Effects of Melatonin Treatment of Postharvest Pear Fruit on Aromatic Volatile Biosynthesis

 and
and
Abstract
:1. Introduction
2. Results
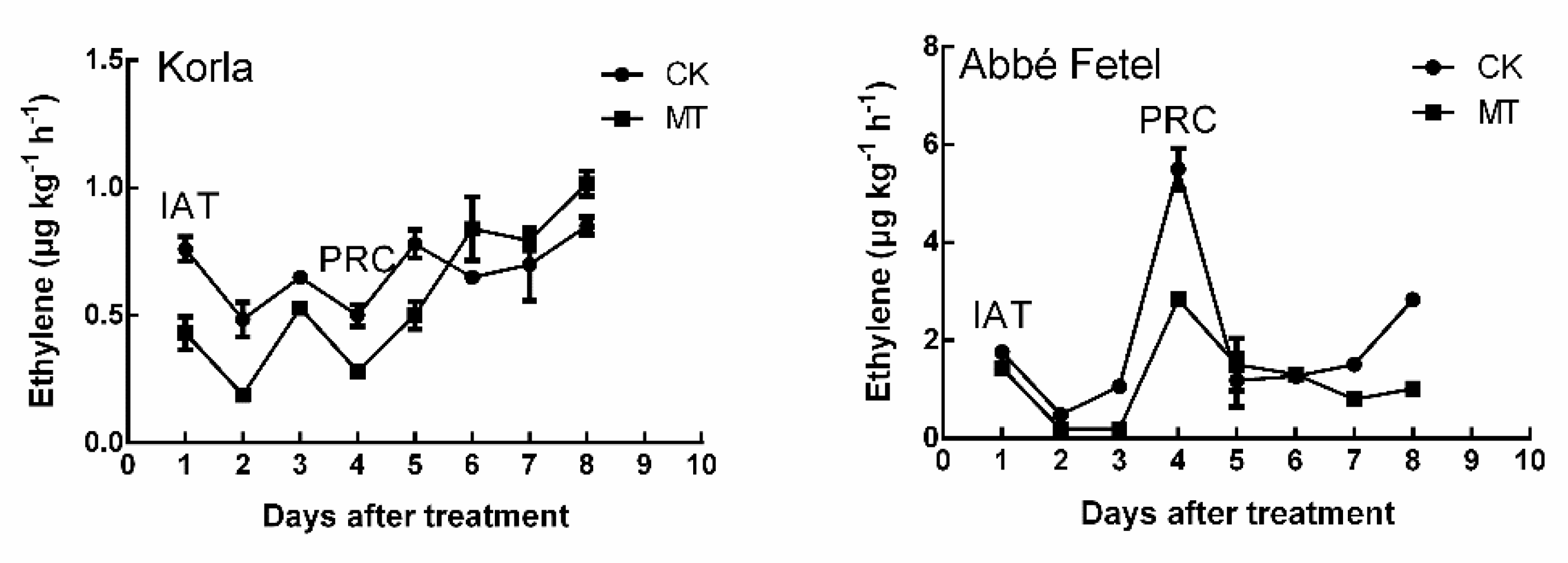
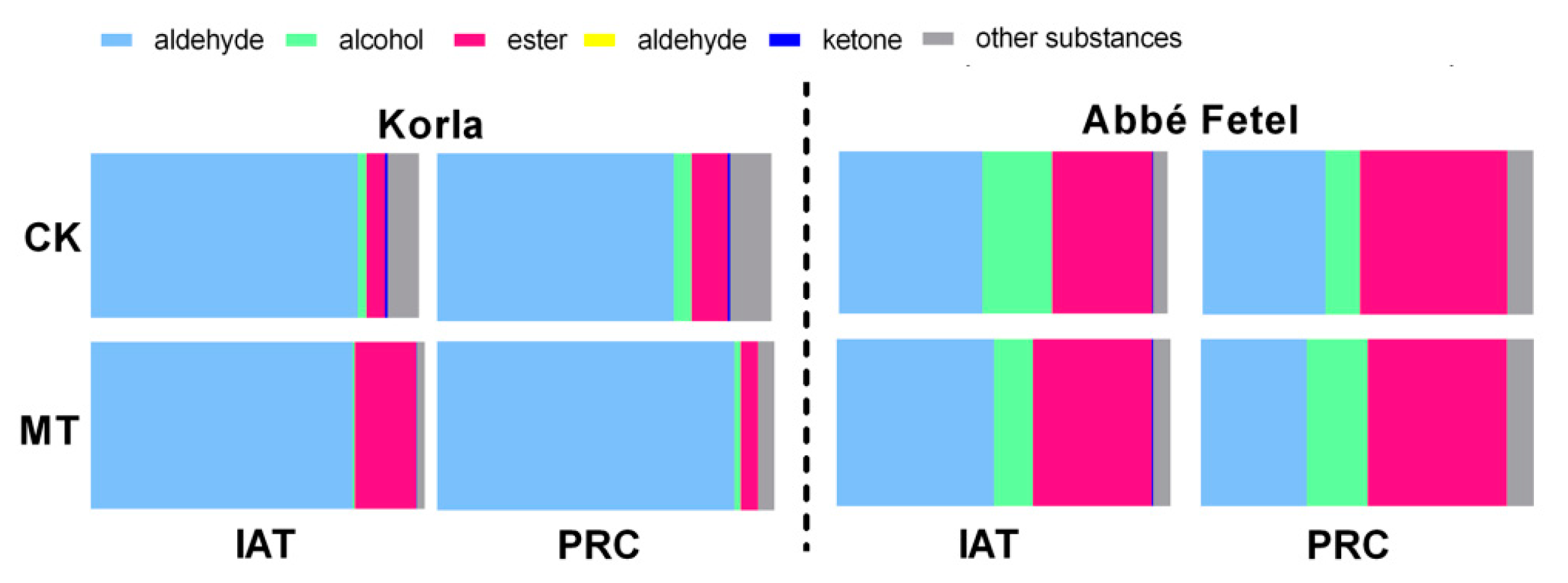
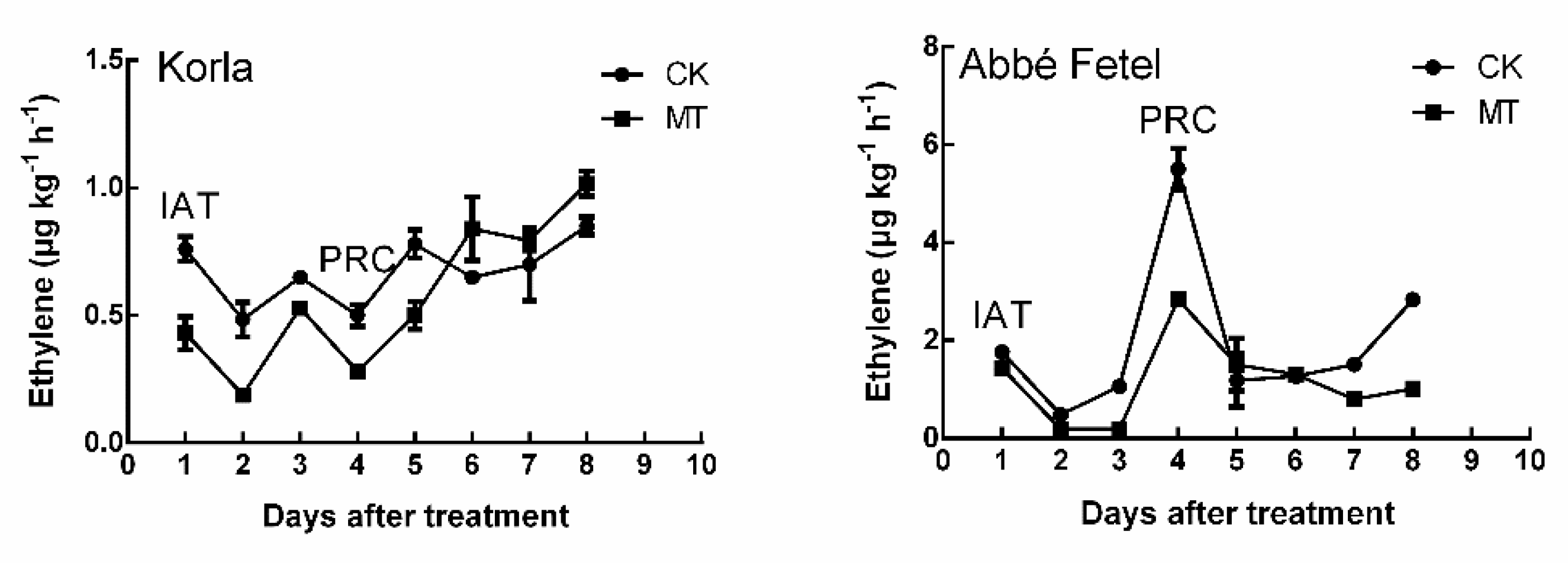
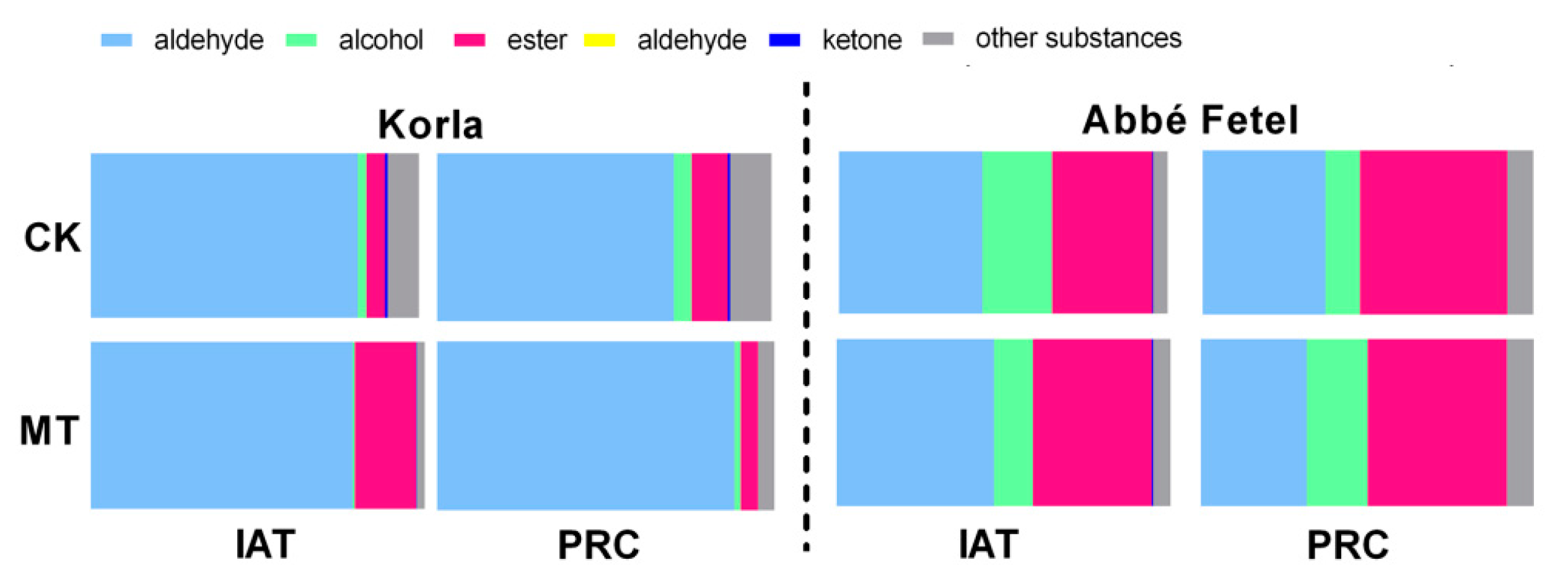
2.1. Effects of MT on Volatile Production during Postharvest Ripening
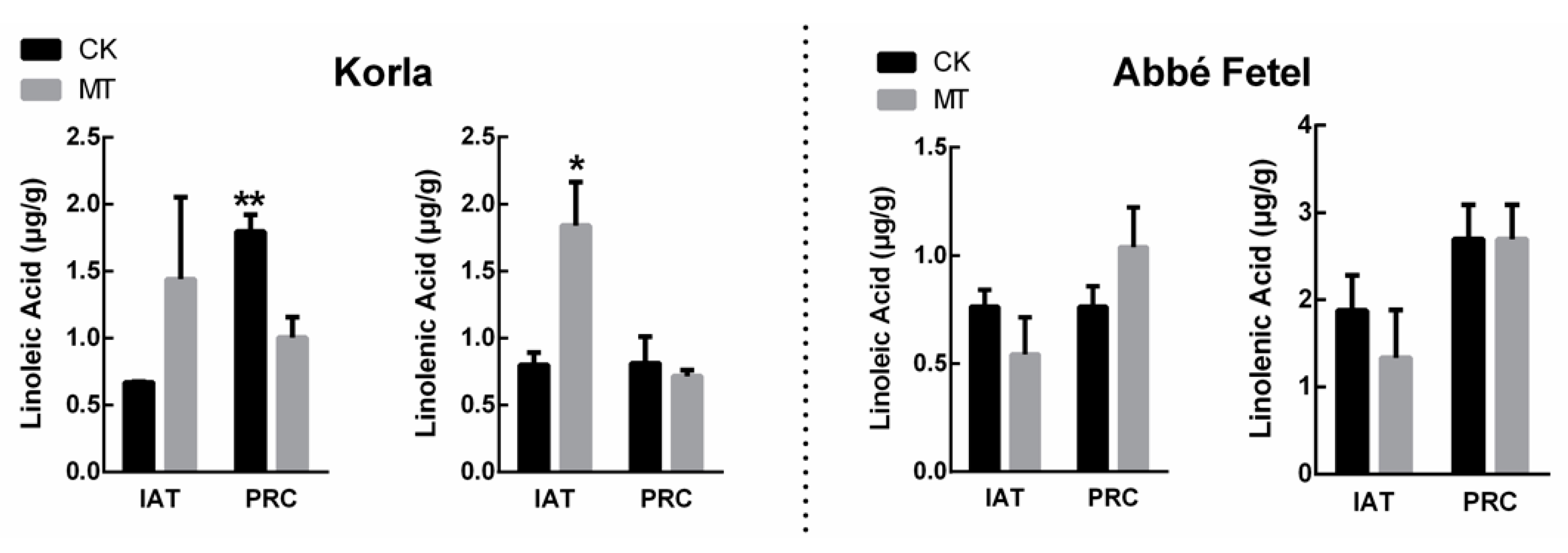
2.2. Effects of MT on Linoleic Acid and Linolenic Acid Content
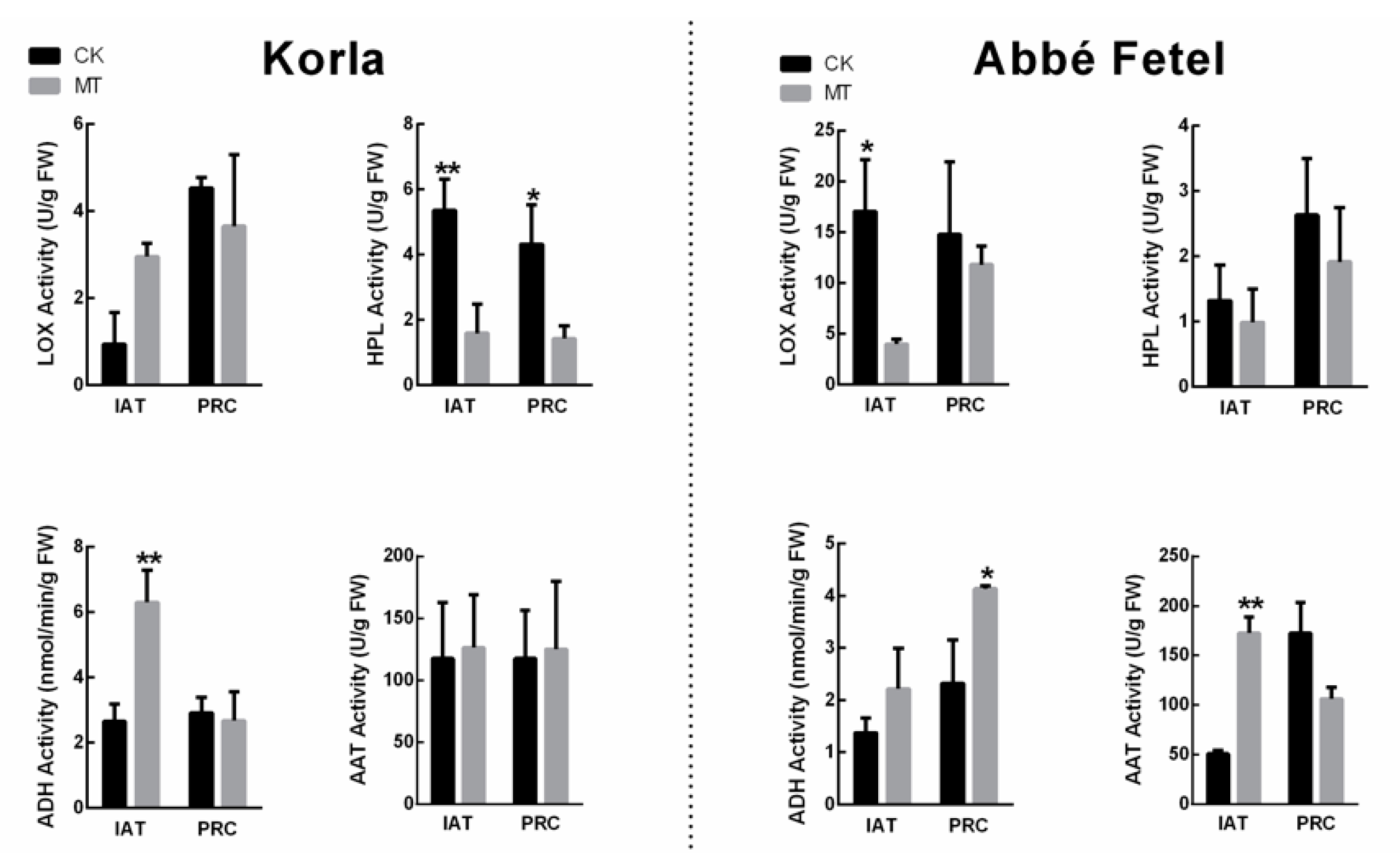
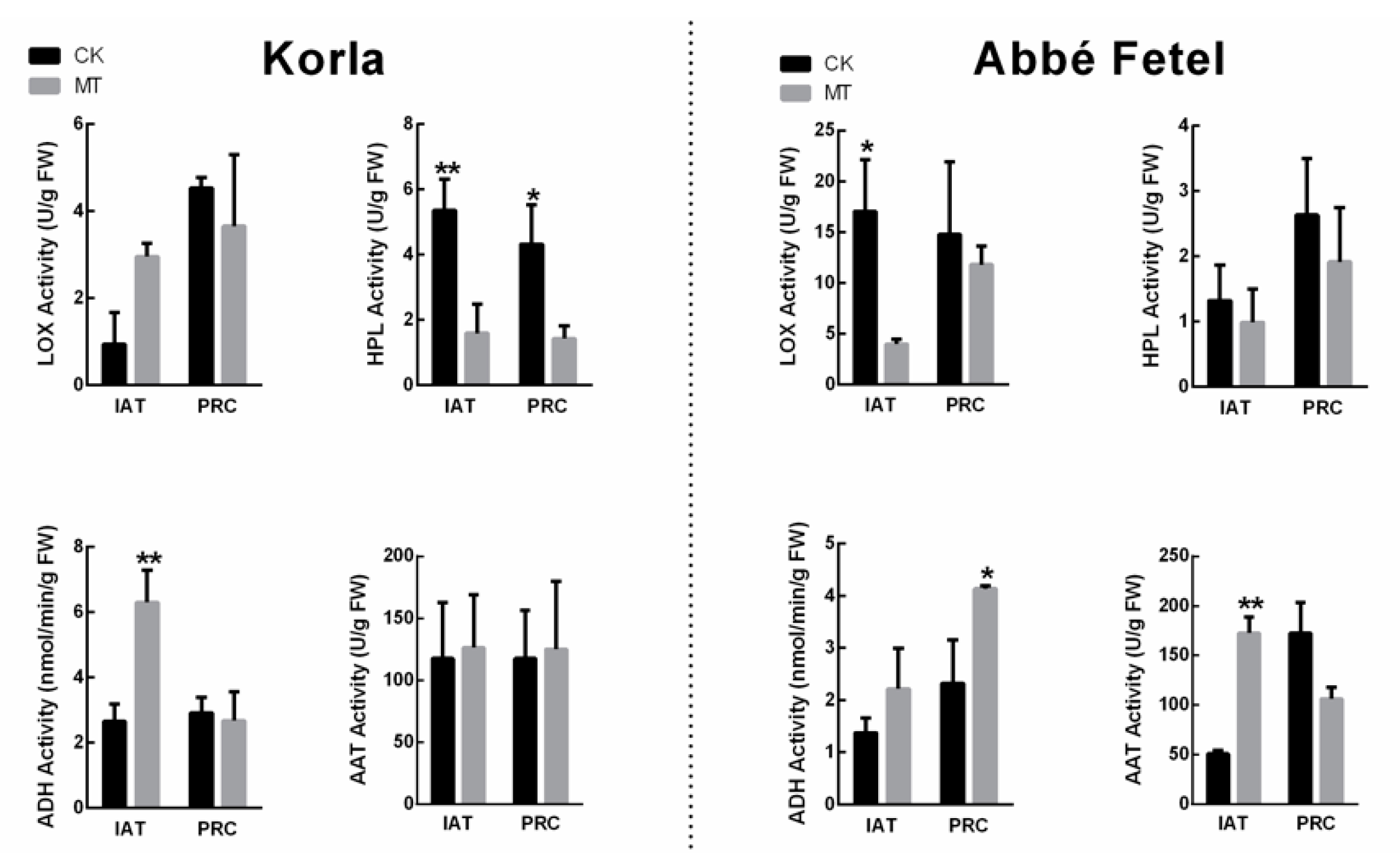
2.3. Effects of MT on Ester-related Enzyme Activity
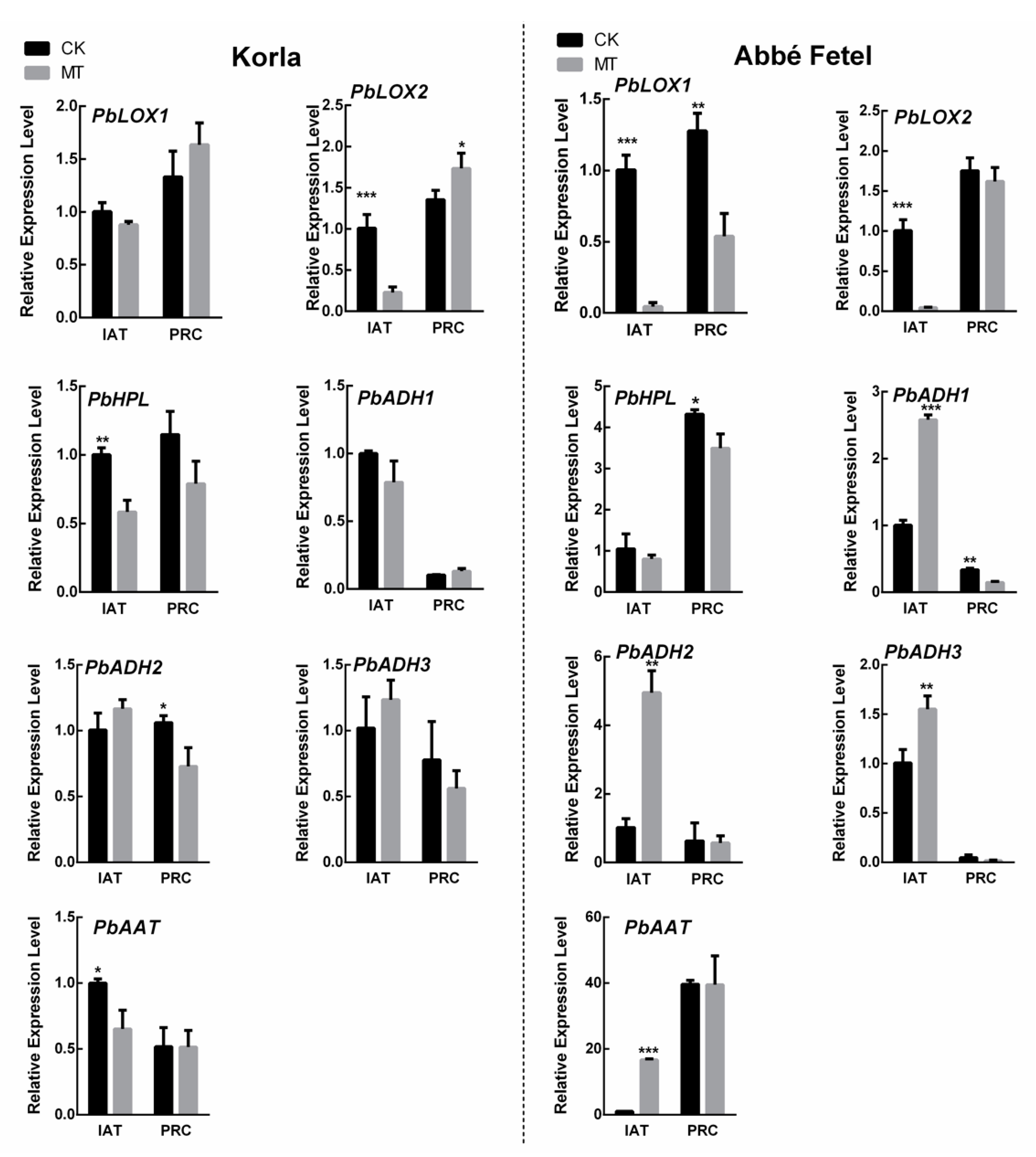
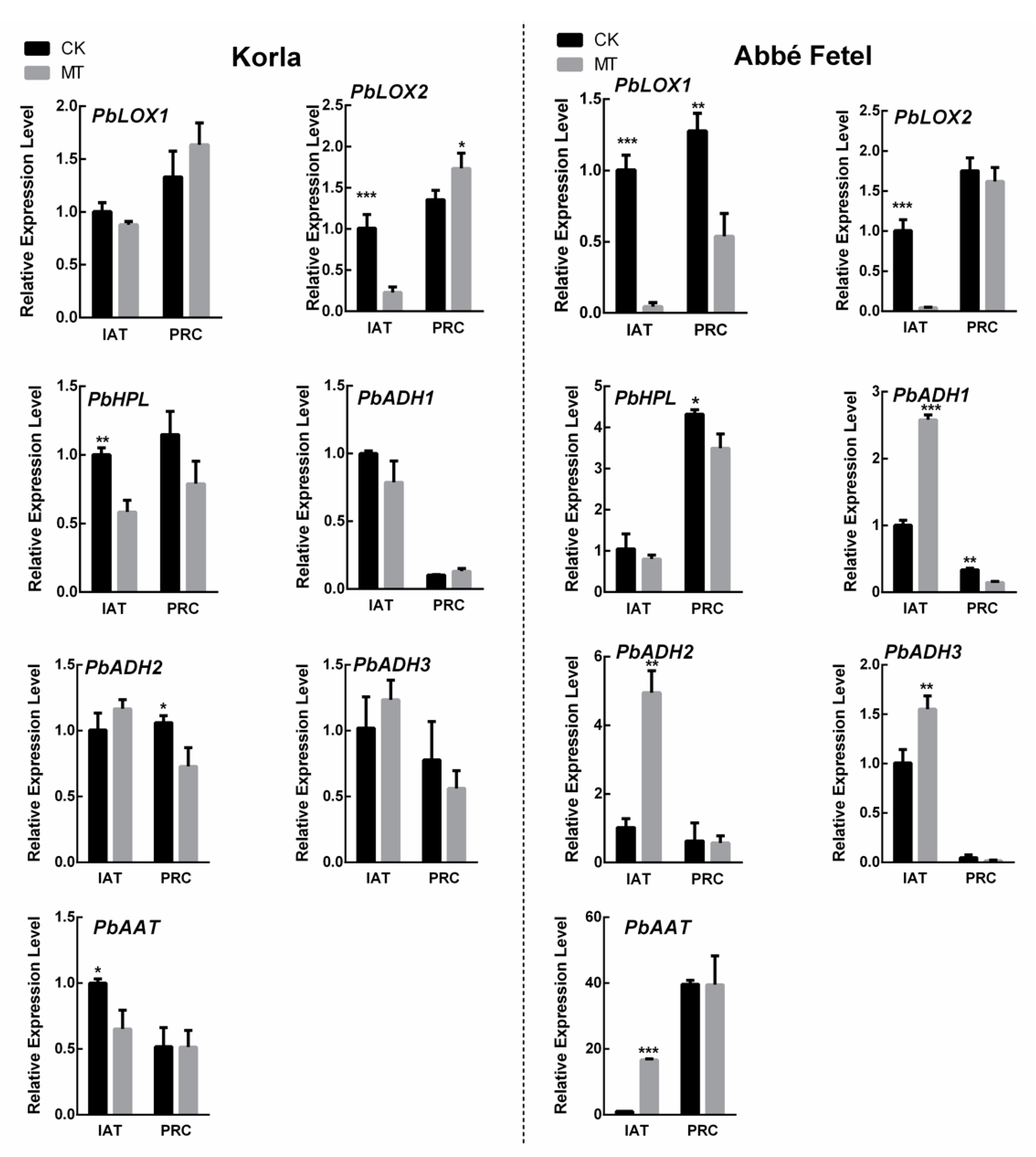
2.4. Effects of Melatonin on Expression of Ester-related Genes
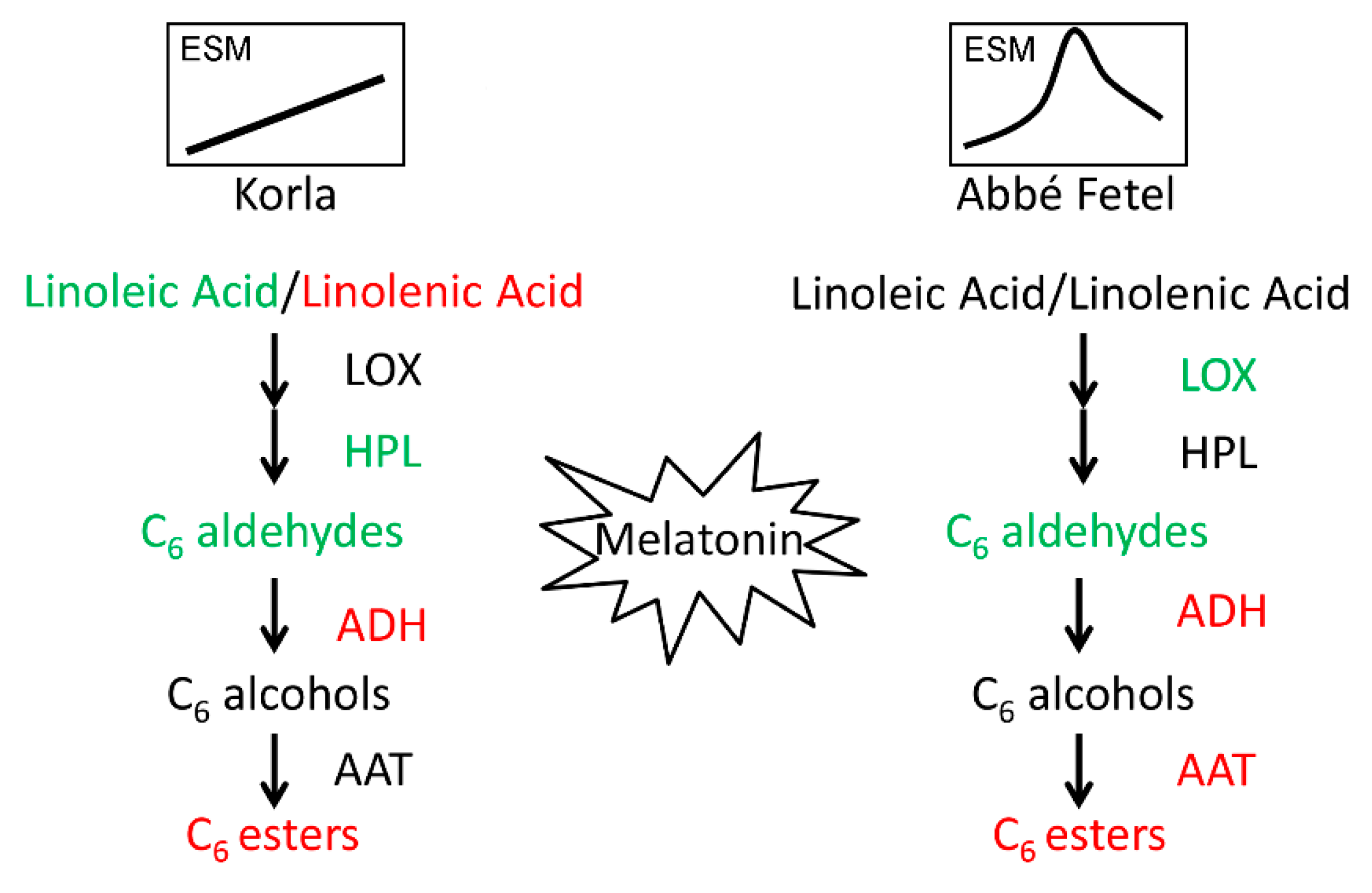
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Fruit Volatiles
4.3. Ethylene Production
4.4. Linoleic and Linolenic Acid Content
4.5. Enzyme Activity
4.6. Gene Expression
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A new plant hormone and/or a plant master regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, M.K.; Yu, J.; Zhou, J. Phytomelatonin: Recent advances and future prospects. J. Pineal Res. 2018, 65, e12526. [Google Scholar] [CrossRef]
- Sharif, R.; Xie, C.; Zhang, H.; Arnao, M.B.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Azher, N.M.; Chen, P.; et al. Melatonin and its effects on plant systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef]
- Wang, Y.; Reiter, R.J.; Chan, Z. Phytomelatonin: A universal abiotic stress regulator. J. Exp. Bot. 2018, 69, 963–974. [Google Scholar] [CrossRef]
- Zhai, R.; Liu, J.; Liu, F.; Zhao, Y.; Liu, L.; Fang, C.; Wang, H.; Li, X.; Wang, Z.; Ma, F.; et al. Melatonin limited ethylene production, softening and reduced physiology disorder in pear (Pyrus communis L.) fruit during senescence. Postharvest Biol. Technol. 2018, 139, 38–46. [Google Scholar] [CrossRef]
- Meng, J.; Yu, Y.; Shi, T.; Fu, Y.; Zhao, T.; Zhang, Z. Melatonin treatment of pre-veraison grape berries modifies phenolic components and antioxidant activity of grapes and wine. Food Sci. Tech-Braz. 2018. [Google Scholar] [CrossRef]
- Hu, W.; Tie, W.; Ou, W.; Yan, Y.; Kong, H.; Zuo, J.; Ding, X.; Ding, Z.; Liu, Y.; Wu, C. Crosstalk between calcium and melatonin affects postharvest physiological deterioration and quality loss in cassava. Postharvest Biol. Technol. 2018, 140, 42–49. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2014, 66, 657–668. [Google Scholar] [CrossRef]
- Zhang, B.; Yin, X.; Li, X.; Yang, S.; Ferguson, I.B.; Chen, K. Lipoxygenase gene expression in ripening kiwifruit in relation to ethylene and aroma production. J. Agric. Food Chem. 2009, 57, 2875–2881. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, R.J.; Friel, E.N.; Souleyre, E.J.F.; Bolitho, K.; Thodey, K.; Ledger, S.; Bowen, J.H.; Ma, J.H.; Nain, B.; Cohen, D.; et al. A genomics approach reveals that aroma production in apple is controlled by ethylene predominantly at the final step in each biosynthetic pathway. Plant Physiol. 2007, 144, 1899–1912. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.T.; Song, J.; Du, L.N.; Forney, C.; Leslie, C.P.; Sherry, F.; Wismer, P.; Zhang, Z.Q. Ethylene and 1-MCP regulate major volatile biosynthetic pathways in apple fruit. Food Chem. 2016, 194, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Lia, G.P.; Jia, H.J.; Li, J.H.; Li, H.X.; Teng, Y.W. Effects of 1-MCP on volatile production and transcription of ester biosynthesis related genes under cold storage in ‘Ruanerli’ pear fruit (Pyrus ussuriensis Maxim.). Postharvest Biol. Technol. 2016, 111, 168–174. [Google Scholar] [CrossRef]
- Wang, B.H.; Sun, X.X.; Dong, F.Y.; Zhang, F.; Niu, J.X. Cloning and expression analysis of an MYB gene associated with calyx persistence in Korla fragrant pear. Plant Cell Rep. 2014, 33, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. Cell Mol. Biol. 2008, 54, 712–732. [Google Scholar] [CrossRef]
- Manríquez, D.; El-Sharkawy, I.; Flores, F.B.; El-Yahyaoui, F.; Regad, F.; Bouzayen, M.; Latché, A.; Pech, J.C. Two highly divergent alcohol dehydrogenases of melon exhibit fruit ripening-specific expression and distinct biochemical characteristics. Plant Mol. Biol. 2006, 61, 675–685. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Z.; Chai, H.; Cheng, N.; Yang, Y.; Wang, D.; Yang, T.; Cao, W. Melatonin treatment delays postharvest senescence and regulates reactive oxygen species metabolism in peach fruit. Postharvest Biol. Technol. 2016, 118, 103–110. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of melatonin treatment on the postharvest quality of strawberry fruit. Postharvest Biol. Technol. 2018, 139, 47–55. [Google Scholar] [CrossRef]
- Zhang, Y.; Huber, D.J.; Hu, M.; Jiang, G.; Gao, Z.; Xu, X.; Jiang, Y.; Zhang, Z. Delay of postharvest browning in litchi fruit by melatonin via the enhancing of antioxidative processes and oxidation repair. J. Agric. Food Chem. 2018, 28, 7475–7484. [Google Scholar] [CrossRef]
- Liu, J.; Jie, Y.; Zhang, H.; Cong, L.; Zhai, R.; Yang, C.; Wang, Z.; Ma, F.; Xu, L. Melatonin inhibits ethylene synthesis via nitric oxide regulation to delay postharvest senescence in pears. J. Agric. Food Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.L.; Zhan, P.; Deng, Z.Y.; Yan, H.Y.; Zhu, X.R. Development of a flavour fingerprint by GC-MS and GC-O combined with chemometric methods for the quality control of Korla pear (Pyrus serotina Reld). Int. J. Food Sci. Tech. 2014, 12, 2546–2552. [Google Scholar] [CrossRef]
- Rapparini, F.; Predieris, S. Pear fruit volatiles. Hortic. Rev. 2003, 28, 237–324. [Google Scholar]
- Gomez-Lobato, M.E.; Civello, P.M.; Martínez, G.A. Expression of a lipoxygenase encoding gene (BoLOX1) during postharvest senescence of broccoli. Postharvest Biol. Technol. 2012, 64, 146–153. [Google Scholar] [CrossRef]
- EcheverrıÁ, G.; Graell, J.; López, M.L.; Lara, I. Volatile production, quality andaroma-related enzyme activities during maturation of ‘Fuji’ apples. Postharvest Biol. Technol. 2004, 31, 217–227. [Google Scholar] [CrossRef]
- Becerra Guerrero, J.; Rodríguez-Palacios, A. Functional characterization of enzymes forming volatile esters from strawberry and banana. Plant Physiol. 2004, 135, 1865. [Google Scholar]
- Souleyre, E.J.; Greenwood, D.R.; Friel, E.N.; Karunairetnam, S.; Newcomb, R.D. An alcohol acyl transferase from apple (cv. Royal Gala), MpAAT1, produces esters involved in apple fruit flavor. FEBS J. 2005, 272, 3132–3144. [Google Scholar] [CrossRef]
- Defilippi, B.G.; Kader, A.A.; Dandekar, A.M. Apple aroma: Alcohol acyltransferase, a rate limiting step for ester biosynthesis, is regulated by ethylene. Plant Sci. 2005, 168, 1199–1210. [Google Scholar] [CrossRef]
- Zhou, X.; Dong, L.; Zhou, Q.; Wang, J.; Chang, N.; Liu, Z.; Ji, S. Effects of intermittent warming on aroma-related esters of 1-methylcyclopropene-treated ‘Nanguo’ pears during ripening at room temperature. Sci. Hortic. 2015, 185, 82–89. [Google Scholar] [CrossRef]
- Zhang, B.; Shen, J.Y.; Wei, W.W.; Xi, W.P.; Xu, C.J.; Ferguson, I.; Chen, K. Expression of genes associated with aroma formation derived from the fatty acid pathway during peach fruit ripening. J. Agric. Food Chem. 2010, 58, 6157. [Google Scholar] [CrossRef]
- Xie, X.; Einhorn, T.; Wang, Y. Inhibition of ethylene biosynthesis and associated gene expression by aminoethoxyvinylglycine and 1-methylcyclopropene and their consequences on eating quality and internal browning of ‘Starkrimson’ pears. J. Am. Soc. Hortic. Sci. 2015, 140, 587–596. [Google Scholar] [CrossRef]
- Axelrod, B.; Cheesbrough, T.M.; Laakso, S.; John, M.L. Lipoxygenase from soybeans: Ec 1.13.11.12 linoleate:oxygen oxidoreductase. Method. Enzym. 1981, 71, 441–451. [Google Scholar]
- Vick, B.A. A spectrophotometric assay for hyderperoxidelyase. Lipids 1991, 26, 315–320. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT | Compound Name | Cas # | Molecular Formula | Korla Volatile Contents (μg/kg) | Abbé Fetel Volatile Contents (μg/kg) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IAT | PRC | IAT | PRC | ||||||||
| CK | MT | CK | MT | CK | MT | CK | MT | ||||
| 4.18 | Acetaldehyde b | 75-07-0 | C2H4O | - | - | - | - | 0.558 ± 0.229 | 0.287 ± 0.109 | 1.045 ± 0.017 | 1.095 ± 0.273 |
| 5.51 | Ethyl Acetate a | 141-78-6 | C4H8O2 | 1.44 ± 0.406 | 5.87 ± 1.895 | 1.45 ± 0.544 | 8.74 * ± 2.756 | 2.37 ± 0.500 | 2.04 ± 1.12 | 6.54 ± 0.370 | 4.955 ± 1.06 |
| 6.69 | n-Propyl acetate a | 109-60-4 | C5H10O2 | - | - | - | - | 0.670 * ± 0.188 | 0.207 ± 0.022 | 1.20 ± 0.281 | 3.81 * ± 1.11 |
| 7.82 | Butanoic acid, ethyl ester a | 105-54-4 | C6H12O2 | 1.51 ± 0.263 | 8.02 * ± 1.320 | 1.52 ± 0.060 | 4.68 ± 2.766 | - | - | - | - |
| 8.50 | Acetic acid, butyl ester a | 123-86-4 | C6H12O2 | 0.417 ± 0.141 | 0.373 ± 0.150 | 0.461 ± 0.074 | 0.340 ± 0.179 | 3.56 ± 0.839 | 2.37 ± 0.338 | 16.58 ± 1.94 | 10.12 ± 3.87 |
| 8.78 | Hexanal a | 66-25-1 | C6H12O | 225.30 * ± 33.18 | 150.84 ± 29.58 | 201.03 ± 75.47 | 230.80 ± 34.22 | 45.57 ± 8.36 | 36.70 ± 8.83 | 102.40 ± 27.58 | 62.91 ± 18.64 |
| 9.63 | 1-Butanol, 2-methyl-, acetate a | 624-41-9 | C7H14O2 | - | - | - | - | 2.24 ± 0.665 | 0.877 ± 0.330 | 10.57 ± 1.31 | 14.61 ± 6.64 |
| 10.11 | 4-Pentenal, 2-methyl- b | 5187-71-3 | C6H10O | 0.151 ± 0.0284 | 0.262 ± 0.197 | 0.161 ± 0.062 | 0.564 ± 0.302 | 0.197 ± 0.0499 | 0.133 ± 0.0158 | - | - |
| 10.90 | Acetic acid, pentyl ester a | 628-63-7 | C7H14O2 | - | - | - | - | 2.26 ± 1.03 | 2.41 ± 0.355 | 17.00 ± 5.06 | 18.499 ± 3.26 |
| 11.80 | 2-Hexyn-1-ol b | 764-60-3 | C6H10O | 0.414 ± 0.030 | 0.773 ± 0.439 | 0.474 ± 0.196 | 1.352 ± 0.459 | 0.251 ± 0.0792 | 0.253 ± 0.0379 | 0.311 ± 0.109 | 0.116 ± 0.0803 |
| 12.39 | 2-Hexenal a | 505-57-7 | C6H10O | 19.51 ± 6.29 | 16.25 ± 1.33 | 12.61 ± 2.37 | 17.60 ± 0.469 | 13.25 ** ± 0.696 | 9.168 ± 0.249 | 23.67 ** ± 8.87 | 7.25 ± 1.10 |
| 12.64 | 1-Butanol, 2-methyl- a | 137-32-6 | C5H12O | - | - | - | - | 0.577 ± 0.0971 | 0.108 ± 0.0380 | 1.49 *** ± 0.0495 | 0.167 ± 0.0170 |
| 13.52 | Acetic acid, hexyl ester a | 142-92-7 | C8H16O2 | 1.87 ± 0.796 | 4.58 ± 1.619 | 6.72 ± 1.017 | 3.10 ± 0.770 | 20.14 ± 5.47 | 14.21 ± 6.58 | 76.22 ± 0.488 | 123.91 ** ± 15.24 |
| 15.36 | 5-Hepten-2-one, 6-methyl- b | 110-93-0 | C8H14O | 0.112 ± 0.005 | 0.336 ± 0.164 | 0.207 ± 0.091 | 0.187 ± 0.024 | 0.238 ± 0.0344 | 0.390 ± 0.220 | - | - |
| 16.50 | 1-Hexanol a | 111-27-3 | C6H14O | 0.043 ± 0.020 | 0.021 ± 0.013 | 0.016 ± 0.008 | 0.025 ± 0.011 | 0.780 ± 0.00552 | 0.678 ± 0.346 | 0.612 ± 0.0795 | 0.417 ± 0.310 |
| 18.93 | Formic acid, heptyl ester b | 112-23-2 | C8H16O2 | - | - | - | - | 0.0180 ± 0.00572 | 0.692 ± 0.0382 | 0.0496 ± 0.0197 | 0.0584 ± 0.00119 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Liu, H.; Wu, T.; Zhai, R.; Yang, C.; Wang, Z.; Ma, F.; Xu, L. Effects of Melatonin Treatment of Postharvest Pear Fruit on Aromatic Volatile Biosynthesis. Molecules 2019, 24, 4233. https://doi.org/10.3390/molecules24234233
Liu J, Liu H, Wu T, Zhai R, Yang C, Wang Z, Ma F, Xu L. Effects of Melatonin Treatment of Postharvest Pear Fruit on Aromatic Volatile Biosynthesis. Molecules. 2019; 24(23):4233. https://doi.org/10.3390/molecules24234233
Chicago/Turabian StyleLiu, Jianlong, Hanting Liu, Ting Wu, Rui Zhai, Chengquan Yang, Zhigang Wang, Fengwang Ma, and Lingfei Xu. 2019. "Effects of Melatonin Treatment of Postharvest Pear Fruit on Aromatic Volatile Biosynthesis" Molecules 24, no. 23: 4233. https://doi.org/10.3390/molecules24234233
APA StyleLiu, J., Liu, H., Wu, T., Zhai, R., Yang, C., Wang, Z., Ma, F., & Xu, L. (2019). Effects of Melatonin Treatment of Postharvest Pear Fruit on Aromatic Volatile Biosynthesis. Molecules, 24(23), 4233. https://doi.org/10.3390/molecules24234233