Semi-Synthetic Ingenol Derivative from Euphorbia tirucalli Inhibits Protein Kinase C Isotypes and Promotes Autophagy and S-Phase Arrest on Glioma Cell Lines

 , ,
, ,  , ,
, , 
Abstract
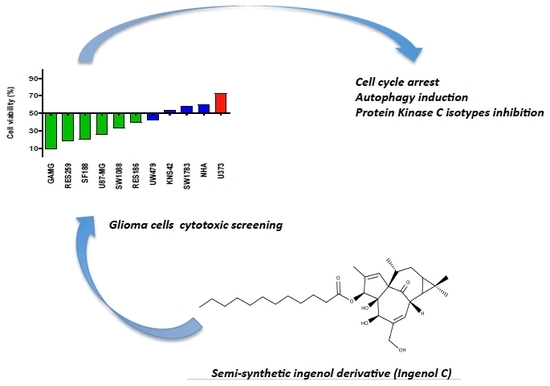
1. Introduction
2. Results
2.1. IngC promotes Cytotoxic Activity on Glioma Cell Lines More Effectively than Temozolomide but Their Combination Is Not Synergistic
2.2. IngC Exhibts Higher Cytotoxic Activity than Other Ingenol-Ester Class on Glioma Cells
2.3. Biological Properties of IngC in Cancer Cell Lines
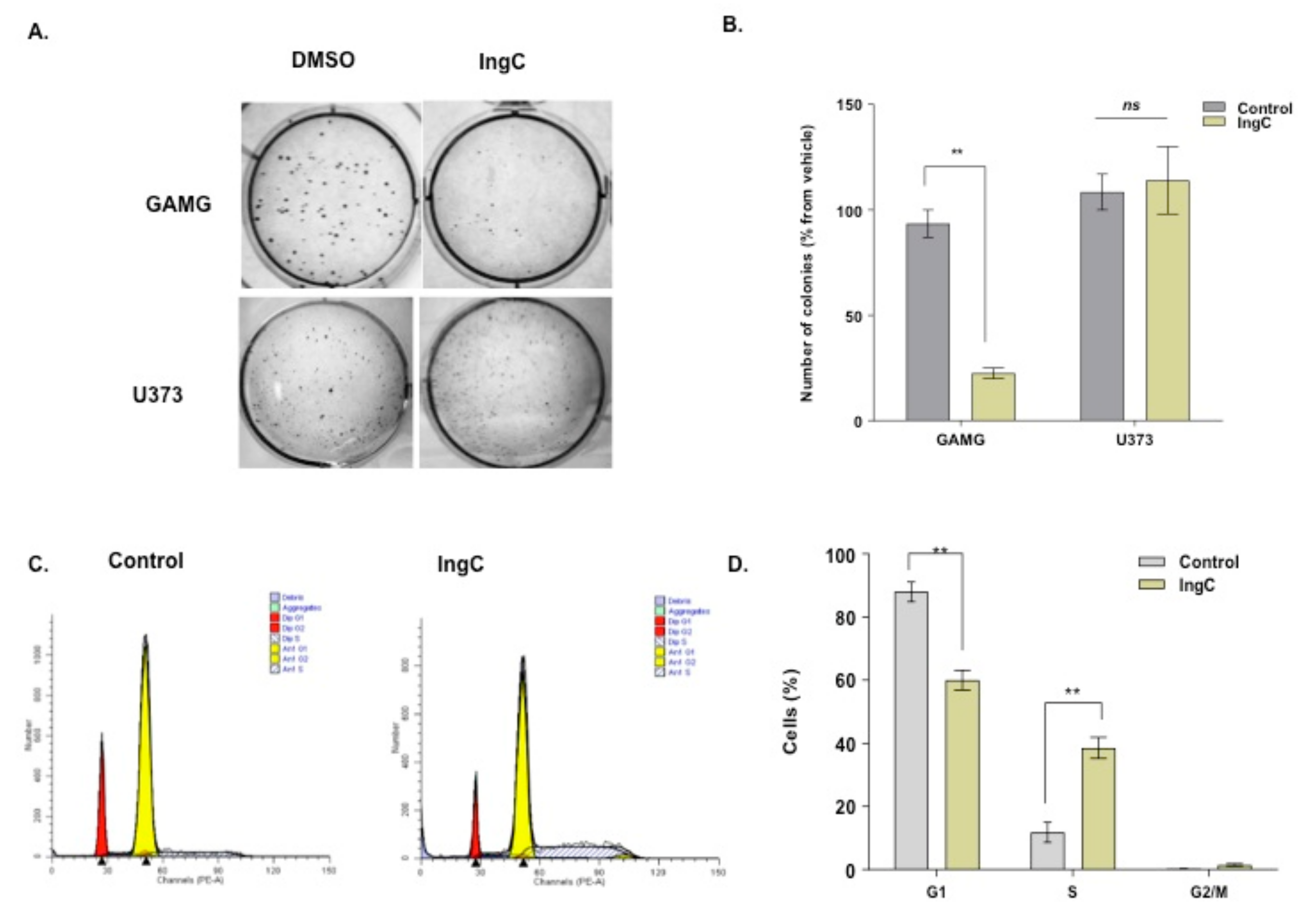
2.3.1. IngC Inhibits Proliferation and Induces S-Phase Arrest but Fails to Attenuate Migration and Invasion on Glioma Cells
2.3.2. IngC Induces Cell Death by Other Mechanisms, Not Apoptosis
2.3.3. IngC Induces Autophagy in Glioma Cells
2.3.4. IngC Exposure Inhibits Protein Kinase C Isotypes on Glioma Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Semi-Synthetic Ingenol
4.3. Cell Viability Analysis and IC50 Determination
4.4. Colony Formation-Assay
4.5. Migration and Invasion Assays
4.6. Cell Cycle and Cell Death Assays
4.7. Proteome Arrays
4.8. Western Blotting
4.9. Autophagy Analysis: LC3 Expression, Acidic Vesicular Organelles (AVOs) and IngC and Autophagy Inhibitor Combination Effect on Glioma Cell Lines
4.10. Drug Combination Studies
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AVOs | Acidic Vesicular Organelles |
| ANOVA | Analysis of variance |
| ATCC | American Type Culture Collection |
| Baf | bafilomycin A1 |
| CI | Combination index |
| DNA | Deoxyribonucleic acid |
| DSMZ | German Collection of Microorganisms and Cell Cultures |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | Dimethyl sulfoxide |
| ECACC | European Collection of Authenticated Cultures |
| FBS | Fetal bovine serum |
| FDA | Food and Drug Administration |
| GBM | Glioblastoma |
| GI | growth inhibition |
| g | Gram |
| IC50, | Half maximal inhibitory concentration |
| IngC, | ingenol-3-dodecanoate; I3A, ingenol-3-angelate |
| IDB | ingenol 3,20-dibenzoate |
| mL | Milliliter |
| MTS | 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H91tetrazolium) |
| NCI | National Cancer Institute |
| PKC | Protein kinase C |
| P/S | Penicillin/streptomycin solution |
| RPMI-1640 | Roswell Park Memorial Institute |
| SD | Standard deviation |
| STR | Short tandem repeat |
| TMZ | temozolamide |
| WHO | World Health Organization |
| uM | Micromolar |
References
- Weller, M.; Wick, W.; Aldape, K.; Brada, M.; Berger, M.; Pfister, S.M.; Nishikawa, R.; Rosenthal, M.; Wen, P.Y.; Stupp, R.; et al. Glioma. Nat. Rev. Dis Primers. 2015, 1, 15017. [Google Scholar] [CrossRef] [PubMed]
- Miranda-Filho, A.; Pineros, M.; Soerjomataram, I.; Deltour, I.; Bray, F. Cancers of the brain and CNS: Global patterns and trends in incidence. Neuro Oncol. 2017, 19, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Vecchione-Koval, T.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary brain and other central nervous system tumors diagnosed in the United States in 2010-2014. Neuro Oncol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; van den Bent, M.; Tonn, J.C.; Stupp, R.; Preusser, M.; Cohen-Jonathan-Moyal, E.; Henriksson, R.; Le Rhun, E.; Balana, C.; Chinot, O.; et al. European Association for Neuro-Oncology Task Force on, G., European Association for Neuro-Oncology (EANO) guideline on the diagnosis and treatment of adult astrocytic and oligodendroglial gliomas. Lancet Oncol. 2017, 18, 315–329. [Google Scholar] [CrossRef]
- Gladson, C.L.; Prayson, R.A.; Liu, W.M. The pathobiology of glioma tumors. Annu Rev. Pathol 2010, 5, 33–50. [Google Scholar] [CrossRef]
- Do Carmo, A.; Balca-Silva, J.; Matias, D.; Lopes, M.C. PKC signaling in glioblastoma. Cancer Biol. Ther. 2013, 14, 287–294. [Google Scholar] [CrossRef]
- Da Rocha, A.B.; Mans, D.R.; Regner, A.; Schwartsmann, G. Targeting protein kinase C: New therapeutic opportunities against high-grade malignant gliomas? Oncologist 2002, 7, 17–33. [Google Scholar] [CrossRef]
- Zellner, A.; Fetell, M.R.; Bruce, J.N.; De Vivo, D.C.; O’Driscoll, K.R. Disparity in expression of protein kinase C alpha in human glioma versus glioma-derived primary cell lines: Therapeutic implications. Clin. Cancer Res. 1998, 4, 1797–1802. [Google Scholar]
- Bredel, M.; Pollack, I.F. The role of protein kinase C (PKC) in the evolution and proliferation of malignant gliomas, and the application of PKC inhibition as a novel approach to anti-glioma therapy. Acta Neurochir. 1997, 139, 1000–1013. [Google Scholar] [CrossRef]
- Pollack, I.F.; DaRosso, R.C.; Robertson, P.L.; Jakacki, R.L.; Mirro, J.R., Jr.; Blatt, J.; Nicholson, S.; Packer, R.J.; Allen, J.C.; Cisneros, A.; et al. A phase I study of high-dose tamoxifen for the treatment of refractory malignant gliomas of childhood. Clin. Cancer Res. 1997, 3, 1109–1115. [Google Scholar]
- Muanza, T.; Shenouda, G.; Souhami, L.; Leblanc, R.; Mohr, G.; Corns, R.; Langleben, A. High dose tamoxifen and radiotherapy in patients with glioblastoma multiforme: A phase IB study. Can. J. Neurol. Sci. 2000, 27, 302–306. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schwartz, G.K.; Ward, D.; Saltz, L.; Casper, E.S.; Spiess, T.; Mullen, E.; Woodworth, J.; Venuti, R.; Zervos, P.; Storniolo, A.M.; et al. A pilot clinical/pharmacological study of the protein kinase C-specific inhibitor safingol alone and in combination with doxorubicin. Clin. Cancer Res. 1997, 3, 537–543. [Google Scholar] [PubMed]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. European Organisation for, R.; Treatment of Cancer Brain, T.; Radiotherapy, G.; National Cancer Institute of Canada Clinical Trials, G., Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Dutra, R.C.; Campos, M.M.; Santos, A.R.; Calixto, J.B. Medicinal plants in Brazil: Pharmacological studies, drug discovery, challenges and perspectives. Pharmacol. Res. 2016, 112, 4–29. [Google Scholar] [CrossRef]
- Dutra, R.C.; Bicca, M.A.; Segat, G.C.; Silva, K.A.; Motta, E.M.; Pianowski, L.F.; Costa, R.; Calixto, J.B. The antinociceptive effects of the tetracyclic triterpene euphol in inflammatory and neuropathic pain models: The potential role of PKCepsilon. Neuroscience 2015, 303, 126–137. [Google Scholar] [CrossRef]
- Passos, G.F.; Medeiros, R.; Marcon, R.; Nascimento, A.F.; Calixto, J.B.; Pianowski, L.F. The role of PKC/ERK1/2 signaling in the anti-inflammatory effect of tetracyclic triterpene euphol on TPA-induced skin inflammation in mice. Eur. J. Pharmacol. 2013, 698, 413–420. [Google Scholar] [CrossRef]
- Silva, V.A.O.; Rosa, M.N.; Tansini, A.; Oliveira, R.J.S.; Martinho, O.; Lima, J.P.; Pianowski, L.F.; Reis, R.M. In vitro screening of cytotoxic activity of euphol from Euphorbia tirucalli on a large panel of human cancer-derived cell lines. Exp. Ther. Med. 2018, 16, 557–566. [Google Scholar] [CrossRef]
- Silva, V.A.O.; Rosa, M.N.; Miranda-Goncalves, V.; Costa, A.M.; Tansini, A.; Evangelista, A.F.; Martinho, O.; Carloni, A.C.; Jones, C.; Lima, J.P.; et al. Euphol, a tetracyclic triterpene, from Euphorbia tirucalli induces autophagy and sensitizes temozolomide cytotoxicity on glioblastoma cells. Investig. New Drugs. 2019, 37, 223–237. [Google Scholar] [CrossRef]
- Silva, V.A.O.; Rosa, M.N.; Martinho, O.; Tanuri, A.; Lima, J.P.; Pianowski, L.F.; Reis, R.M. Modified ingenol semi-synthetic derivatives from Euphorbia tirucalli induce cytotoxicity on a large panel of human cancer cell lines. Investig. New Drugs 2019, 37, 1029–1035. [Google Scholar] [CrossRef]
- Abreu, C.M.; Price, S.L.; Shirk, E.N.; Cunha, R.D.; Pianowski, L.F.; Clements, J.E.; Tanuri, A.; Gama, L. Dual role of novel ingenol derivatives from Euphorbia tirucalli in HIV replication: Inhibition of de novo infection and activation of viral LTR. PLoS ONE 2014, 9, e97257. [Google Scholar] [CrossRef] [PubMed]
- Vasas, A.; Rédei, D.; Csupor, D.; Molnár, J.; Hohmann, J. Diterpenes from European Euphorbia Species Serving as Prototypes for Natural-Product-Based Drug Discovery. Eur. J. Org. Chem. 2012, 2012, 5115–5130. [Google Scholar] [CrossRef]
- Beres, T.; Dragull, K.; Pospisil, J.; Tarkowska, D.; Dancak, M.; Biba, O.; Tarkowski, P.; Dolezal, K.; Strnad, M. Quantitative Analysis of Ingenol in Euphorbia species via Validated Isotope Dilution Ultra-high Performance Liquid Chromatography Tandem Mass Spectrometry. Phytochem. Anal. 2018, 29, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Lebwohl, M.; Swanson, N.; Anderson, L.L.; Melgaard, A.; Xu, Z.; Berman, B. Ingenol mebutate gel for actinic keratosis. N. Engl. J. Med. 2012, 366, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Berman, B. New developments in the treatment of actinic keratosis: Focus on ingenol mebutate gel. Clin. Cosmet. Investig. Dermatol. 2012, 5, 111–122. [Google Scholar] [CrossRef]
- Gillespie, S.K.; Zhang, X.D.; Hersey, P. Ingenol 3-angelate induces dual modes of cell death and differentially regulates tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis in melanoma cells. Mol. Cancer Ther. 2004, 3, 1651–1658. [Google Scholar]
- Hampson, P.; Chahal, H.; Khanim, F.; Hayden, R.; Mulder, A.; Assi, L.K.; Bunce, C.M.; Lord, J.M. PEP005, a selective small-molecule activator of protein kinase C, has potent antileukemic activity mediated via the delta isoform of PKC. Blood 2005, 106, 1362–1368. [Google Scholar] [CrossRef]
- Wang, D.; Liu, P. Ingenol-3-Angelate Suppresses Growth of Melanoma Cells and Skin Tumor Development by Downregulation of NF-kappaB-Cox2 Signaling. Med. Sci. Monit. 2018, 24, 486–502. [Google Scholar] [CrossRef]
- Duarte, N.; Gyemant, N.; Abreu, P.M.; Molnar, J.; Ferreira, M.J. New macrocyclic lathyrane diterpenes, from Euphorbia lagascae, as inhibitors of multidrug resistance of tumour cells. Planta Med. 2006, 72, 162–168. [Google Scholar] [CrossRef]
- Vigone, A.; Tron, G.C.; Surico, D.; Baj, G.; Appendino, G.; Surico, N. Ingenol derivatives inhibit proliferation and induce apoptosis in breast cancer cell lines. Eur. J. Gynaecol. Oncol. 2005, 26, 526–530. [Google Scholar]
- Mochly-Rosen, D.; Das, K.; Grimes, K.V. Protein kinase C, an elusive therapeutic target? Nat. Rev. Drug Discov. 2012, 11, 937–957. [Google Scholar] [CrossRef] [PubMed]
- Paugh, B.S.; Qu, C.; Jones, C.; Liu, Z.; Adamowicz-Brice, M.; Zhang, J.; Bax, D.A.; Coyle, B.; Barrow, J.; Hargrave, D.; et al. Integrated molecular genetic profiling of pediatric high-grade gliomas reveals key differences with the adult disease. J. Clin. Oncol. 2010, 28, 3061–3068. [Google Scholar] [CrossRef] [PubMed]
- Suffness, M. Assays related to cancer drug discovery. Methods Plant. Biochem 1990, 6, 71–133. [Google Scholar]
- Blanco-Molina, M.; Tron, G.C.; Macho, A.; Lucena, C.; Calzado, M.A.; Munoz, E.; Appendino, G. Ingenol esters induce apoptosis in Jurkat cells through an AP-1 and NF-kappaB independent pathway. Chem. Biol. 2001, 8, 767–778. [Google Scholar] [CrossRef]
- Serova, M.; Ghoul, A.; Benhadji, K.A.; Faivre, S.; Le Tourneau, C.; Cvitkovic, E.; Lokiec, F.; Lord, J.; Ogbourne, S.M.; Calvo, F.; et al. Effects of protein kinase C modulation by PEP005, a novel ingenol angelate, on mitogen-activated protein kinase and phosphatidylinositol 3-kinase signaling in cancer cells. Mol. Cancer Ther. 2008, 7, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Lizarte Neto, F.S.; Tirapelli, D.P.; Ambrosio, S.R.; Tirapelli, C.R.; Oliveira, F.M.; Novais, P.C.; Peria, F.M.; Oliveira, H.F.; Carlotti Junior, C.G.; Tirapelli, L.F. Kaurene diterpene induces apoptosis in U87 human malignant glioblastoma cells by suppression of anti-apoptotic signals and activation of cysteine proteases. Braz. J. Med. Biol. Res. 2013, 46, 71–78. [Google Scholar] [CrossRef]
- Cavalcanti, B.C.; Costa-Lotufo, L.V.; Moraes, M.O.; Burbano, R.R.; Silveira, E.R.; Cunha, K.M.; Rao, V.S.; Moura, D.J.; Rosa, R.M.; Henriques, J.A.; et al. Genotoxicity evaluation of kaurenoic acid, a bioactive diterpenoid present in Copaiba oil. Food Chem. Toxicol. 2006, 44, 388–392. [Google Scholar] [CrossRef]
- Hasler, C.M.; Acs, G.; Blumberg, P.M. Specific binding to protein kinase C by ingenol and its induction of biological responses. Cancer Res. 1992, 52, 202–208. [Google Scholar]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef]
- Noonan, J.; Zarrer, J.; Murphy, B.M. Targeting Autophagy in Glioblastoma. Crit. Rev. Oncog. 2016, 21, 241–252. [Google Scholar] [CrossRef]
- Kanzawa, T.; Germano, I.; Komata, T.; Ito, H.; Kondo, Y.; Kondo, S. Role of autophagy in temozolomide-induced cytotoxicity for malignant glioma cells. Cell Death Differ. 2004, 11, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Kaza, N.; Kohli, L.; Roth, K.A. Autophagy in brain tumors: A new target for therapeutic intervention. Brain Pathol. 2012, 22, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Lomonaco, S.L.; Finniss, S.; Xiang, C.; Decarvalho, A.; Umansky, F.; Kalkanis, S.N.; Mikkelsen, T.; Brodie, C. The induction of autophagy by gamma-radiation contributes to the radioresistance of glioma stem cells. Int. J. Cancer 2009, 125, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Johannes, F.J.; Prestle, J.; Eis, S.; Oberhagemann, P.; Pfizenmaier, K. PKCu is a novel, atypical member of the protein kinase C family. J. Biol. Chem. 1994, 269, 6140–6148. [Google Scholar]
- Valverde, A.M.; Sinnett-Smith, J.; Van Lint, J.; Rozengurt, E. Molecular cloning and characterization of protein kinase D: A target for diacylglycerol and phorbol esters with a distinctive catalytic domain. Proc. Natl. Acad. Sci. USA 1994, 91, 8572–8576. [Google Scholar] [CrossRef]
- Patel, R.; Win, H.; Desai, S.; Patel, K.; Matthews, J.A.; Acevedo-Duncan, M. Involvement of PKC-iota in glioma proliferation. Cell Prolif. 2008, 41, 122–135. [Google Scholar] [CrossRef]
- Ersvaer, E.; Kittang, A.O.; Hampson, P.; Sand, K.; Gjertsen, B.T.; Lord, J.M.; Bruserud, O. The protein kinase C agonist PEP005 (ingenol 3-angelate) in the treatment of human cancer: A balance between efficacy and toxicity. Toxins 2010, 2, 174–194. [Google Scholar] [CrossRef]
- Kedei, N.; Lundberg, D.J.; Toth, A.; Welburn, P.; Garfield, S.H.; Blumberg, P.M. Characterization of the interaction of ingenol 3-angelate with protein kinase C. Cancer Res. 2004, 64, 3243–3255. [Google Scholar] [CrossRef]
- Kulkosky, J.; Sullivan, J.; Xu, Y.; Souder, E.; Hamer, D.H.; Pomerantz, R.J. Expression of latent HAART-persistent HIV type 1 induced by novel cellular activating agents. AIDS Res. Hum. Retroviruses 2004, 20, 497–505. [Google Scholar] [CrossRef]
- Balca-Silva, J.; Matias, D.; do Carmo, A.; Girao, H.; Moura-Neto, V.; Sarmento-Ribeiro, A.B.; Lopes, M.C. Tamoxifen in combination with temozolomide induce a synergistic inhibition of PKC-pan in GBM cell lines. Biochim. Biophys. Acta 2015, 1850, 722–732. [Google Scholar] [CrossRef]
- Cruvinel-Carloni, A.S.R.; Torrieri, R.; Bidinotto, L.T.; Berardinelli, G.N.; Oliveira-Silva, V.A.; Clara, C.A.; de Almeida, G.C.; Martinho, O.; Squire, J.A.; Reis, R.M. Molecular characterization of short-term primary cultures and comparison with corresponding tumor tissue of Brazilian glioblastoma patients. Transl. Cancer Res. 2017, 6, 332–345. [Google Scholar]
- Dirks, W.G.; Faehnrich, S.; Estella, I.A.; Drexler, H.G. Short tandem repeat DNA typing provides an international reference standard for authentication of human cell lines. ALTEX 2005, 22, 103–109. [Google Scholar]
- Konecny, G.E.; Glas, R.; Dering, J.; Manivong, K.; Qi, J.; Finn, R.S.; Yang, G.R.; Hong, K.L.; Ginther, C.; Winterhoff, B.; et al. Activity of the multikinase inhibitor dasatinib against ovarian cancer cells. Br. J. Cancer 2009, 101, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Martinho, O.; Silva-Oliveira, R.; Miranda-Goncalves, V.; Clara, C.; Almeida, J.R.; Carvalho, A.L.; Barata, J.T.; Reis, R.M. In Vitro and In Vivo Analysis of RTK Inhibitor Efficacy and Identification of Its Novel Targets in Glioblastomas. Transl. Oncol. 2013, 6, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Bruzzese, F.; Di Gennaro, E.; Avallone, A.; Pepe, S.; Arra, C.; Caraglia, M.; Tagliaferri, P.; Budillon, A. Synergistic antitumor activity of epidermal growth factor receptor tyrosine kinase inhibitor gefitinib and IFN-alpha in head and neck cancer cells in vitro and in vivo. Clin. Cancer Res. 2006, 12, 617–625. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | IngC IC50 ±S.D (μM) | I3A IC50 ±S.D (μM) | TMZ IC50 ±S.D (μM) | Combination Index (CI) *** TMZ+IngC | IngC Growth Inhibition in % at 10 μM * | IngC Growth Inhibition (GI) Score * | S.D | IngC SI ** | TMZ SI ** | Origin | Culture Conditions | Tumor Type |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| U87-MG | 4.02 ± 2.29 | 95.15 ± 14.35 | 746.76 ± 3.15 | 1.46 | 74.18 | HS | 10.46 | 1.85 | 0.15 | ATCC | DMEM + 10% FBS + 1% P/S | Adult glioma |
| U373 | 13.09± 0.84 | 76.39 ± 19.24 | 544.75 ± 1.53 | 0.80 | 26.88 | R | 8.19 | 0.57 | 0.20 | Kindly provided by Dr. Joseph Costello | DMEM + 10% FBS + 1% P/S | |
| GAMG | 0.19 ± 0.05 | 0.010 ± 0.13 | 97.00 ± 2.05 | UD | 90.58 | HS | 1.32 | 39.05 | 1.13 | DSMZ | DMEM + 10% FBS + 1% P/S | |
| SW1088 | 7.48 ± 0.47 | 87.62 ± 0.33 | 979.2 ± 4.00 | 1.20 | 66.64 | HS | 10.18 | 0.99 | 0.11 | ATCC | DMEM + 10% FBS + 1% P/S | |
| SW1783 | 7.4 ± 0.93 | 91.12 ± 1.59 | >1000 | 1.83 | 41.39 | MS | 0.63 | 1.00 | UD | ATCC | DMEM + 10% FBS + 1% P/S | |
| RES186 | 10.76 ± 2.6 | 89.56 ± 3.62 | 714.75 ± 7.08 | 1.35 | 60.4 | HS | 23.2 | 0.69 | 0.15 | kindly provided by Dr. Chris Jones | DMEM + 10% FBS + 1% P/S | Pediatric glioma |
| RES259 | 5.28 ± 1.54 | 78.15 ± 23.66 | 206.05 ± 6.09 | 1.13 | 81.4 | HS | 5.51 | 1.41 | 0.53 | kindly provided by Dr. Chris Jones | DMEM + 10% FBS + 1% P/S | |
| KNS42 | 8.10 ± 1.17 | 84.84 ± 34.86 | >1000 | 1.9 | 46.14 | MS | 5.07 | 0.92 | UD | kindly provided by Dr. Chris Jones | DMEM + 10% FBS + 1% P/S | |
| UW479 | 8.89 ± 0.86 | 72.63 ± 12.45 | >1000 | 1.2 | 57.57 | MS | 8.61 | 0.83 | UD | kindly provided by Dr. Chris Jones | DMEM + 10% FBS + 1% P/S | |
| SF188 | 3.38 ± 1.24 | 0.039±0.02 | >1000 | 1.80 | 80.24 | HS | 2.8 | 2.20 | UD | kindly provided by Dr. Chris Jones | DMEM + 10% FBS + 1% P/S | |
| HCB2 | 11.79± 1.04 | ND | ND | ND | 59.4 | MS | ND | 0.63 | ND | Barretos Cancer Hospital | DMEM + 10% FBS + 1% P/S | Primary culture |
| HCB149 | 20.16± 1.34 | ND | ND | ND | 20.3 | R | ND | 0.37 | ND | Barretos Cancer Hospital | DMEM + 10% FBS + 1% P/S | |
| NHA | 7.42 ± 2.46 | 37.59 ± 8.34 | 110.5 ± 1.05 | ND | 59.77 | MS | ND | ECACC | DMEM + 10% FBS + 1% P/S | Normal Human Astrocyte |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.A.O.; Rosa, M.N.; Tansini, A.; Martinho, O.; Tanuri, A.; Evangelista, A.F.; Cruvinel Carloni, A.; Lima, J.P.; Pianowski, L.F.; Reis, R.M. Semi-Synthetic Ingenol Derivative from Euphorbia tirucalli Inhibits Protein Kinase C Isotypes and Promotes Autophagy and S-Phase Arrest on Glioma Cell Lines. Molecules 2019, 24, 4265. https://doi.org/10.3390/molecules24234265
Silva VAO, Rosa MN, Tansini A, Martinho O, Tanuri A, Evangelista AF, Cruvinel Carloni A, Lima JP, Pianowski LF, Reis RM. Semi-Synthetic Ingenol Derivative from Euphorbia tirucalli Inhibits Protein Kinase C Isotypes and Promotes Autophagy and S-Phase Arrest on Glioma Cell Lines. Molecules. 2019; 24(23):4265. https://doi.org/10.3390/molecules24234265
Chicago/Turabian StyleSilva, Viviane Aline Oliveira, Marcela Nunes Rosa, Aline Tansini, Olga Martinho, Amilcar Tanuri, Adriane Feijó Evangelista, Adriana Cruvinel Carloni, João Paulo Lima, Luiz Francisco Pianowski, and Rui Manuel Reis. 2019. "Semi-Synthetic Ingenol Derivative from Euphorbia tirucalli Inhibits Protein Kinase C Isotypes and Promotes Autophagy and S-Phase Arrest on Glioma Cell Lines" Molecules 24, no. 23: 4265. https://doi.org/10.3390/molecules24234265
APA StyleSilva, V. A. O., Rosa, M. N., Tansini, A., Martinho, O., Tanuri, A., Evangelista, A. F., Cruvinel Carloni, A., Lima, J. P., Pianowski, L. F., & Reis, R. M. (2019). Semi-Synthetic Ingenol Derivative from Euphorbia tirucalli Inhibits Protein Kinase C Isotypes and Promotes Autophagy and S-Phase Arrest on Glioma Cell Lines. Molecules, 24(23), 4265. https://doi.org/10.3390/molecules24234265