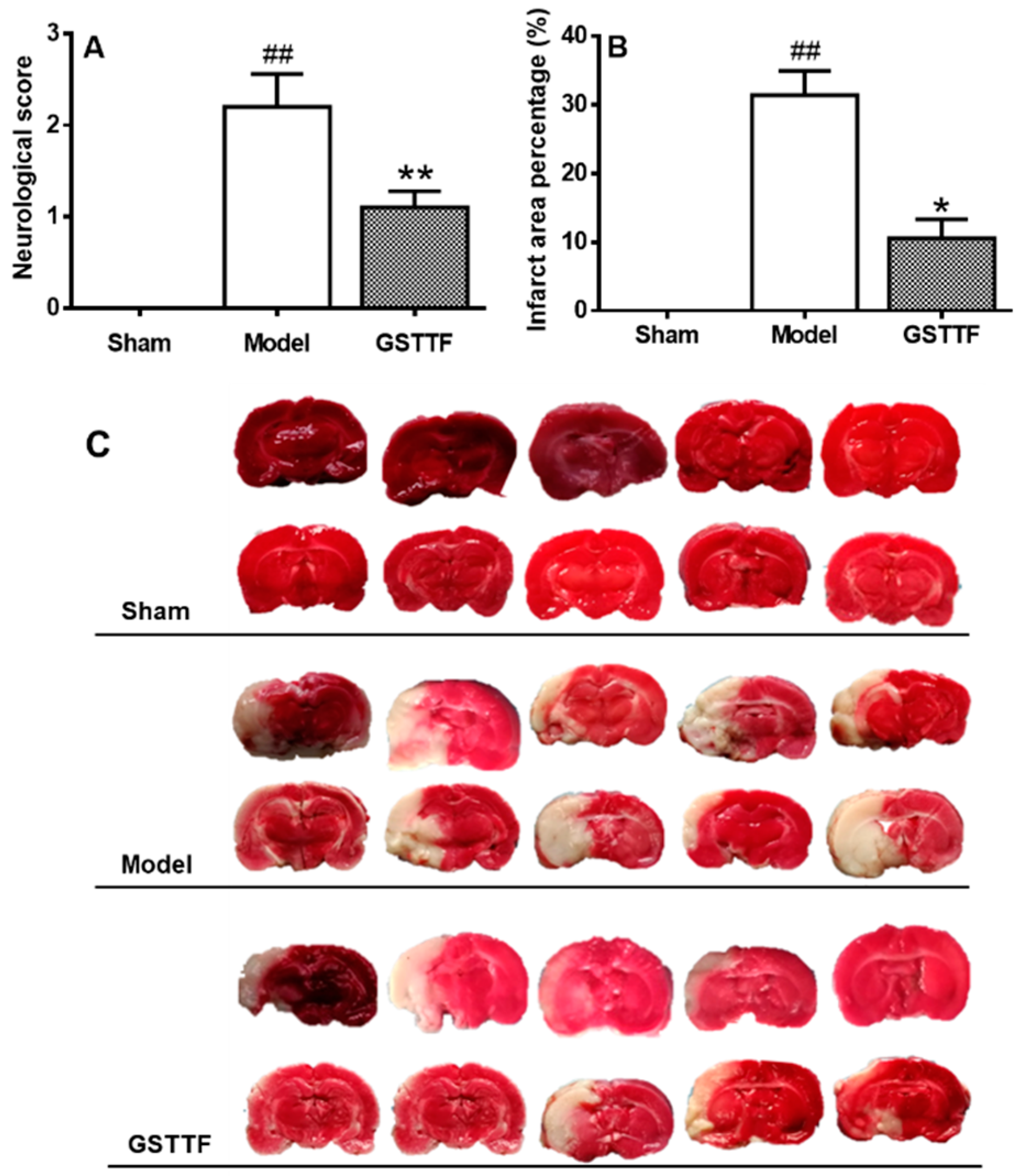
2.1. The Effects of GSTTF on MACO in Rats
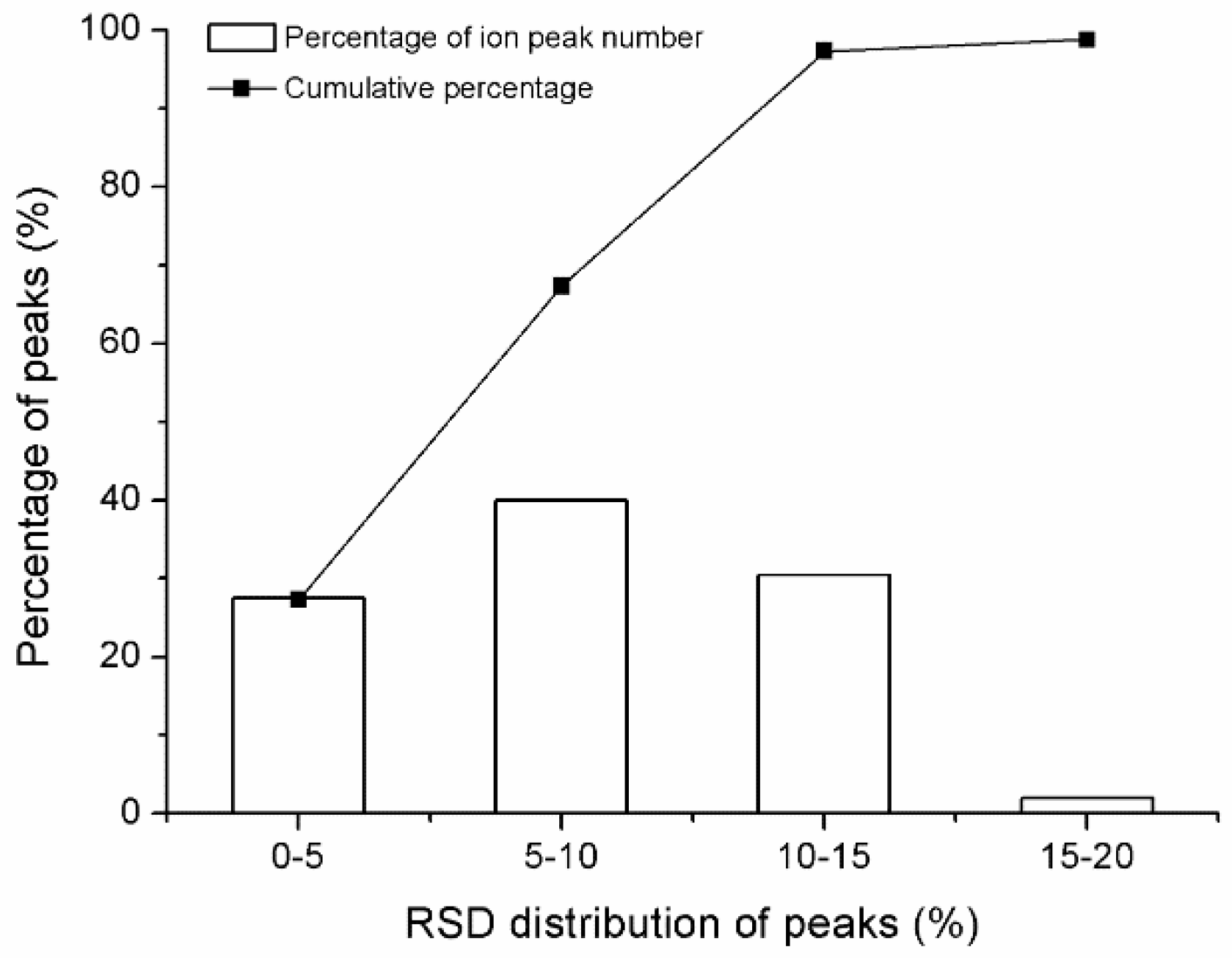
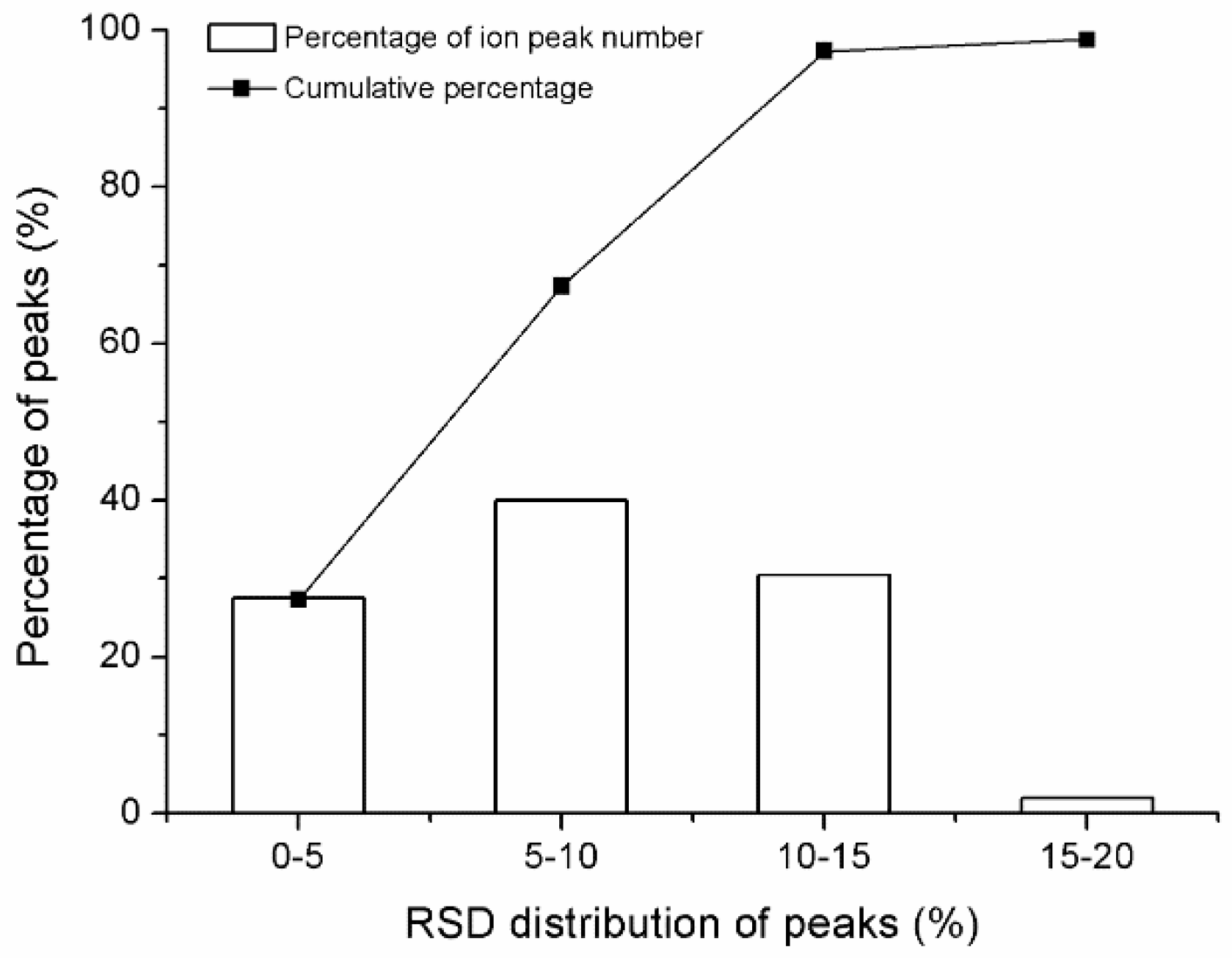
2.2. GC-TOF/MS Method Validation
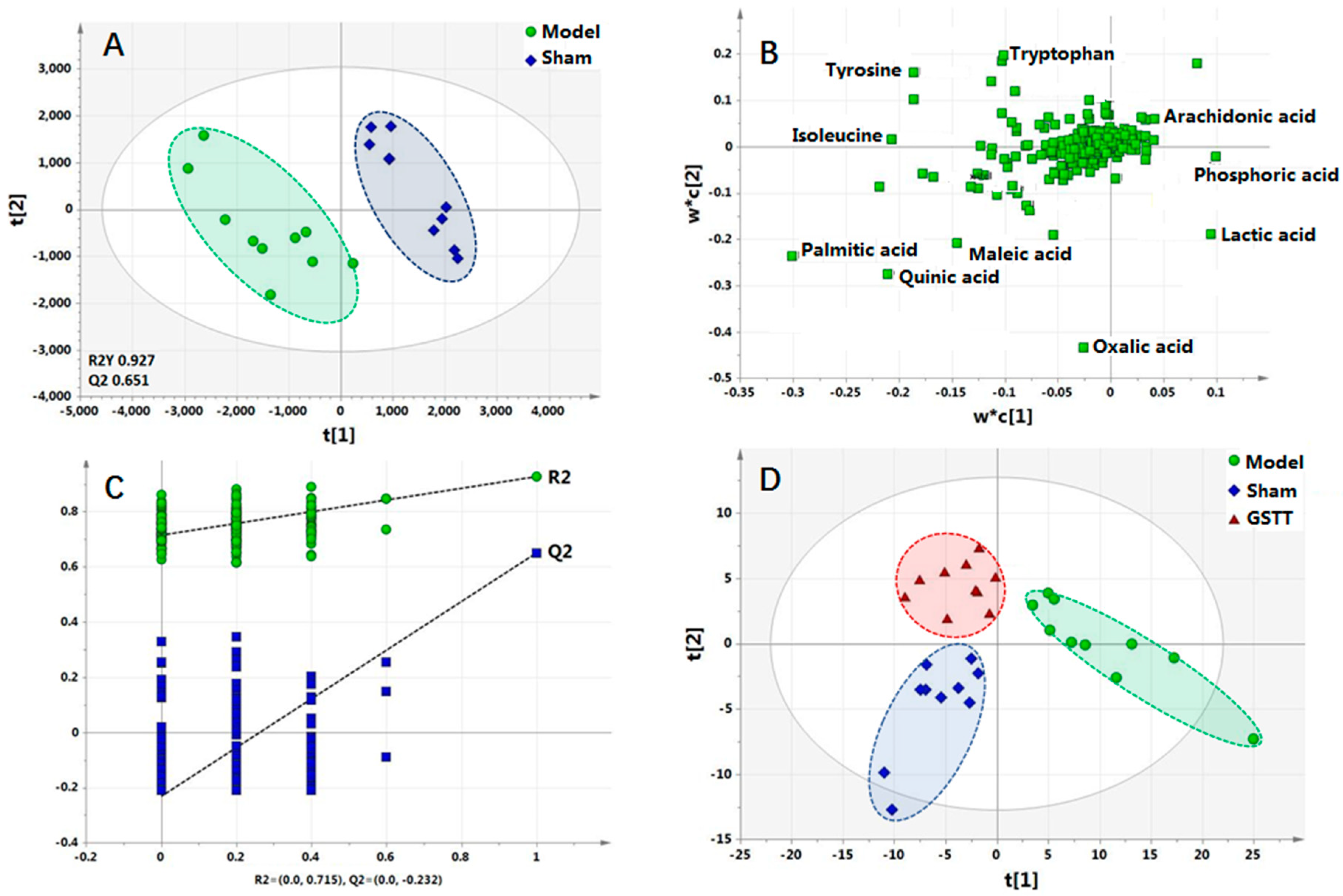
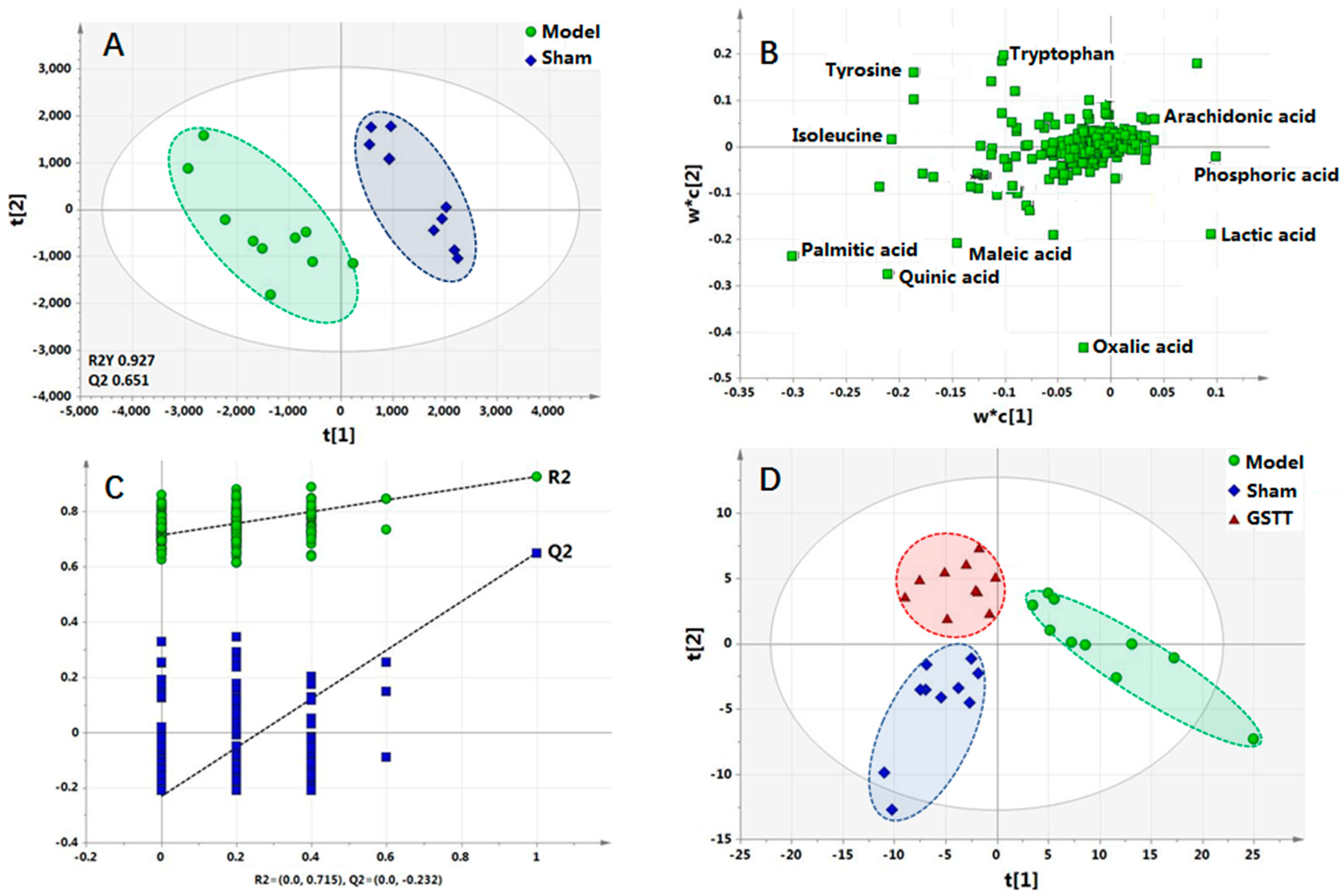
2.3. Metabolic Profile in Serum
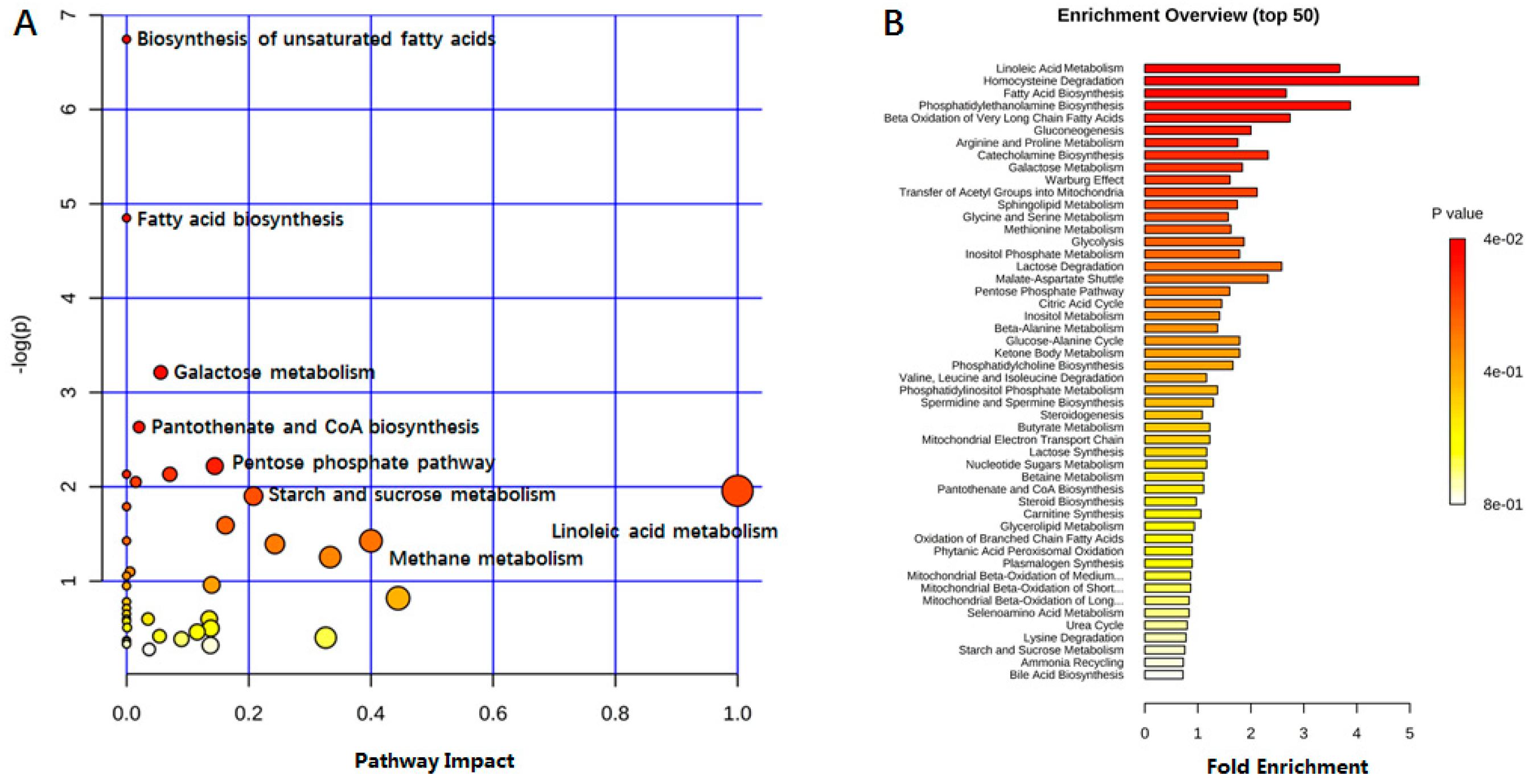
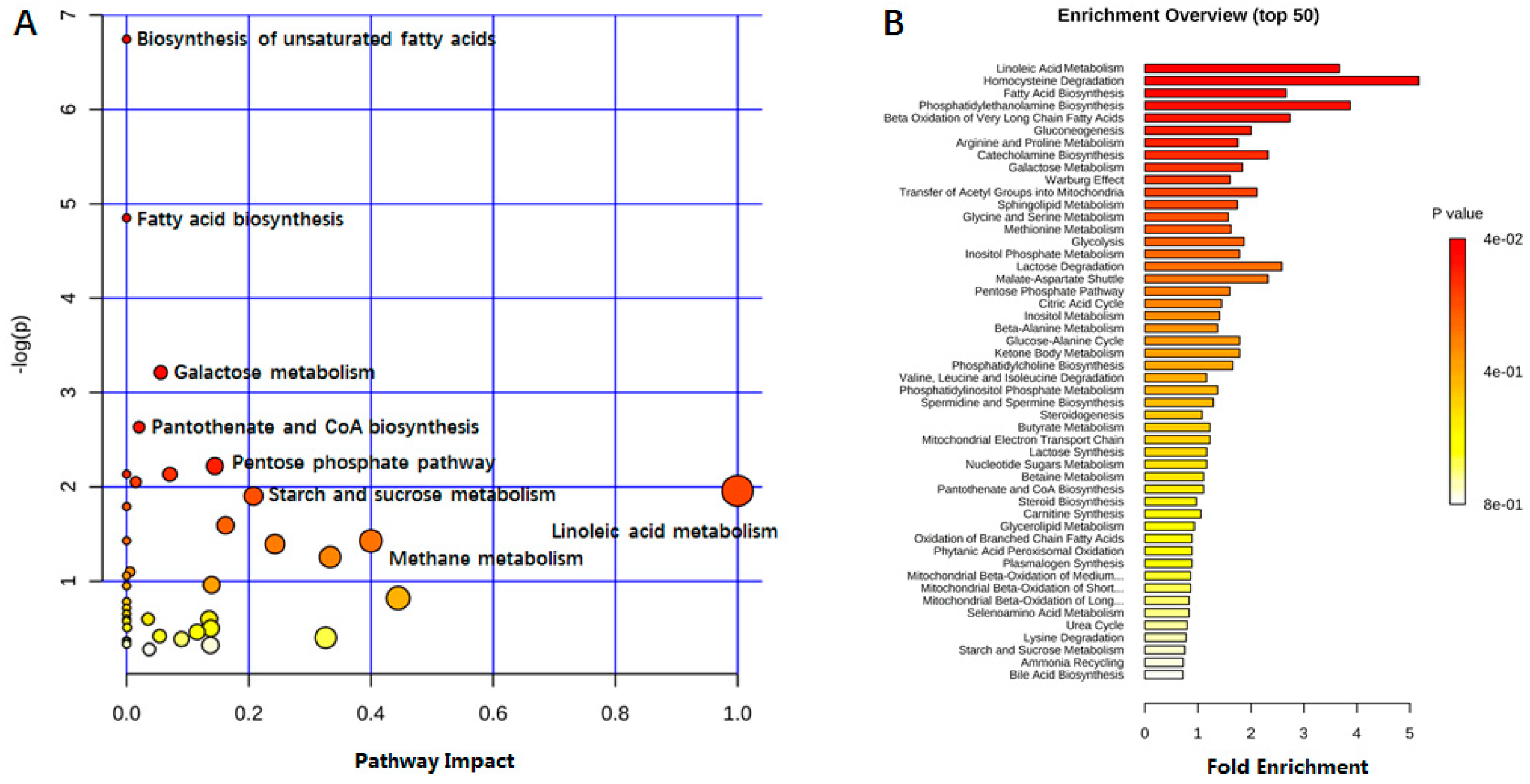
2.4. Biomarker Selection and Metabolic Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Treatments
4.3. Evaluation of Neurological Defects
4.4. Infarct Volume Measurement
4.5. Sample Preparation
4.6. GC-MS Analysis
4.7. Data Processing and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Strong, K.; Mathers, C.; Bonita, R. Preventing stroke: Saving lives around the world. Lancet Neurol. 2007, 6, 182–187. [Google Scholar] [CrossRef]
- Kunz, A.; Iadecola, C. Cerebral vascular dysregulation in the ischemic brain. Handbook Clin. Neurol. 2008, 92, 283–305. [Google Scholar]
- Berkhemer, O.A.; Fransen, P.S.; Beumer, D.; Van Den Berg, L.A.; Lingsma, H.F.; Yoo, A.J.; Schonewille, W.J.; Vos, J.A.; Nederkoorn, P.J.; Wermer, M.J. A randomized trial of intraarterial treatment for acute ischemic stroke. New Engl. J. Med. 2015, 372, 11–20. [Google Scholar] [CrossRef]
- Fonarow, G.C.; Smith, E.E.; Saver, J.L.; Reeves, M.J.; Bhatt, D.L.; Grau-Sepulveda, M.V.; Olson, D.M.; Hernandez, A.F.; Peterson, E.D.; Schwamm, L.H. Timeliness of tissue-type plasminogen activator therapy in acute ischemic stroke: Patient characteristics, hospital factors, and outcomes associated with door-to-needle times within 60 min. Circulation 2011, 125, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Venketasubramanian, N.; Gan, R.N.; Lambert, C.; Picard, D.; Chan, B.P.; Chan, E.; Bousser, M.G.; Xuemin, S. Danqi Piantang Jiaonang (DJ), a traditional Chinese medicine, in poststroke recovery. Stroke 2009, 40, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.-L.; Aa, J.-Y.; Feng, S.-Q.; Wang, S.-Y.; Wang, P.; Zhang, Y.; Ouyang, B.-C.; Wang, J.-K.; Zhu, Y.-J.; Huang, W.-Z. Exploring the neuroprotective effects of ginkgolides injection in a rodent model of cerebral ischemia–reperfusion injury by GC–MS based metabolomic profiling. J. Pharmaceut. Biomed. Anal. 2017, 142, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, X.; Wang, H.; Xiao, H.; Jing, F.; Tang, L.; Li, D.; Zhang, Y.; Wu, H.; Yang, H. Metabolomics study on the effects of Buchang Naoxintong capsules for treating cerebral ischemia in rats using UPLC-Q/TOF-MS. J. Ethnopharmacol. 2016, 180, 1–11. [Google Scholar] [CrossRef]
- Kostova, I.; Dinchev, D. Saponins in Tribulus terrestris–chemistry and bioactivity. Phytochem. Rev. 2005, 4, 111–137. [Google Scholar] [CrossRef]
- Sucher, N.J. Insights from molecular investigations of traditional Chinese herbal stroke medicines: Implications for neuroprotective epilepsy therapy. Epilepsy Behav. 2006, 8, 350–362. [Google Scholar] [CrossRef]
- Jiang, E.-P.; Li, H.; Chen, J.-G.; Yang, S.-J. Protection by the gross saponins of Tribulus terrestris against cerebral ischemic injury in rats involves the NF-κB pathway. Acta Pharm. Sinica B 2011, 1, 21–26. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, S.; Hu, Y.; Li, P.; Wan, J.-B. Current state of the art of mass spectrometry-based metabolomics studies–a review focusing on wide coverage, high throughput and easy identification. RSC Adv. 2015, 5, 78728–78737. [Google Scholar] [CrossRef]
- Sévin, D.C.; Kuehne, A.; Zamboni, N.; Sauer, U. Biological insights through nontargeted metabolomics. Curr. Opin. Biotech. 2015, 34, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zaitsu, K.; Hayashi, Y.; Kusano, M.; Tsuchihashi, H.; Ishii, A. Application of metabolomics to toxicology of drugs of abuse: A mini review of metabolomics approach to acute and chronic toxicity studies. Drug Metab. Pharmacok. 2016, 31, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Van der Worp, H.B.; van Gijn, J. Acute ischemic stroke. New Engl. J. Med. 2007, 357, 572–579. [Google Scholar] [CrossRef]
- Hickey, J. Clinical Practice of Neurological & Neurosurgical Nursing; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- Dole, V.P. A relation between non-esterified fatty acids in plasma and the metabolism of glucose. J. Clin. Investig. 1956, 35, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Stinkens, R.; Goossens, G.H.; Jocken, J.W.; Blaak, E.E. Targeting fatty acid metabolism to improve glucose metabolism. Obes. Rev. 2015, 16, 715–757. [Google Scholar] [CrossRef]
- Rambold, A.S.; Cohen, S.; Lippincott-Schwartz, J. Fatty acid trafficking in starved cells: Regulation by lipid droplet lipolysis, autophagy, and mitochondrial fusion dynamics. Dev. Cell 2015, 32, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Olson, P.; Evans, R.M. Minireview: Lipid metabolism, metabolic diseases, and peroxisome proliferator-activated receptors. Endocrinology 2003, 144, 2201–2207. [Google Scholar] [CrossRef] [PubMed]
- Weisinger, H.S.; Armitage, J.A.; Sinclair, A.J.; Vingrys, A.J.; Burns, P.L.; Weisinger, R.S. Perinatal omega-3 fatty acid deficiency affects blood pressure later in life. Nat. Med. 2001, 7, 258–259. [Google Scholar] [CrossRef]
- Rizos, E.C.; Ntzani, E.E.; Bika, E.; Kostapanos, M.S.; Elisaf, M.S. Association between omega-3 fatty acid supplementation and risk of major cardiovascular disease events: A systematic review and meta-analysis. Jama 2012, 308, 1024–1033. [Google Scholar] [CrossRef]
- Siess, W.; Scherer, B.; Böhlig, B.; Roth, P.; Kurzmann, I.; Weber, P. Platelet-membrane fatty acids, platelet aggregation, and thromboxane formation during a mackerel diet. Lancet 1980, 315, 441–444. [Google Scholar] [CrossRef]
- Sanders, T.; Roshanai, F. The influence of different types of ω3 polyunsaturated fatty acids on blood lipids and platelet function in healthy volunteers. Clin. Sci. 1983, 64, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Iso, H.; Sato, S.; Umemura, U.; Kudo, M.; Koike, K.; Kitamura, A.; Imano, H.; Okamura, T.; Naito, Y.; Shimamoto, T. Linoleic acid, other fatty acids, and the risk of stroke. Stroke 2002, 33, 2086–2093. [Google Scholar] [CrossRef]
- Yamagishi, K.; Folsom, A.R.; Steffen, L.M.; Investigators, A.S. Plasma fatty acid composition and incident ischemic stroke in middle-aged adults: The atherosclerosis risk in communities (ARIC) study. Cerebrovas. Dis. 2013, 36, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Tilvis, R.; Erkinjuntti, T.; Sulkava, R.; Färkkilä, M.; Miettinen, T. Serum lipids and fatty acids in ischemic strokes. Am. Heart J. 1987, 113, 615–619. [Google Scholar] [CrossRef]
- Yaemsiri, S.; Sen, S.; Tinker, L.F.; Robinson, W.R.; Evans, R.W.; Rosamond, W.; Wasserthiel-Smoller, S.; He, K. Serum fatty acids and incidence of ischemic stroke among postmenopausal women. Stroke 2013, 44, 2710–2717. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Manson, J.E.; Willett, W.C. Types of dietary fat and risk of coronary heart disease: A critical review. J. Am. Coll. Nutr. 2001, 20, 5–19. [Google Scholar] [CrossRef]
- Ricci, S.; Celani, M.G.; Righetti, E.; Caruso, A.; De Medio, G.; Trovarelli, G.; Romoli, S.; Stragliotto, E.; Spizzichino, L.; Group, U.S. Fatty acid dietary intake and the risk of ischaemic stroke: A multicentre case-control study. J. Neurol. 1997, 244, 360–364. [Google Scholar] [CrossRef]
- Bucalossi, A.; Mori, S. Fatty acid composition of adipose tissue in ischemic heart disease and stroke. Gerontol. Clin. 1972, 14, 339–345. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Yu, S. Individual fatty acid effects on plasma lipids and lipoproteins: Human studies. Am. J. Clin. Nutr. 1997, 65, 1628S–1644S. [Google Scholar] [CrossRef]
- Neumar, R.W. Molecular mechanisms of ischemic neuronal injury. Ann. Emerg. Med. 2000, 36, 483–506. [Google Scholar] [CrossRef]
- Zhang, Z.-X.; Gao, P.-F.; Guo, X.-F.; Wang, H.; Zhang, H.-S. 1,3,5,7-Tetramethyl-8-(N-hydroxysuccinimidyl butyric ester) difluoroboradiaza-S-indacene as a new fluorescent labeling reagent for HPLC determination of amino acid neurotransmitters in the cerebral cortex of mice. Anal. Bioanal. Chem. 2011, 401, 1905–1914. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, J.; Zhang, C.; Liao, S.; Li, P.; Xu, D.; Lv, Y.; Yang, M.; Kong, L. The components of Huang-Lian-Jie-Du-Decoction act synergistically to exert protective effects in a rat ischemic stroke model. Oncotarget 2016, 7, 80872–80887. [Google Scholar] [CrossRef]
- Su, L.; Zhao, H.; Zhang, X.; Lou, Z.; Dong, X. UHPLC-Q-TOF-MS based serum metabonomics revealed the metabolic perturbations of ischemic stroke and the protective effect of RKIP in rat models. Mol. BioSyst. 2016, 12, 1831–1841. [Google Scholar] [CrossRef]
- Bie, X.-D.; Chen, Y.-Q.; Han, J.; Dai, H.-B.; Wan, H.-T.; Zhao, T.-F. Effects of gastrodin on amino acids after cerebral ischemia-reperfusion injury in rat striatum. Asia Pac. J. Clin. Nutr. 2007, 16, 305–308. [Google Scholar] [PubMed]
- Pettigrew, L.C.; Bang, H.; Chambless, L.E.; Howard, V.J.; Toole, J.F.; Investigators, V. Assessment of pre-and post-methionine load homocysteine for prediction of recurrent stroke and coronary artery disease in the Vitamin Intervention for Stroke Prevention Trial. Atherosclerosis 2008, 200, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Chiang, T.; Messing, R.O.; Chou, W.-H. Mouse model of middle cerebral artery occlusion. J. Visual. Exper. 2011, 13, e2761. [Google Scholar] [CrossRef]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar] [CrossRef]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoqlu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: Mass spectral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal. Chem. 2009, 24, 10038–10048. [Google Scholar] [CrossRef]
- Chen, Q.; Lu, X.; Guo, X.; Guo, Q.; Li, D. Metabolomics characterization of two Apocynaceae plants, Catharanthus roseus and Vinca minor, using GC-MS and LC-MS methods in combination. Molecules 2017, 22, 997. [Google Scholar] [CrossRef]
- Qiu, Y.; Cai, G.; Su, M.; Chen, T.; Zheng, X.; Xu, Y.; Ni, Y.; Zhao, A.; Xu, L.X.; Cai, S.; Jia, W. Serum metabolite profiling of human colorectal cancer using GC-TOFMS and UPLC-QTOFMS. J. Proteome Res. 2009, 8, 4844–4850. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Qiu, Y.; Ni, Y.; Su, M.; Jia, W.; Du, X. An automated data analysis pipeline for GC-TOF-MS metabonomics studies. J. Proteome Res. 2010, 9, 5974–5981. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Qiu, Y.; Jiang, W.; Suttlemyre, K.; Su, M.; Zhang, W.; Jia, W.; Du, X. ADAP-GC 2.0: Deconvolution of coeluting metabolites from GC/TOF-MS data for metabolomics studies. Anal. Chem. 2012, 84, 6619–6629. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Su, M.; Qiu, Y.; Jia, W.; Du, X. ADAP-GC 3.0: Improved peak detection and deconvolution of co-eluting metabolites from GC/TOF-MS data for metabolomics studies. Anal. Chem. 2016, 88, 8802–8811. [Google Scholar] [CrossRef]
Sample Availability: Gross saponins of Tribulus terrestris fruit are available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolites | Formula | HMDB ID | Classification | p-Value | |
|---|---|---|---|---|---|---|
| Model vs. Sham | GSTTF vs. Model | |||||
| 1 | 4-Hydroxy-proline | C5H9NO3 | 06055 | Amino Acid | ↑ ## | ↓ * |
| 2 | Allantoic acid | C4H8N4O4 | 01209 | Purine Derivative | ↑ # | ↓ * |
| 3 | Beta-Alanine | C3H7NO2 | 00056 | Amino Acid | ↑ ## | ↓ |
| 4 | Creatine | C4H9N3O2 | 00064 | Amino Acid | ↑ ## | ↓ * |
| 5 | Creatinine | C4H7N3O | 00562 | Amino Acid | ↑ ## | ↓ ** |
| 6 | Homocysteine | C4H9NO2S | 00742 | Amino Acid | ↑ ## | ↓ ** |
| 7 | Isoleucine | C6H13NO2 | 00172 | Amino Acid | ↑ ### | ↓ ** |
| 8 | Serine | C3H7NO3 | 00187 | Amino Acid | ↑ # | ↓ * |
| 9 | Methionine sulfoxide | C5H11NO3S | 02005 | Amino Acid | ↑ ## | ↓ |
| 10 | Ornithine | C5H12N2O2 | 00214 | Amino Acid | ↑ ### | ↓ |
| 11 | Arabitol | C5H12O5 | 00568 | Carbohydrates | ↑ # | ↓ * |
| 12 | Glucose | C6H12O6 | 00122 | Carbohydrates | ↑ # | ↓ * |
| 13 | Ribose | C5H10O5 | 00283 | Carbohydrates | ↑ # | ↓ * |
| 14 | Xylose | C5H10O5 | 00098 | Carbohydrates | ↑ ## | ↓ ** |
| 15 | Glucose 6-phosphate | C6H13O9P | 01401 | Carbohydrates | ↑ # | ↓ ** |
| 16 | Ribonolactone | C5H8O5 | 01900 | Carbohydrates | ↑ ## | ↓ |
| 17 | Arachidonic acid | C20H32O2 | 01043 | Fatty Acids | ↓ ### | ↑ *** |
| 18 | Caproic acid | C6H12O2 | 00535 | Fatty Acids | ↑ # | ↓ |
| 19 | Docosahexaenoic acid | C22H32O2 | 02183 | Fatty Acids | ↑ # | ↓ |
| 20 | Dodecanoic acid | C12H24O2 | 00638 | Fatty Acids | ↑ ## | ↓ |
| 21 | Heptadecanoic acid | C17H34O2 | 02259 | Fatty Acids | ↑ # | ↓ * |
| 22 | Linoleic acid | C18H32O2 | 00673 | Fatty Acids | ↑ ## | ↓ *** |
| 23 | Myristic acid | C14H28O2 | 00806 | Fatty Acids | ↑ ## | ↓ * |
| 24 | Myristoleic acid | C14H26O2 | 02000 | Fatty Acids | ↑ ### | ↓ ** |
| 25 | Oleic acid | C18H34O2 | 00207 | Fatty Acids | ↑ # | ↓ |
| 26 | Palmitic acid | C16H32O2 | 00220 | Fatty Acids | ↑ ## | ↓ * |
| 27 | Palmitoleic acid | C16H30O2 | 03229 | Fatty Acids | ↑ # | ↓ |
| 28 | Pentadecanoic acid | C15H30O2 | 00826 | Fatty Acids | ↑ ## | ↓ * |
| 29 | Stearic acid | C18H36O2 | 00827 | Fatty Acids | ↑ # | ↓ ** |
| 30 | Cholesterol | C27H46O | 00067 | Lipids | ↑ ## | ↓ |
| 31 | O-Phosphoethanolamine | C2H8NO4P | 00224 | Lipids | ↑ # | ↓ * |
| 32 | Pregnenolone | C21H32O2 | 00253 | Lipids | ↑ # | ↓ |
| 33 | Guanosine | C10H13N5O5 | 00133 | Nucleotide | ↑ ## | ↓ |
| 34 | Inosine | C10H12N4O5 | 00195 | Nucleotide | ↑ # | ↓ * |
| 35 | Pipecolic acid | C6H11NO2 | 00716 | Organic Acids | ↑ # | ↓ * |
| 36 | Malic acid | C4H6O5 | 00744 | Organic Acids | ↑ ### | ↓ * |
| 37 | Petroselinic acid | C18H34O2 | 02080 | Organic Acids | ↑ ## | ↓ |
| 48 | Picolinic acid | C6H5NO2 | 02243 | Organic Acids | ↑ # | ↓ * |
| 49 | Succinic acid | C4H6O4 | 00254 | Organic Acids | ↑ # | ↓ |
| 40 | Dopamine | C8H11NO2 | 00073 | Phenols | ↑ ### | ↓ *** |
| 41 | Pantothenic acid | C9H17NO5 | 00210 | Vitamin | ↑ # | ↓ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhao, H.; Liu, Y.; Guo, W.; Bao, Y.; Zhang, M.; Xu, T.; Xie, S.; Liu, X.; Xu, Y. GC-MS-Based Metabolomics to Reveal the Protective Effect of Gross Saponins of Tribulus terrestris Fruit against Ischemic Stroke in Rat. Molecules 2019, 24, 793. https://doi.org/10.3390/molecules24040793
Wang Y, Zhao H, Liu Y, Guo W, Bao Y, Zhang M, Xu T, Xie S, Liu X, Xu Y. GC-MS-Based Metabolomics to Reveal the Protective Effect of Gross Saponins of Tribulus terrestris Fruit against Ischemic Stroke in Rat. Molecules. 2019; 24(4):793. https://doi.org/10.3390/molecules24040793
Chicago/Turabian StyleWang, Yang, Hongyu Zhao, Yue Liu, Wenjun Guo, Yanru Bao, Manqi Zhang, Tunhai Xu, Shengxu Xie, Xinyu Liu, and Yajuan Xu. 2019. "GC-MS-Based Metabolomics to Reveal the Protective Effect of Gross Saponins of Tribulus terrestris Fruit against Ischemic Stroke in Rat" Molecules 24, no. 4: 793. https://doi.org/10.3390/molecules24040793
APA StyleWang, Y., Zhao, H., Liu, Y., Guo, W., Bao, Y., Zhang, M., Xu, T., Xie, S., Liu, X., & Xu, Y. (2019). GC-MS-Based Metabolomics to Reveal the Protective Effect of Gross Saponins of Tribulus terrestris Fruit against Ischemic Stroke in Rat. Molecules, 24(4), 793. https://doi.org/10.3390/molecules24040793