Sciadopitys verticillata Resin: Volatile Components and Impact on Plant Pathogenic and Foodborne Bacteria

Abstract
1. Introduction
2. Results
2.1. Types of Resin
- Summer: collected in summer and evaluated within 72 h,
- Summer-Stored: collected in the summer and stored for 6 months before evaluation, and
- Winter: collected in winter and evaluated within 72 h.
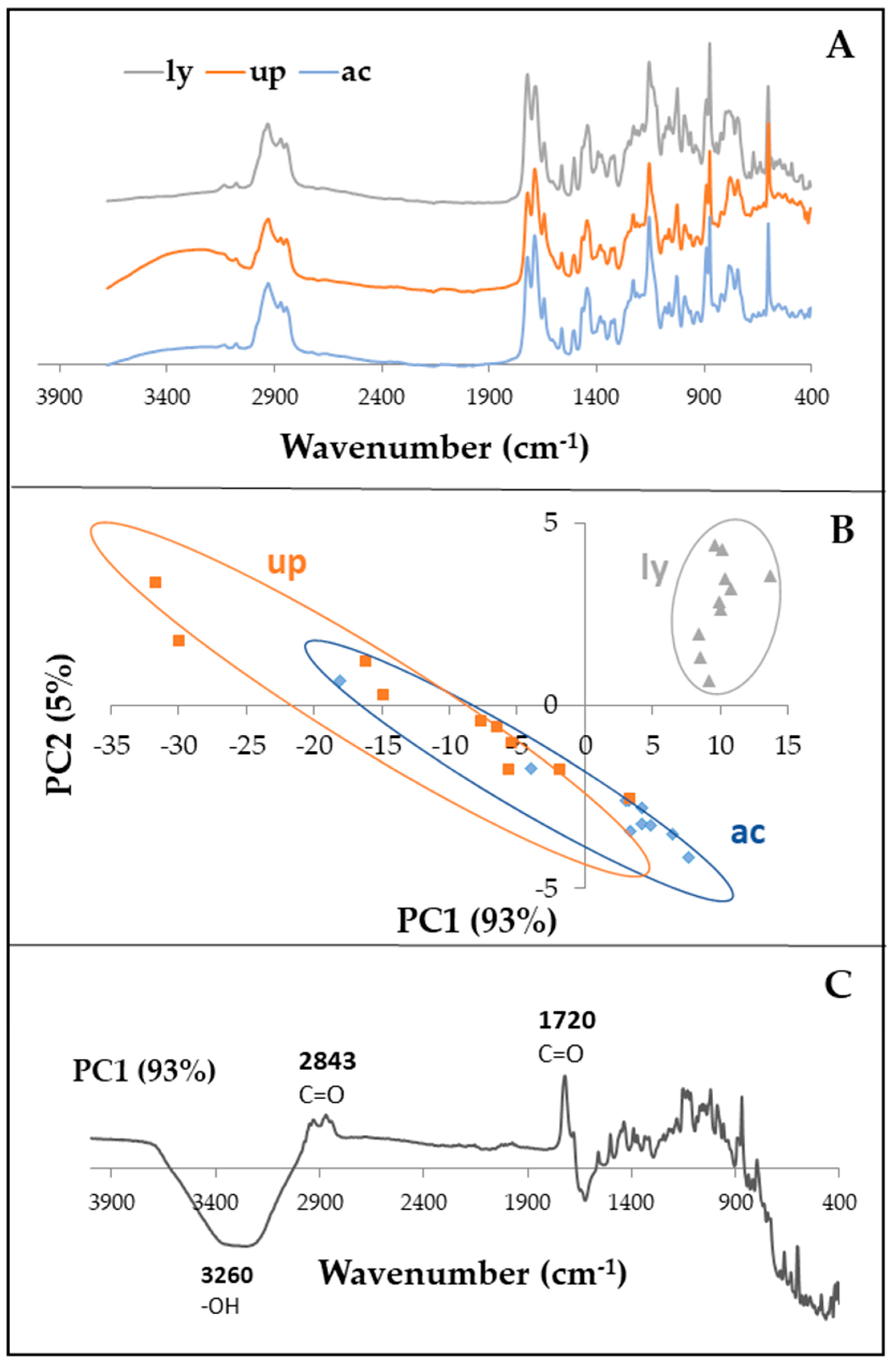
2.2. Effects of Treatment on Resin Chemical Fingerprint
2.3. Gas Chromatography/Mass Spectrometry (GC/MS) Analysis of Resin
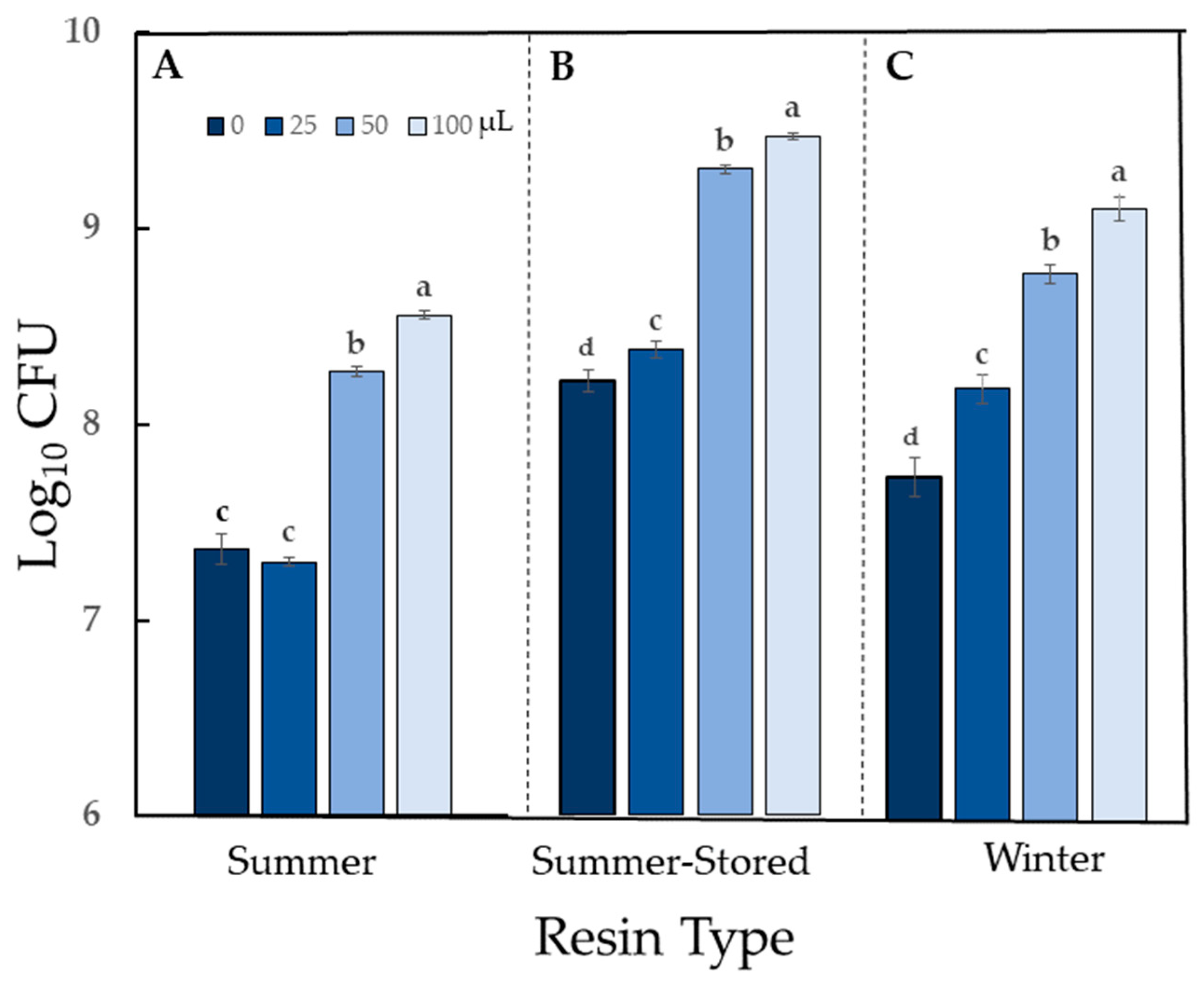
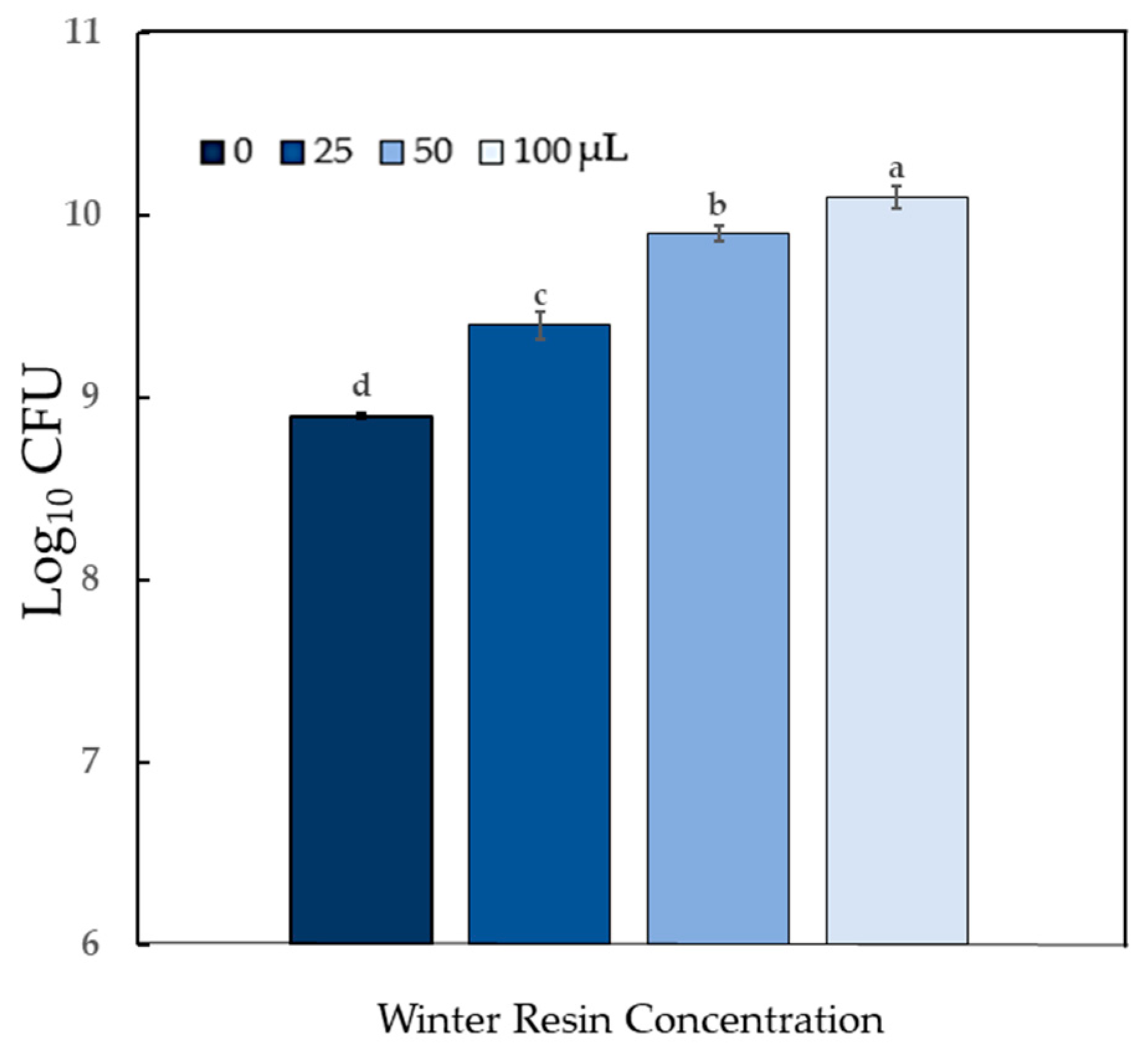
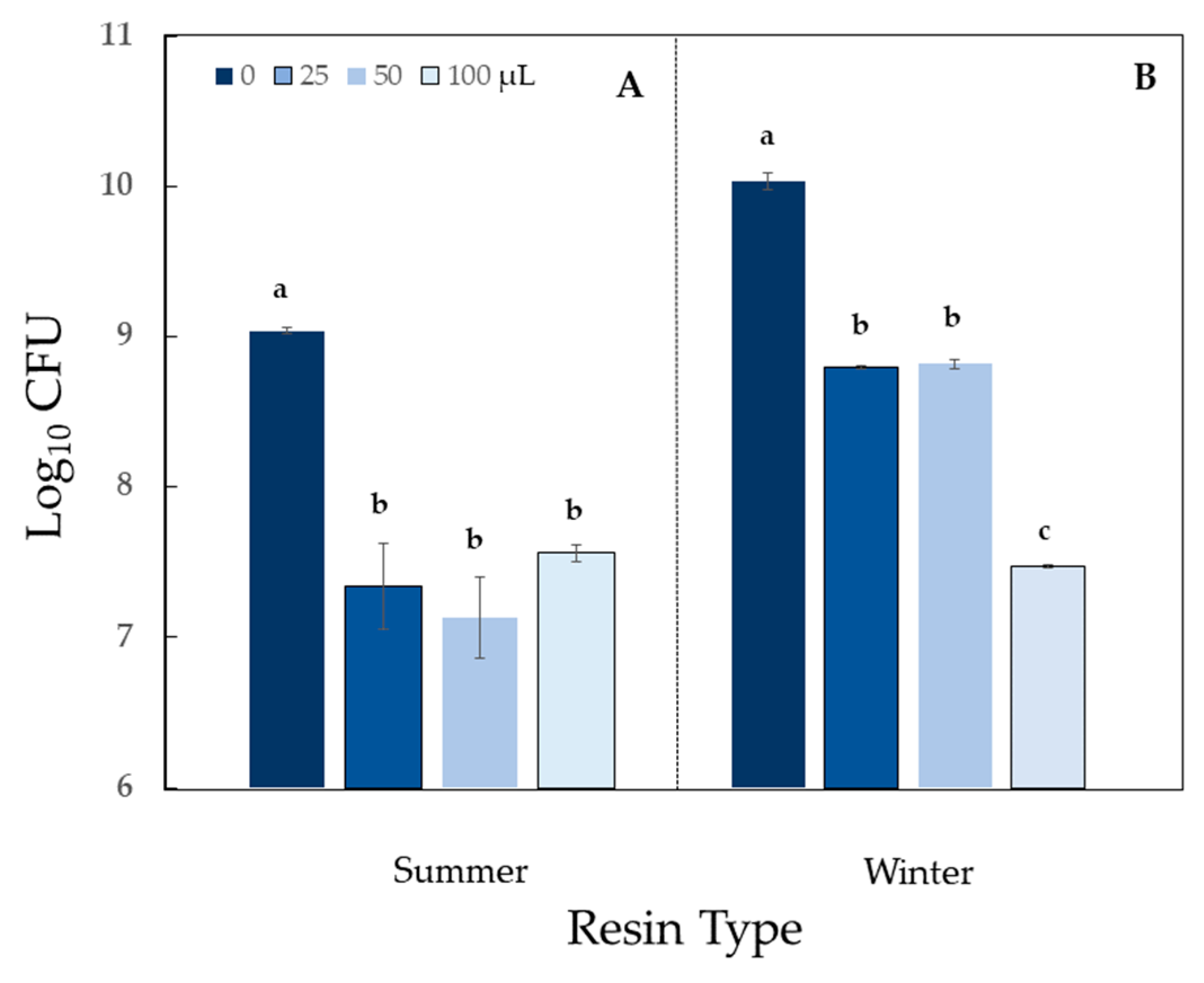
2.4. Impact of Sciadopitys verticillata Resin on Bacterial Population Counts
- Strong antimicrobial/stimulation response: x > 1 log10,
- Mild antimicrobial/stimulation response: x < 0.5 log10,
- Moderate antimicrobial/stimulation response: 0.5 log10 < x < 1 log10.
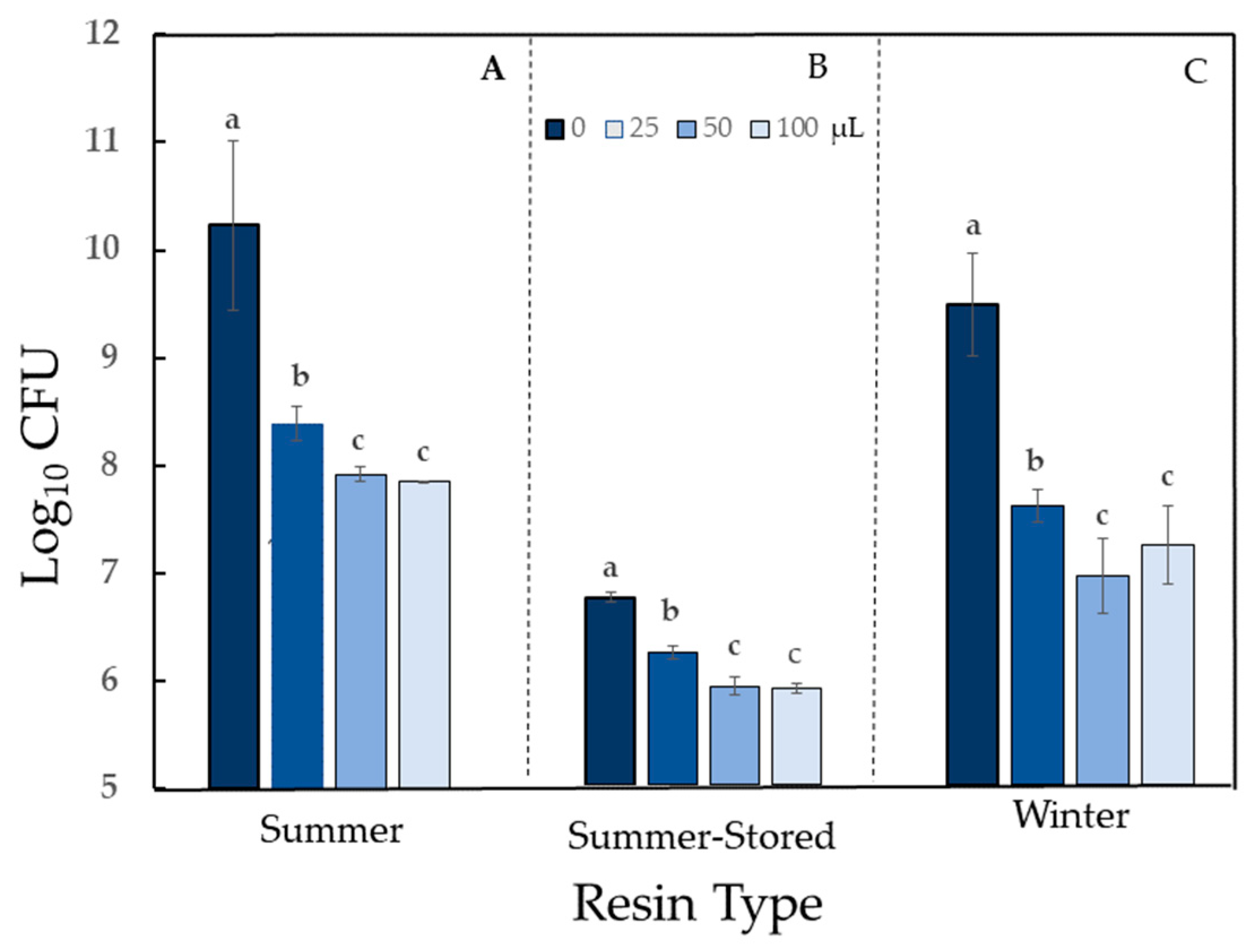
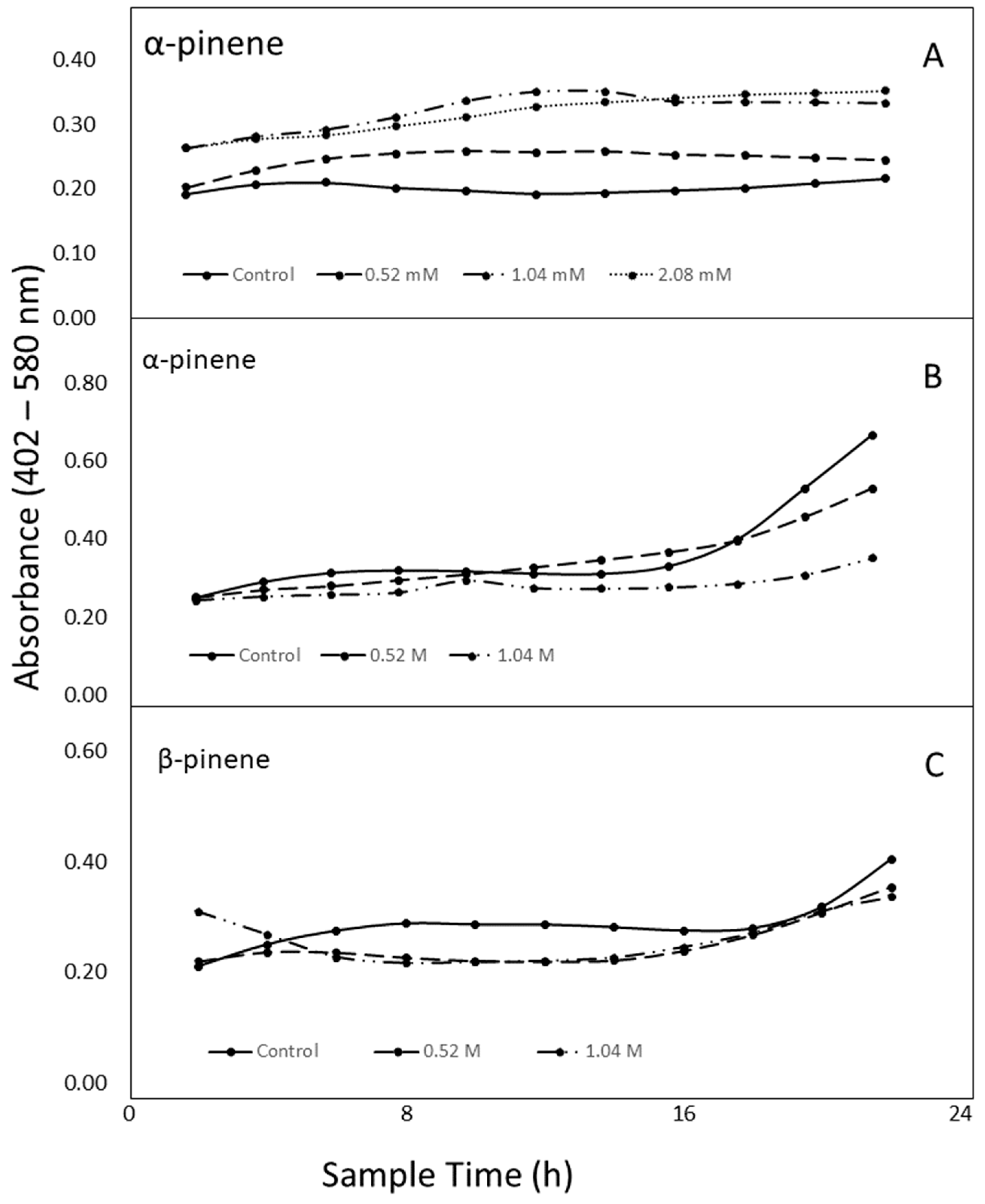
2.5. Antibacterial Activity of α- and β-pinene
3. Discussion
4. Materials and Methods
4.1. Effects of Treatment on Resin Chemical Fingerprint
4.2. GC/MS Analysis of Resin
4.3. Impact of Sciadopitys verticillata Resin on Bacterial Population Counts
4.4. Antibacterial Activity of α- and β-pinene
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Strange, R.N.; Scott, P.R. Plant disease: A threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef] [PubMed]
- Soni, A.; Oey, I.; Silcock, P.; Bremer, P. Bacillus spores in the food industry: A review on resistance and response to novel inactivation technologies. Compr. Rev. Food Sci. F. 2016, 15, 1139–1148. [Google Scholar] [CrossRef]
- Castro, V.S.; Carvalho, R.C.T.; Conte-Junior, C.A.; Souza, E.E. Shiga-toxin producing Escherichia coli: Pathogenicity, supershedding, diagnostic methods, occurrence, and foodborne outbreaks. Compr. Rev. Food Sci. F. 2017, 16, 1269–1280. [Google Scholar] [CrossRef]
- Moloney, M.G. Natural products as a source for novel antibiotics. Trends Pharmacol. Sci. 2016, 37, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Gwinn, K.D. Bioactive natural products in plant disease control. Stud. Nat. Prod. Chem. 2018, 56, 229–246. [Google Scholar] [CrossRef]
- Seiber, J.N.; Coats, J.; Duke, S.O.; Gross, A.D. Pest management with biopesticides. Front. Agri. Sci. Eng. 2018, 5, 295–300. [Google Scholar] [CrossRef]
- Worth, J.R.; Sakaguchi, S.; Tanaka, N.; Yamaski, M.; Isagi, Y. Northern richness and southern poverty: Contrasting genetic footprints of glacial refugia in the relictual tree Sciadopitys verticillata. Biol. J. Linn. Soc. 2013, 108, 263–277. [Google Scholar] [CrossRef]
- Sadowski, E.M.; Schmidt, A.R.; Kunzman, L.; Grohn, C.; Seyfullah, L.J. Sciadopitys cladodes from Eocene Baltic amber. Biol. J. Linn. Soc. 2016, 180, 258–268. [Google Scholar] [CrossRef]
- Yates, D.I.; Earp, B.L.; Walker, E.S.; Levy, F. Propagation of Sciadopitys verticillata (Thunb.) Sieb. & Zucc. by stem cuttings and properties of its latex-like sap. HortScience 2006, 41, 1662–1666. [Google Scholar]
- Trapp, S.; Croteau, R. Defensive resin biosynthesis in conifers. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 2001, 52, 689–724. [Google Scholar] [CrossRef]
- Wolfe, A.P.; Tappert, R.; Muehlenbachs, K.; Boudreau, M.; McKellar, R.C.; Basinger, J.F. A new proposal concerning the botanical origin of Baltic amber. Proc. R. Soc. Lond. 2009, 276, 3403–3412. [Google Scholar] [CrossRef] [PubMed]
- Tappert, R.; Wolfe, A.P.; McKellar, R.C.; Tappert, M.C.; Muehlenbachs, K. Characterizing modern and fossil gymnosperm exudates using micro-Fourier infrared spectroscopy. Int. J. Plant Sci. 2011, 172, 120–138. [Google Scholar] [CrossRef]
- Yates, D.I. Antibacterial activity and chemical characterization of resin from Sciadopitys verticillata (Thunb.) Siebold and Zuccarini. Ph.D. Dissertation, University of Tennessee, Knoxville, TN, USA, 2016. [Google Scholar]
- Sutton, S.V.; Porter, D. Development of the antimicrobial effectiveness test as USP <51>. PDA J. Pharm. Sci. Tech. 2002, 56, 300–311. [Google Scholar]
- Periche, A.; Castelló, M.L.; Heredia, A.; Escriche, I. Effect of different drying methods on the phenolic, flavonoid and volatile compounds of Stevia rebaudiana leaves. Flavour. Fragr. J. 2016, 31, 173–177. [Google Scholar] [CrossRef]
- Langenheim, J.H. Plant Resins: Chemistry, Evolution, Ecology, and Ethnobotany, 1st ed.; Timber Press: Portland, OR, USA, 2003. [Google Scholar]
- Meccia, J.; Rojas, L.B.; Velasco, J.; Díaz, T.; Usubillaga, A.; Arzola, J.C.; Ramos, S. Chemical composition and antibacterial activity of the essential oil of Cordia verbenaceae from the Venezuelan Andes. Nat. Prod. Commun. 2009, 8, 1119–1122. [Google Scholar] [CrossRef]
- Montenegro, I.; Madrid, A.; Cuellar, M.; Seeger, M.; Alfaro, J.F.; Besoain, X.; Martinez, J.P.; Ramirez, I.; Olguin, Y.; Valenzuela, M. Biopesticide activity from drimanic compounds to control tomato pathogens. Molecules 2018, 23, 2053. [Google Scholar] [CrossRef]
- Violanova, C.; Marin, M.; Baixeras, J.; Latorre, A.; Porcar, M. Selecting microbial strains from pine tree resin: Biotechnological applications from a terpene world. PLoS ONE 2014, 9, e100740. [Google Scholar] [CrossRef]
- Zengin, H.; Baysal, A.H. Antibacterial and antioxidant activity of essential oil terpenes against pathogenic and spoilage-forming bacteria and cell structure-activity relationships evaluated by SEM microscopy. Molecules 2014, 19, 17773–17798. [Google Scholar] [CrossRef]
- da Silva, A.C.R.; Lopes, P.M.; de Azevedo, M.M.B.; Costa, D.C.M.; Alviano, C.S.; Alviano, D.S. Biological activities of α-pinene and β-pinene enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef]
- Cheng, Z.W.; Chen, J.M.; Yu, J.M.; Li, S.S. Structural characterization and property analysis of surface-active substance accumulated during biodegradation of hydrophobic α-pinene by Pseudomonas fluorescens. Water Air Soil Poll. 2013, 224, 1457–1461. [Google Scholar] [CrossRef]
- Wang, X.; Yu, Y.; Ge, J.; Xie, B.; Zhu, S.; Cheng, X. Effects of α-pinene on the pinewood nematode (Bursaphelenchus xylophilus) and its symbiotic bacteria. PLOS ONE 2019, 14, e0221099. [Google Scholar] [CrossRef] [PubMed]
- Scortichini, M.; Rossi, M.P. Preliminary in vitro evaluation of the antimicrobial activity of terpenes and terpenoids towards Erwinia amylovora (Burrill). J. Appl. Bacteriol. 1991, 71, 108–112. [Google Scholar] [CrossRef]
- Dhar, P.; Chan, P.Y.; Cohen, D.T.; Khawam, F.; Gibbons, S.; Snyder-Leiby, T.; Dickstein, E.; Rai, P.K.; Watal, G. Synthesis, antimicrobial evaluation, and structure activity relationship of α-pinene derivatives. J. Agric. Food Chem. 2014, 62, 3548–3552. [Google Scholar] [CrossRef] [PubMed]
- Kennelly, M.M.; Cazoria, F.M.; Vincente, A.D.; Ramos, C.; Sundin, G.W. Pseudomonas syringae diseases of fruit trees, progress toward understanding and control. Plant Dis. 2007, 91, 4–17. [Google Scholar] [CrossRef]
- Maughan, H.; Van der Auwera, G. Bacillus taxonomy in the genomic era finds phenotypes to be essential though often misleading. Infect. Genet. Evol. 2011, 11, 789–797. [Google Scholar] [CrossRef]
- Liu, Y.; Lai, Q.; Göker, M.; Meier-Kolthoff, J.P.; Wang, M.; Sun, Y.; Wang, L.; Shao, Z. Genomic insights into the taxonomic status of the Bacillus cereus group. Sci. Rep. 2015, 5, 14082. [Google Scholar] [CrossRef]
- Amor, M.G.-B.; Jan, S.; Baron, F.; Grosset, N.; Culot, A.; Gdoura, R.; Gautier, M.; Techer, C. Toxigenic potential and antimicrobial susceptibility of Bacillus cereus group bacteria isolated from Tunisian foodstuffs. BMC Microbiol. 2019, 19, 196. [Google Scholar] [CrossRef]
- Saleh-Lakha, S.; Leon-Velarde, C.G.; Chen, S.; Lee, S.; Shannon, K.; Fabri, M.; Downing, G.; Keown, B. A study to assess the numbers and prevalence of Bacillus cereus and its toxins in pasteurized fluid milk. J. Food Protect. 2017, 80, 1085–1089. [Google Scholar] [CrossRef]
- Savithiry, N.; Gage, D.; Fu, W.; Orielet, P. Degradation of pinene by Bacillus pallidus BR425. Biodegradation 1998, 9, 337–341. [Google Scholar] [CrossRef]
- Miῆana-Galbis, D.; Pinzón, D.L.; Gaspar Lorén, J.; Manresa, A.; Oliart-Ros, R.M. Reclassification of Geobacillus pallidus (Scholz et al. 1988) Banat et al. 2004 as Aeribacillus pallidus gen. nov., comb. nov. Int. J. Syst. Evol. 2010, 60, 1600–1604. [Google Scholar] [CrossRef]
- Adams, A.S.; Boone, C.K.; Bohlmann, J. Responses of bark beetle-associated bacteria to host monoterpenes and their relationship to insect life histories. J. Chem. Ecol. 2011, 37, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Dee, M.M.; Quigley, N.B.; Ownley, B.H. Micro-dilution plating technique for assessing population counts of microorganisms. Phytopathology 1995, 85, 1204. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SUMMER-STORED | WINTER | |||||
|---|---|---|---|---|---|---|
| Compound a | Retention Time (min) | Vapor Pressure b | Score | % Peak Area | Score | % Peak Area |
| Monoterpenes | 94.738 | 92.232 | ||||
| 1R-α-Pinene c | 5.474 | 4.75 | 93 | 70.677 | 92 | 82.003 |
| Tricyclene | 5.399 | 4.45 | 88 | 16.313 | NP | NP |
| β-Pinene c | 5.794 | 2.93 | 81 | 5.394 | 83 | 7.656 |
| D-Limonene | 6.132 | 1.98 | 88 | 1.570 | 88 | 1.784 |
| Camphene | 5.600 | 2.5 | 81 | 0.784 | 80 | 0.789 |
| Methane monoterpenes | 0.855 | 0.703 | ||||
| 3 7 α-Terpinyl propionate | 8.306 | 0.013 | 78 | 0.415 | 80 | 0.579 |
| 7 α-Terpinyl propionate | 9.588 | 0.013 | 63 | 0.040 | 69 | 0.124 |
| Sesquiterpenes | 3.193 | 4.925 | ||||
| β-Elemene | 9.222 | 0.0276 | 94 | 2.441 | 94 | 3.635 |
| β-Cubebene | 8.907 | 0.014 | 82 | 0.373 | 85 | 0.541 |
| γ-Cadinene | 8.627 | 0.003 | 85 | 0.164 | 88 | 0.267 |
| Caryophyllene | 8.867 | 0.300 | 87 | 0.109 | 88 | 0.195 |
| Copaene | 8.558 | 0.038 | 82 | 0.106 | 79 | 0.159 |
| γ-Cadinene | 9.067 | 0.003 | NP | 0.000 | 93. | 0.128 |
| Other | 1.248 | 2.139 | ||||
| Contaminant (Siloxane) | 10.189 | NA | NA | 0.796 | NA | 0.900 |
| Unknown d | 8.844 | NA | 85 | 0.369 | 87 | 0.586 |
| 1-Naphthalenol | 9.405 | NA | 77 | 0.304 | 78 | 0.472 |
| β-Ionone | 9.954 | 0.054 | 72 | 0.104 | 74 | 0.181 |
| 1,5,9,9-Tetramethyl-1,4,7-cycloundecatriene | 9.073 | NA | 70 | 0.043 | NP | 0.000 |
| 11-Acetoxy tetracyclododecane | 9.845 | 0.135 | 61 | 0.032 | NP | 0.000 |
| Family | Pathogen | Economic Importance | Reaction to Resin 2 | ||
|---|---|---|---|---|---|
| (Gram stain) | Strain 1 | Summer | Winter | Summer-Stored | |
| Bacilliaceae (+) | Bacillus cereus CB# 154869 | foodborne illness | --- | -- | n.d. |
| Enterobacteriaceae (-) | Erwinia amylovora UTBO# E9 | plant pathogen | --- | --- | -- |
| Escherichia coli CB# 155068 | foodborne illness; biotechnology | - | - | - | |
| Pseudomonadaceae (-) | Pseudomonas fluorescens CB# 155255 | food contaminant; bio-manufacturing; biological control | +++ | ++ | +++ |
| Pseudomonas syringae pv. tomato UTBO# 268 | plant pathogen | n.d. | +++ | n.d. | |
| Rhizobiaceae (-) | Agrobacterium tumefaciens UTBO# C58 | plant pathogen | - | - | n.d. |
| Xanthomonadaceae (-) | Xanthomonas perforans UTBO# SB1 | plant pathogen; bio-manufacturing | + | + | n.d. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yates, D.I.; Ownley, B.H.; Labbé, N.; Bozell, J.J.; Klingeman, W.E.; Batson, E.K.; Gwinn, K.D. Sciadopitys verticillata Resin: Volatile Components and Impact on Plant Pathogenic and Foodborne Bacteria. Molecules 2019, 24, 3767. https://doi.org/10.3390/molecules24203767
Yates DI, Ownley BH, Labbé N, Bozell JJ, Klingeman WE, Batson EK, Gwinn KD. Sciadopitys verticillata Resin: Volatile Components and Impact on Plant Pathogenic and Foodborne Bacteria. Molecules. 2019; 24(20):3767. https://doi.org/10.3390/molecules24203767
Chicago/Turabian StyleYates, David I., Bonnie H. Ownley, Nicole Labbé, Joseph J. Bozell, William E. Klingeman, Emma K. Batson, and Kimberly D. Gwinn. 2019. "Sciadopitys verticillata Resin: Volatile Components and Impact on Plant Pathogenic and Foodborne Bacteria" Molecules 24, no. 20: 3767. https://doi.org/10.3390/molecules24203767
APA StyleYates, D. I., Ownley, B. H., Labbé, N., Bozell, J. J., Klingeman, W. E., Batson, E. K., & Gwinn, K. D. (2019). Sciadopitys verticillata Resin: Volatile Components and Impact on Plant Pathogenic and Foodborne Bacteria. Molecules, 24(20), 3767. https://doi.org/10.3390/molecules24203767