1. Introduction
Alarm over the impact of pesticides on the environment and human health is increasing year after year, and rigorous pesticide registration procedures have been introduced. Through Directive 2009/128/EC, the European Community has severely restricted the use of synthetic pesticides in plant protection, and the new regulations have reduced the number of chemicals available in agriculture [
1]. Novel pesticides, including natural product-based formulations, have been developed to counteract the evolution of resistance among plant pathogen and pest populations [
2].
Microorganisms biosynthesize thousands of compounds displaying different biological activities, e.g., acting as antibiotics, therapeutic agents, toxins, hormones, etc. [
3]. Thus, the exploitation of the microbial metabolome has become an important area of research to isolate novel natural products potentially useful for agricultural applications [
3]. The improvement of screening technologies can be used to extend the search of new microbial active metabolites, i.e., through (i) upgrading of fermentation techniques, (ii) the development of new methods for detection, and (iii) genetic manipulation to create mutants able to produce qualitatively or quantitatively different metabolites [
4].
Microorganisms have been extensively screened for antibiotic production; the main producers are fungi and bacteria, particularly the actinomycetes
Streptomyces spp. [
5]. A large number of manuscripts deal with microbial metabolites, which have demonstrated efficacy as crop protection agents, but only a few compounds have been marketed so far [
2]. Examples of commercially available microbial metabolites with insecticide activity are: (1) abamectin and anthelmintic produced by the soil-dwelling actinomycete
S. avermitilis; (2) milbemycin (also known as milbemectin), an insecticide and acaricide from
S. hygroscopicus subsp.
aureolacrimosus; (3) polynactins, secondary metabolites from the actinomycete
S. aureus, isolated and applied as a mixture of tetranactin, trinactin and dynactin; (4) spinosad, a secondary metabolite from the soil actinomycete
Saccharopolyspora spinosa [
2].
The interaction between some fungal strains and the plant establish a molecular cross-talk in which fungal metabolites can act as elicitors that activate the expression of genes involved in plant defence response, and promote the growth of the plant [
6]. Identification of new bioactive compounds may be obtained with a good microbial collection or isolating metabolites not expressed under standard laboratory conditions [
6,
7,
8].
In this work, we show the results of a screening aimed to isolate microbial metabolites with insecticidal activity. Various genera of fungi, including some entomopathogenic species, were grown in liquid culture and their major secondary metabolites were investigated. Bioassay-guided fractionation used to isolate the bioactive metabolites allowed the identification of seven known compounds and a new metabolite named chlamyphilone. This compound was fully characterized by spectrometric analysis and chemical derivatization. The fungal metabolites have been screened for insecticidal activity against
Acyrthosiphon pisum (Hemiptera: Aphididae), the pea aphid, whose high rate of increase makes it a useful model for screening [
9].
2. Results
The microbes used in the present study includes various genera of fungi identified according to morphological features and molecular analyses (rDNA-ITS and β-tubulin gene sequencing). In particular, 5 isolates belong to the Trichoderma genus (T. tomentosum F19, T. asperellum CINO1, T. harzianum M10, T. harzianum F53, T. velutinum F28) and 4 to Penicillium (P. chrysogenum F5, P. decumbens F29, P. griseofulvum F11, P. restrictum F55). Beauveria bassiana BB1, Chloridium virescens F57, Metarhizium anisopliae MA3, Paecilomyces lilacinus 100379, Pochonia chlamydosporia B and Talaromyces pinophilus F36CF are present as single species.
All fungi have been grown in static conditions and the culture filtrates were extracted exhaustively with ethyl acetate. The crude extracts were fractionated by column chromatography to provide a total of 79 organic fractions. The extracts and all the fractions were tested at different concentrations on
Acyrthosiphon pisum.
Table 1 reports the percentage of mortality at 72h of the active extracts/fractions obtained from different fungal isolates. Only extracts obtained from
P. lilacinus,
P. griseofulvum,
P. chlamydosporia,
B. bassiana,
M. anisopliae and
T. pinophilus showed significant insecticidal activity (data obtained from the other fungal strains are not showed). Moreover, the bioassay-guided fractionation indicated that only one fraction per strain was responsible of a significant aphid mortality after 72 h exposure (
Table 1).
Chemical characterization of the active fractions was obtained using NMR and/or LC-MS qTOF analyses. Fraction No. 3 of
P. lilacinus (showing 30% aphid mortality;
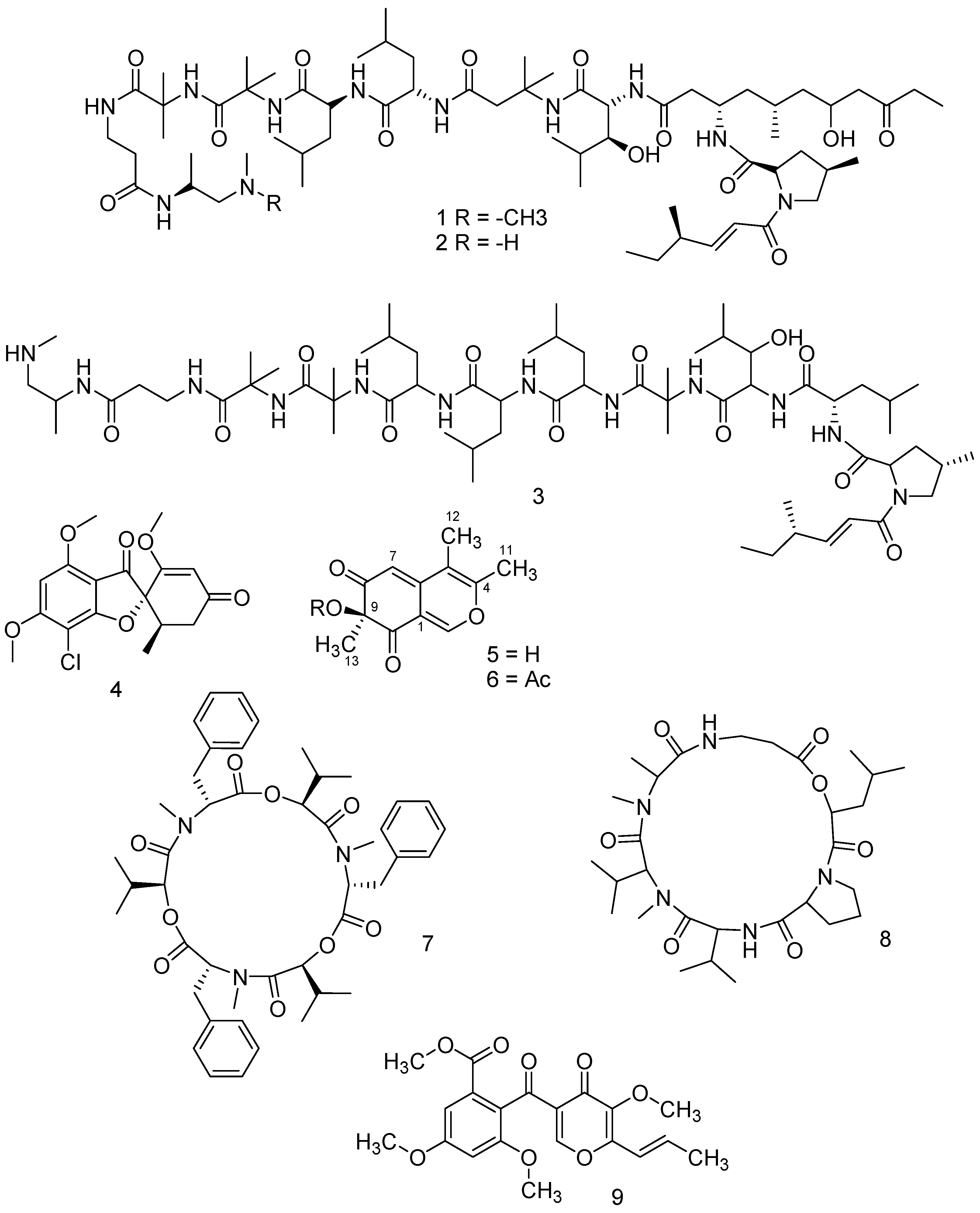
Table 1) was constituted by a mixture of leucinostatins A, B and D (
1,
2 and
3, respectively;
Figure 1), with the molecular weights (MW) 1218.634 g/mol, 1204.607 g/mol and 1104.4799 g/mol, respectively (
Figure S1).
The mycotoxin griseofulvin (
4 in
Figure 1) was determined as the main metabolite of
P. griseofulvum and was isolated in the active fraction number 7 (showing 73,3% aphid mortality;
Table 1).
Figure S3 reports the mass spectrum of the isolated compound.
The residue recovered after the organic extraction (350 mg) of
P. chlamydosporia culture filtrate was subjected to flash column chromatography, eluting with CH
2Cl
2/MeOH (90:10
v/
v). Fractions showing similar thin-layer chromatography (TLC) profiles were combined and further purified by using preparative TLC separation (Si gel; CH
2Cl
2/MeOH 90:10
v/
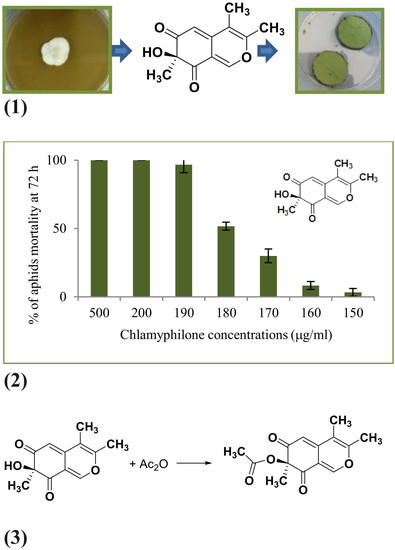
v). Both fractions and pure compounds were tested for insecticidal activity. Twelve milligrams of a novel compound, named chlamyphilone (
5 in
Figure 1; 5 mg/L), were obtained as a yellow amorphous solid in pure form after TLC (Rf in CH
2Cl
2/MeOH 90:10
v/
v = 0.8) and has: [α]
25 D −21.9° (c 1; CH
2Cl
2); UV (CH
2Cl
2) λ max (log ε) 219 (3.57), 332 (4.23).
1H and
13C NMR spectral data of chlamyphilone (in CDCl
3) are presented in
Table 2. LC-MS qTOF analysis detected the precursor ions at
m/
z 221.0814 (pseudomolecular ion [M + H]
+); 463.1360 [M
2 + Na]
+; 243.0916 [M + Na]
+; 221.0814 [M + H]
+ (calcd. 221.0814); 203.1313 [M + H − H
2O]
+ (
Figure S4). The results obtained by
13C NMR (
Table 2 and
Figure S6) and LC-MS qTOF analyses (
Figure S4) were consistent with a compound having a molecular weight of 220.0814, corresponding to the molecular formula C
12H
12O
4 with seven unsaturations.
The UV spectrum showed absorption peaks characteristic of the azaphilone family, which are natural products containing a 6
H-isochromene-6,8(7
H)-dione. Six carbon signals at 102.9, 114.4, 115.3, 145.2, 151.7, and 155.1 ppm revealed three double bonds, whereas signals at 196.3 and 196.5 ppm indicated the presence of two ketones. The other four signals in the
13C spectrum were all shifted upfield in the 12.7–83.11 ppm range. The DEPT data demonstrated that one of the protons in the molecule is bound to oxygen, and that in the molecule there are three CH
3, two CH, and seven fully substituted C atoms. The
1H-
1H COSY, HSQC analysis, and the chemical shift evaluation (
Table 2) allowed the identification of the structural fragment. In
Table 2 all the compound signals are reported.
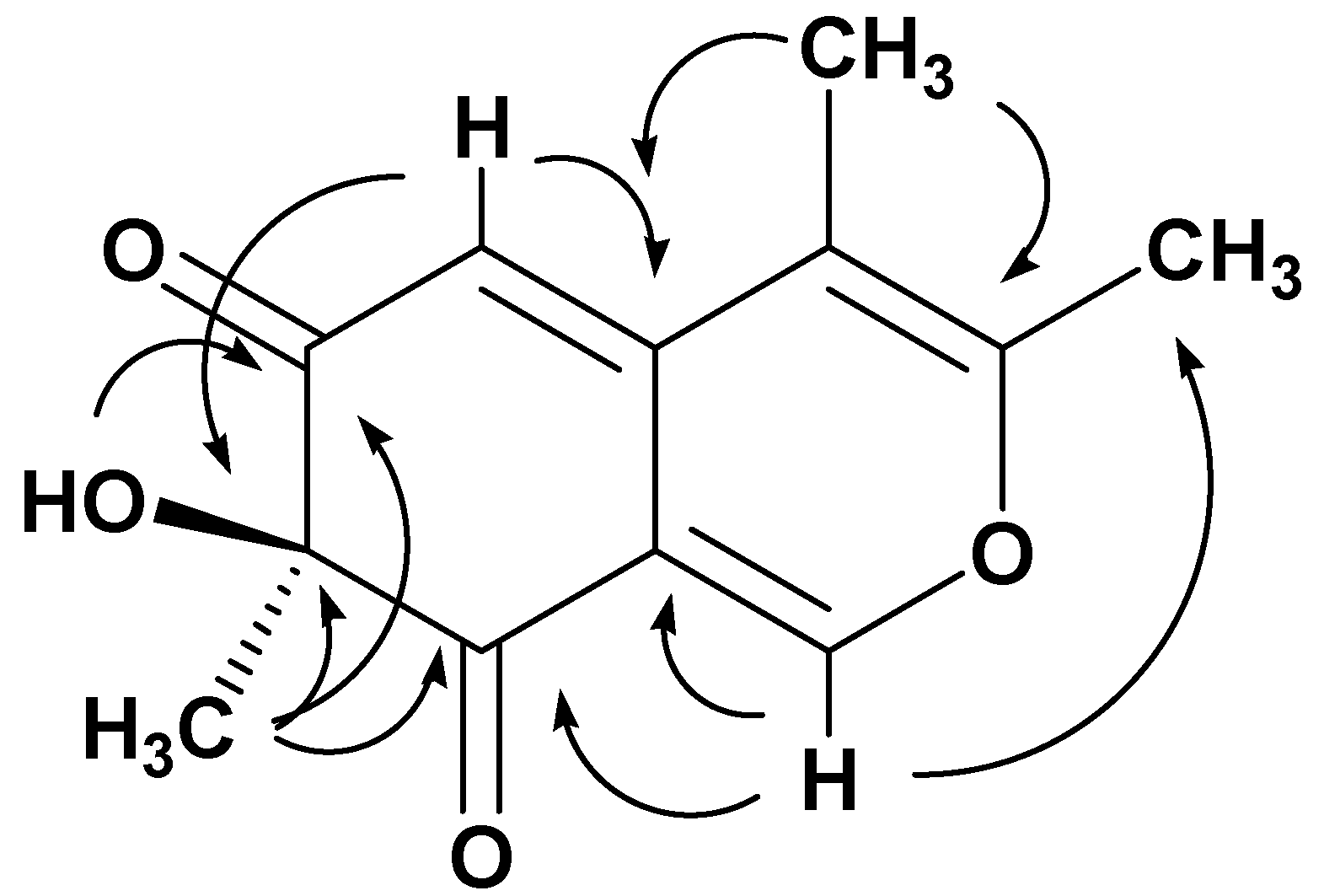
The connectivity of the spin systems was deduced by a long-range
1H-
13C heterocorrelated experiment that was obtained with the HMBC (
Table 2). In particular, the most relevant HMBC correlations are reported in
Figure 2, which implied that the structure of the metabolite
5 is 7-hydroxy-3,4,7-trimethyl-isochromene-6,8-dione. The configuration of chlamyphilone was evident from NOESY experiment. The MM-2 energy calculation (16.0 Kcal/mol) was run to find the most stable conformational model (
Figure S12).
To confirm the proposed structure, a sample of chlamyphilone was acetylated using acetic anhydride/pyridine (
6 in
Figure 1). The
1H-NMR spectrum of the product (acetic acid 3,4,7-trimethyl-6,8-dioxo-7,8-dihydro-6H-isochromen-7-yl ester) contained one acetate resonance, confirming the presence of the hydroxyl group bound to the carbon at 83.1. The
13C-NMR spectrum contained an additional signal for the acetyl carbonyl resonance at 170.8 and for the acetate methyl carbon at 18.4 ppm.
Chlamyphilone (
5 in
Figure 1) showed the highest insecticidal activity against
Acyrthosiphon pisum with a median Lethal Dose (LD50) of 175 μg/mL and a minimal inhibitory concentration (MIC) of 150 μg/mL (
Table 3).
The insecticidal activity of the
B. bassiana strain used in the present work was mainly dependent on the presence of beauvericin (
7 in
Figure 1). This metabolite is a cyclohexadepsipeptide mycotoxin with the molecular formula C
45H
57N
3O
9 (
Figure S13).
From
M. anisopliae extract, destruxin B2 has been characterized (
8 in
Figure 1). Destruxins are cyclic hexadepsipeptides composed of an α-hydroxy acid and five amino acid residues. Individual destruxins differ in terms of the hydroxy acid,
N-methylation, and R group of the amino acid residues. Destruxin B2 has the molecular formula C
29H
49N
5O
7, as revealed by the MS spectrum (
Figure S14).
Finally, the major active metabolite produced by
T. pinophilus strain F36CF was 3-
O-methylfunicone (OMF—
9 in
Figure 1), a well-known γ-pyrone derivative, previously isolated from
Talaromyces spp.
3. Discussion
In this work, a functional screening of fungal strains, preliminarily selected because of their ability to synthetize active metabolites, has been carried out. Bioassay-guided fractionation has been used to isolate microbial metabolites with insecticidal activity against the pea aphid
A. pisum (Hemiptera: Aphididae), long used as a model to study plant‒insect interactions [
9]. This aphid is an important pest of several plants, is a phloem-feeding insect determining direct negative effects on plants in terms of nutritive subtraction and injection of toxic saliva. Moreover, it has been involved in the transmission of diverse plant viruses [
9].
The active fraction obtained from
P. lilacinus was constituted by a mixture of leucinostatins A, B, and D. Leucinostatins are peptides containing an unsaturated fatty acid, nine amino acid residues, and a basic component joined together by amide linkages [
10]. These antibiotic metabolites also showed antitumor and nematocidal activities and an uncoupling effect on rat liver mitochondrial function [
10,
11]. To the best of our knowledge, our work is the first report on the insecticidal activity of leucinostatins against
A. pisum.
Griseofulvin was determined to be the main metabolite of
P. griseofulvum. The insecticidal activity of this metabolite has been previously demonstrated in the corn earworm,
Helicoverpa zea Boddie, and the fall armyworm,
Spodoptera frugiperda (J. E. Smith), by oral administration in an artificial diet (250 ppm) [
12,
13,
14]. In the case of soft-tegument aphids like
A. pisum, the activity is probably topical as oral administration may be excluded because of their peculiar feeding pattern. In fact, aphids use their piercing, sucking mouthparts to feed on plant fluid [
9].
The extract of
P. chlamydosporia culture filtrate yielded 12 mg of a novel compound named chlamyphilone. This metabolite belongs to the class of azaphilones, which contain a 6
H-isochromene-6,8(7
H)-dione or an isoquinoline-6,8(2
H,7
H)-dione skeleton (and its substituted derivatives thereof). This is the first report on the isolation of an azaphilone metabolite from
P. chlamydosporia. A similar azaphilone, named myxostiol, with plant growth regulating activity, has been previously isolated from
Myxotrichum stipitatum [
15]. Over 170 different azaphilone compounds, classified into 10 structural groups, have been isolated from fungi belonging to 23 genera [
16]. Azaphilones display a wide range of biological activities, such as antimicrobial, antifungal, antiviral, antioxidant, cytotoxic, nematocidal, and anti-inflammatory [
16]. Many of these properties may be explained by the reactions of azaphilones with amino groups, such as those found in amino acids, proteins, and nucleic acids, resulting in the formation of vinylogous c-pyridones [
16,
17,
18]. To the best of our knowledge, this is the first evidence of the insecticidal activity of a metabolite belonging to the class of azaphilones. Recently, the involvement of small molecules (e.g., aurovertins) in the interaction between nematodes and
P. chlamydosporia has been demonstrated [
19].
An active beauvericin has been extracted from culture filtrate of
B. bassiana. This metabolite is a mycotoxin whose insecticidal properties have been previously reported (e.g., against the wheat aphid
Schizaphis graminum at 0.5 mg/mL) [
20,
21]. This molecule contains three residues, each with D-2-hydroxyisovaleric acid (Hiv) and L-
N-methylphenylalanine linked alternately [
21].
Destruxin B2 is the active molecule isolated from
M. anisopliae extract. Destruxins exhibited a wide variety of biological activities, but are best known for their insecticidal and phytotoxic properties [
22,
23]. However, the effect of destruxin B2 against
Acyrthosiphon pisum is reported here for the first time.
T. pinophilus strain F36CF produced 3-
O-methylfunicone. This compound was downregulated in the presence of
T. harzianum M10 [
8], exhibited notable antibiotic and antitumor properties, and recently exhibited an insecticidal effect, thus expanding the biological activities of this compound [
24]. Recently, talarodiolide, a new 12-membered macrodiolide, was isolated and characterized from the culture filtrate of a
T. pinophilus strain [
25]. This metabolite did not show insecticidal activity, highlighting that the strain collection is important to select the best producers of bioactive compounds.
4. Materials and Methods
Fungal strains. The microbes used in the present study were present in the fungal collection at Department of Agricultural Sciences, University of Naples Federico II (UNINA Collection) or isolated from different sources (
Table 4).
The fungi were identified according to morphological features and molecular analyses. Briefly, an amount of 10
6 spores/mL was inoculated in a 250-mL Erlenmeyer Flasks containing 100 mL of sterile Potato Dextrose Broth (PDB, HIMEDIA, Mumbai, India). Each flask was incubated at 25 °C in an orbital shaker (150 r.p.m.) and fungi were left to grow for seven days. Mycelium was recovered, ground to a powder in liquid nitrogen, and used to perform the DNA extraction. Genetic analysis was carried out through PCR and sequencing of the rDNA Internal Transcribed Spacer (ITS) and β-tubulin gene, using the most common primers to identify fungal strains: ITS1 and ITS4 [
26], as well as tub2 and BenA [
27]. The PCR products were subjected to gel electrophoresis, excised, and sequenced [
26,
27]. Analysis of the ITS gave 99% of identity with GenBank sequences of the fungi reported in
Table 4, confirming the identity of the microbes.
Secondary metabolites production. The fungi were maintained on potato dextrose agar (PDA, HIMEDIA) at room temperature and sub-cultured bimonthly. Liquid cultures were prepared in 5000 L-Erlenmeyer flasks containing 1 L of PDB and inoculated with five mycelial plugs (7 mm
2) from a fresh PDA culture of each fungal strain. After 30 days of incubation at 25 °C in static conditions, the cultures were filtered through filter paper (Whatman No. 4) and the filtrate was analyzed by LC-MS qTOF [
28].
Extraction and isolation of fungal secondary metabolites. The culture filtrates were acidified to pH 4 with 5 M HCl and extracted exhaustively with ethyl acetate (EtOAc). The combined organic extracts were dried (Na
2SO
4) and solvent eliminated under reduced pressure at 35 °C (Rotavapor RV 10 IKA
® - Werke GmbH & Co. KG, Staufen, Germany). The residues recovered were fractionated by column chromatography (silica gel; 200 g) eluted with different eluents (
Table 5) and the homogeneous fractions were collected as reported in
Table 5. For thin layer chromatography (TLC) the following solvents were used: dichloromethane (CH
2Cl
2)/methanol (MeOH) 90:10; 80:20 (
v/
v), chloroform (CHCl
3)/MeOH 90:10; 80:20 (
v/
v); EtOAc/petroleum ether 90:10; 80:20 (
v/
v) [
28]. All fractions were tested for insecticidal activity against
A. pisum, and, where necessary, further purified by preparative TLC (
Table 5).
General experimental procedures. The isolated molecules were solubilised in 700 µL of deuterated chloroform (99.8% CDCl3 – Sigma-Aldrich, Darmstadt, Germany) and transferred into a stoppered NMR tube (5 mm, 7′′, 507-HP-7, NORELL, Morganton, NC, USA) where remaining void volume was gently degassed by a N2 flux. Proton and carbon solvent signals were used as reference to calibrate both 1H and 13C frequency axes. A 400 MHz Bruker Avance spectrometer (Bruker Co., Billerica, MA, USA), equipped with a 5 mm Bruker Broad Band Inverse probe (BBI), working at the 1H and 13C frequencies of 400.13 and 100.61 MHz, respectively, was used for the NMR measurements (at 25 ±1 °C).
Monodimensional 1H and 13C acquisitions were conducted as follows: proton spectra were acquired with 2 s of thermal equilibrium delay (number of scans = 64), a 90° pulse length 7.7 µs, 50 transients and 16 ppm (6410.2 Hz) as spectral widths, whereas proton-decoupled carbon acquisitions were executed by both inverse-gated and DEPT 135° pulse sequences, adopting 7 and 5 s of equilibrium delay, 12,500 and 2400 transients, respectively, and a spectral width of 250 ppm (25.152 KHz). A time domain of 32,768 points was adopted for all cited mono-dimensional experiments. Homo-nuclear 1H-1H COSY (COrrelation SpectroscopY), TOCSY (TOtal Correlation SpectroscopY), NOESY (Nuclear Overhauser Enhancement SpectroscopY), and hetero-nuclear 1H–13C HSQC (Hetero-nuclear Single-Quantum Correlation) and HMBC (Hetero-nuclear Multiple Bond Coherence) experiments (2D) were used for structural identification of metabolites. 2D homo- and heteronuclear spectra experiments were acquired with 48 and 80 scans, respectively, 16 dummy scans, a time domain of 2k points (F2) and 256 experiments (F1). TOCSY and NOESY experiments were conducted with a mixing time of 80 and 1000 ms, respectively, while HSQC and HMBC experiments were optimized for 145 Hz short and 6.5 Hz long range JCH couplings, respectively. All executed 2D experiments were gradient enhanced, except for the TOCSY acquisition. A Qsine weighting function associated to a magnitude mode was used to process NOESY spectrum with the purpose to emphasize the weak cross-peaks and minimize the noise artefacts. The free induction decay (FID) of mono-dimensional spectra was multiplied by an exponential factor corresponding to 0.1 Hz, for 1H and 13C acquisitions, and to 1 Hz for DEPT 135° experiment. All above mentioned spectra were baseline corrected and processed by using Bruker Topspin Software (v.4.0.2).
LC-MS/MS Q-TOF analysis were done on an Agilent HP 1260 Infinity Series liquid chromatograph equipped with a DAD system (Agilent Technologies, Santa Clara, CA, USA) coupled to a Q-TOF mass spectrometer model G6540B (Agilent Technologies). Separations were performed on a Zorbax Eclips Plus C18 column, 4.6 × 100 mm, with 3.5 µm particles (Agilent Technologies). The analyses were done at a constant temperature of 37 °C and using a linear gradient system composed of A: 0.1% (v/v) formic acid in water, and B: 0.1% (v/v) formic acid in acetonitrile. The flow was 0.6 mL/min, 95% A graduating to 100% B in 12 min, 100% B 12-15 min, 95% A 15-17 and equilibrating 95% A 17–20 min. The UV spectra were collected by DAD every 0.4 s from 190 to 750 nm with a resolution of 2 nm. The MS system was equipped with a Dual Electrospray Ionization (ESI) source and operated with Agilent MassHunter Data Acquisition Software, rev. B.05.01 in the positive or negative mode. Mass spectra were recorded in the range m/z 100–1600 as centroid spectra, with 3 scans per second. Two reference mass compounds were used to perform the real-time lock mass correction, purine (C5H4N4 at m/z 121.050873, 10 µmol/L) and hexakis (1H,1H, 3H-tetrafluoropentoxy)-phosphazene (C18H18O6N3P3F24 at m/z 922.009798, 2 µmol/L). The capillary was maintained at 4000 V, fragmentor voltage at 180 V, cone 1 (skimmer 1) at 45 V, Oct RFV at 750 V. Gas temperature was 350 °C during the run at 11 L/min, and the nebulizer was set at 45 psig. The injected sample volume was 5 µL.
MS/MS spectra were simultaneously recorded for confirmation purposes of new compounds, using the operating parameters described above, unless otherwise stated. The instrument was operated in the range m/z 100-1000, recording two spectra per second in targeted acquisition mode (targeted mass: 244.1197, Z = 1, RT 5.88 ±0.5 min). The sample collision energy was set to 20 V.
LC-MS data were evaluated using MassHunter Qualitative Analysis Software B.06.00 and compared to known compounds included in an in-house database. The database contains information of about 4000 known secondary metabolites isolated from more than 80 different fungal genera, and recorded according to their name, molecular formula, monoisotopic mass and producing organism. Positive identifications of fungal metabolites were reported if the compound was detected with a mass error below 10 ppm and with a sufficient score. Standards were used to confirm the chemical identifications.
UV spectra were recorded with a V-730 UV-Visible Spectrophotometer JASCO (Mary’s Court, Easton, MD, USA). Column chromatography was performed using silica gel (Merck silica gel 60 GF
254; Merck, Darmstadt, Germany), and TLC with glass pre-coated silica gel GF
254 plates (Merck Kieselgel 60 GF
254, 0.25 mm). The compounds were detected on TLC plates using UV light (254 or 366 nm) and/or by dipping the plates in a 5% (
v/
v) H
2SO
4 solution in ethanol followed by heating at 110 °C for 10 min [
28,
29].
Acetylation of chlamyphilone. Acetic anhydride (40 µL) was added to chlamyphilone (
1, 2.5 mg), dissolved in dry pyridine (80 µL 2.5 mg), and the residue was purified by preparative TLC on silica gel (petroleum ether/aceton, 20:80,
v/
v) to yield the acetyl derivative
6 (1.4 mg, 49%) [
29]. The reaction was monitored by LC-MS analysis (
Figure S1).
In vivo insecticidal assay. Insecticidal activity of organic extracts, each column fraction and pure metabolites were tested against the pea aphid
Acyrthosiphon pisum. Aphids were reared on
Vicia faba var.
aguadulce in a growth chamber (20 ± 1 °C, 70% RH, 18 h light/6 h dark photoperiod). Small populations were synchronized to obtain newborn nymphs every 24 h. Third-instar nymphs were dipped in the assay solution for 10 s and put on a paper towel to dry. Then, each insect was carefully transferred by a soft paintbrush on a leaf plug [
30,
31]. Twenty treated (or untreated—solvent control) 3rd-instar nymphs were placed on two circular (35 mm diameter) leaf plugs on 2% water agar layer in a Petri dish, in order to keep the leaf turgid. Plates were incubated at the climatic conditions described above and the number of dead aphids was assessed 24, 48, and 72 h after treatments. Extracts, fractions, and pure compounds were tested at 500, 400, 300, 200, 100, and 50 μg/mL. Each metabolite solution was also tested for phytotoxicity on
V. faba. The aphid mortality has been calculated in each treatment as the mean value ± standard deviation, in three replicates for each time point (24, 48, and 72 h). The experiments were repeated at least twice.
Statistical analysis. Data analysis was performed with SPSS 11.0 software (Statistics for Windows Version 24.0, IBM Corp., Armonk, NY, USA), and statistical analysis was done using one-way analysis of variance (ANOVA). The Least Significant Difference (LSD) post hoc test with p < 0.05 was used to analyse the multiple comparisons. The mean values between different treatments at the same time point and the mean values between the same treatments at different time points were compared.



 ,
, 


{kind=link}
{kind=link}
{kind=link}