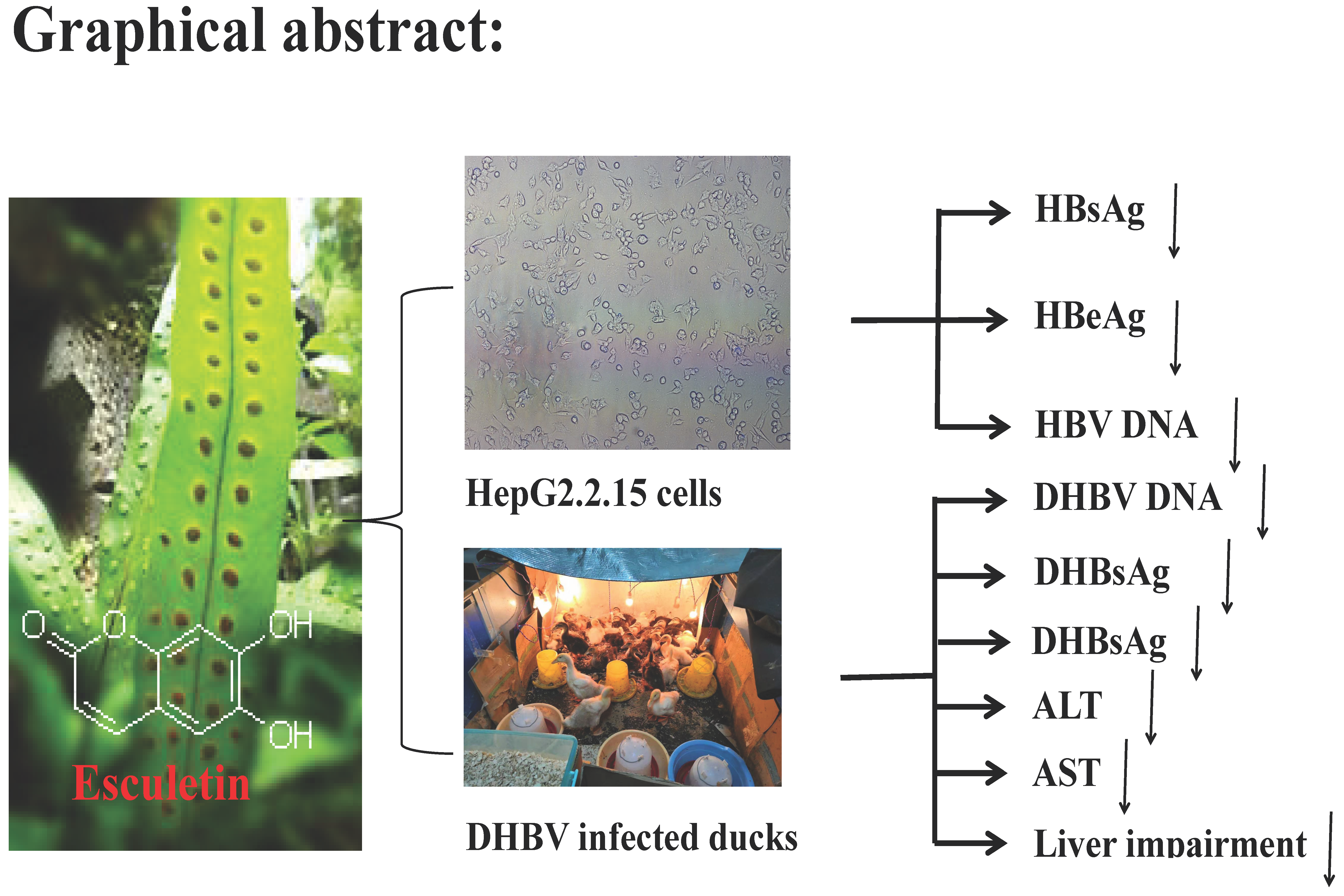
Anti-Hepatitis B Virus Activity of Esculetin from Microsorium fortunei In Vitro and In Vivo

Abstract
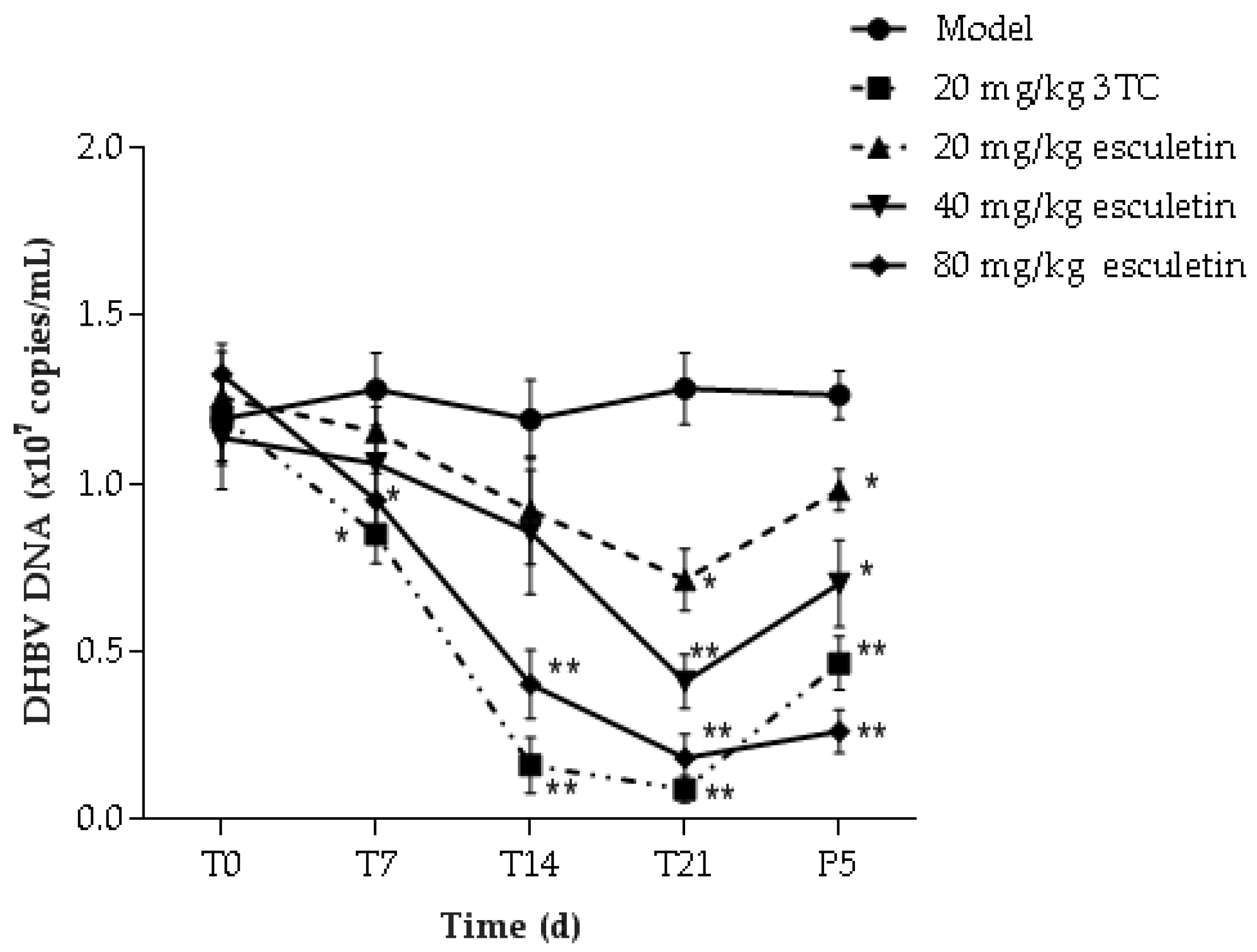
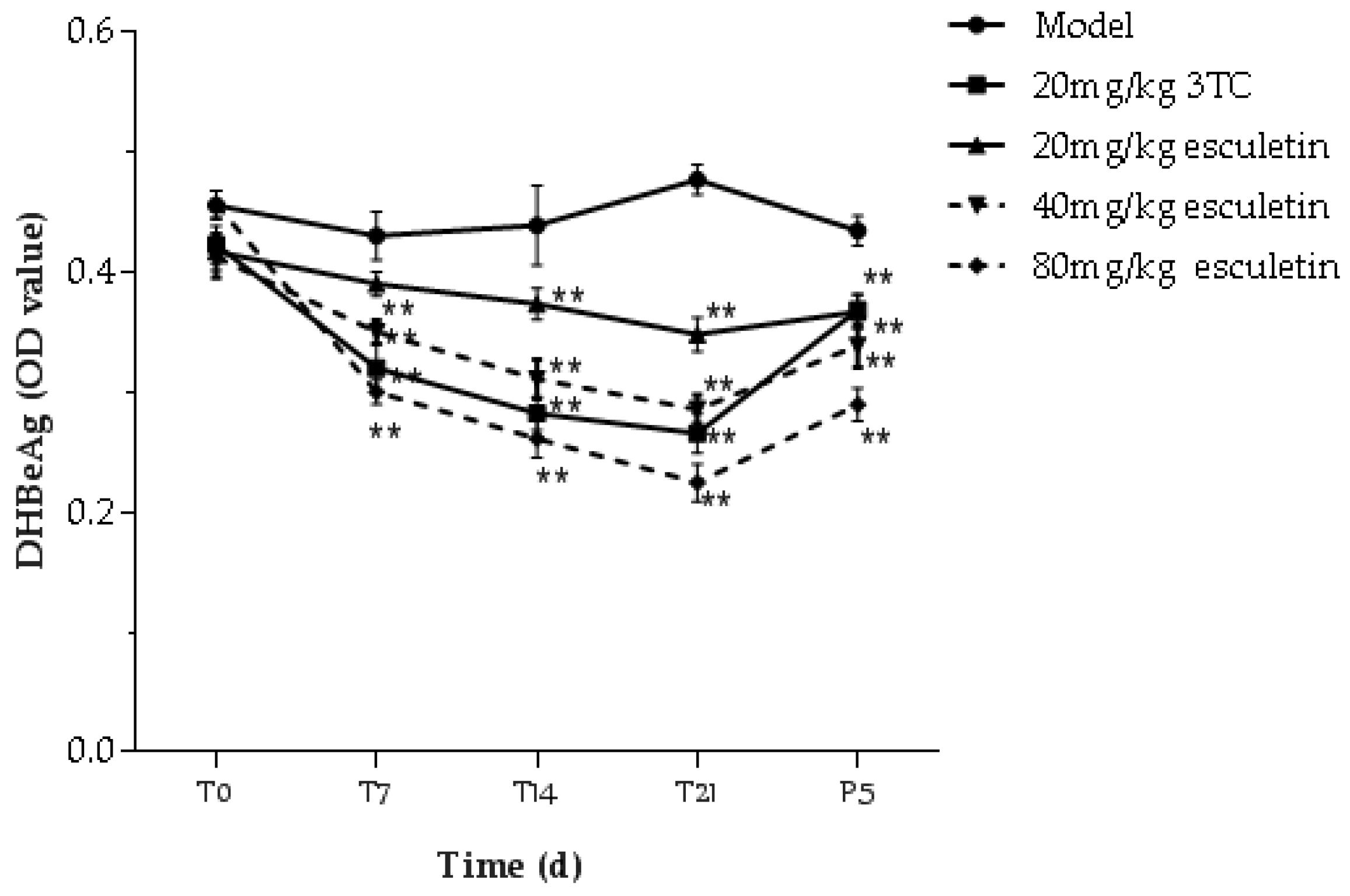
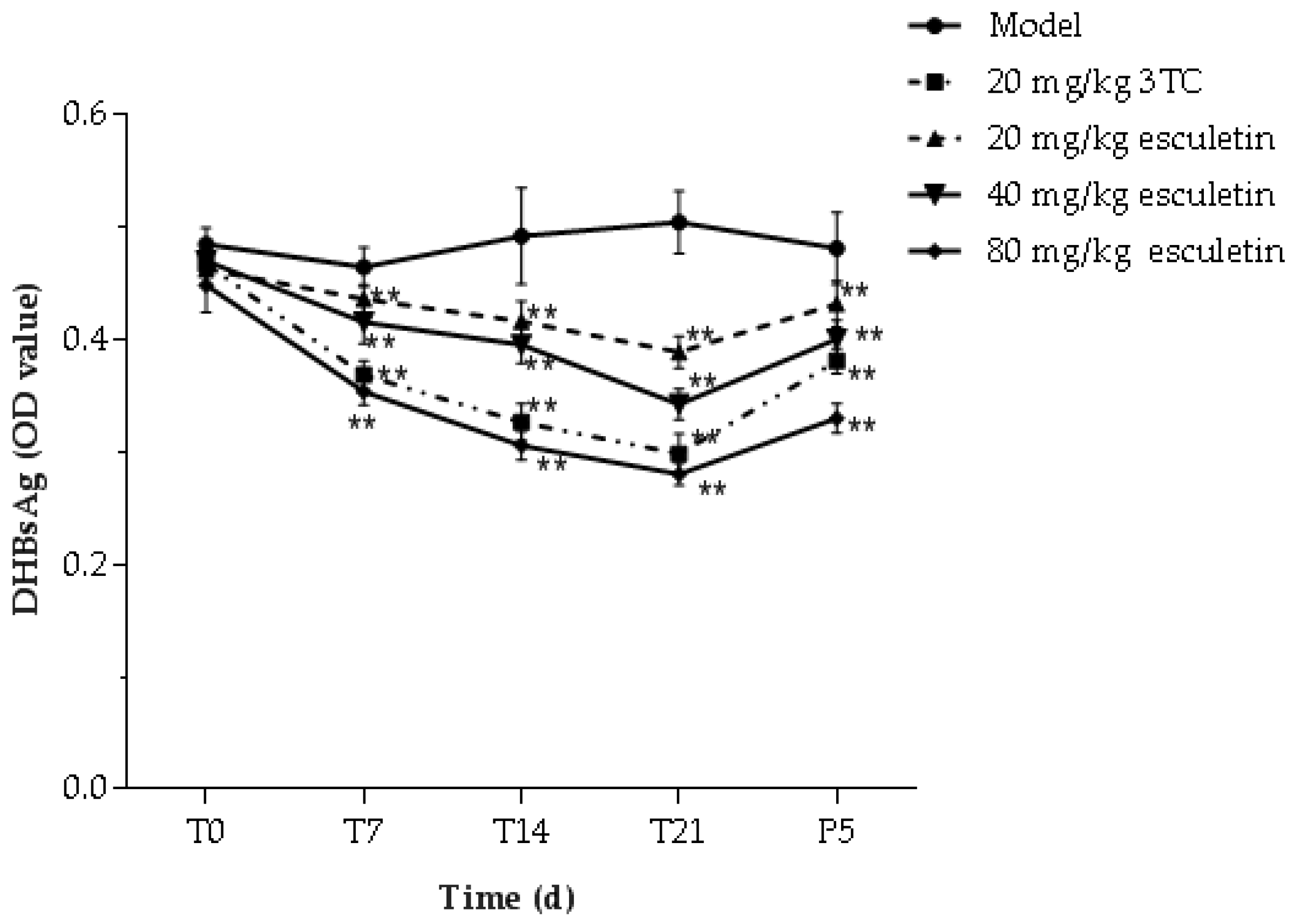
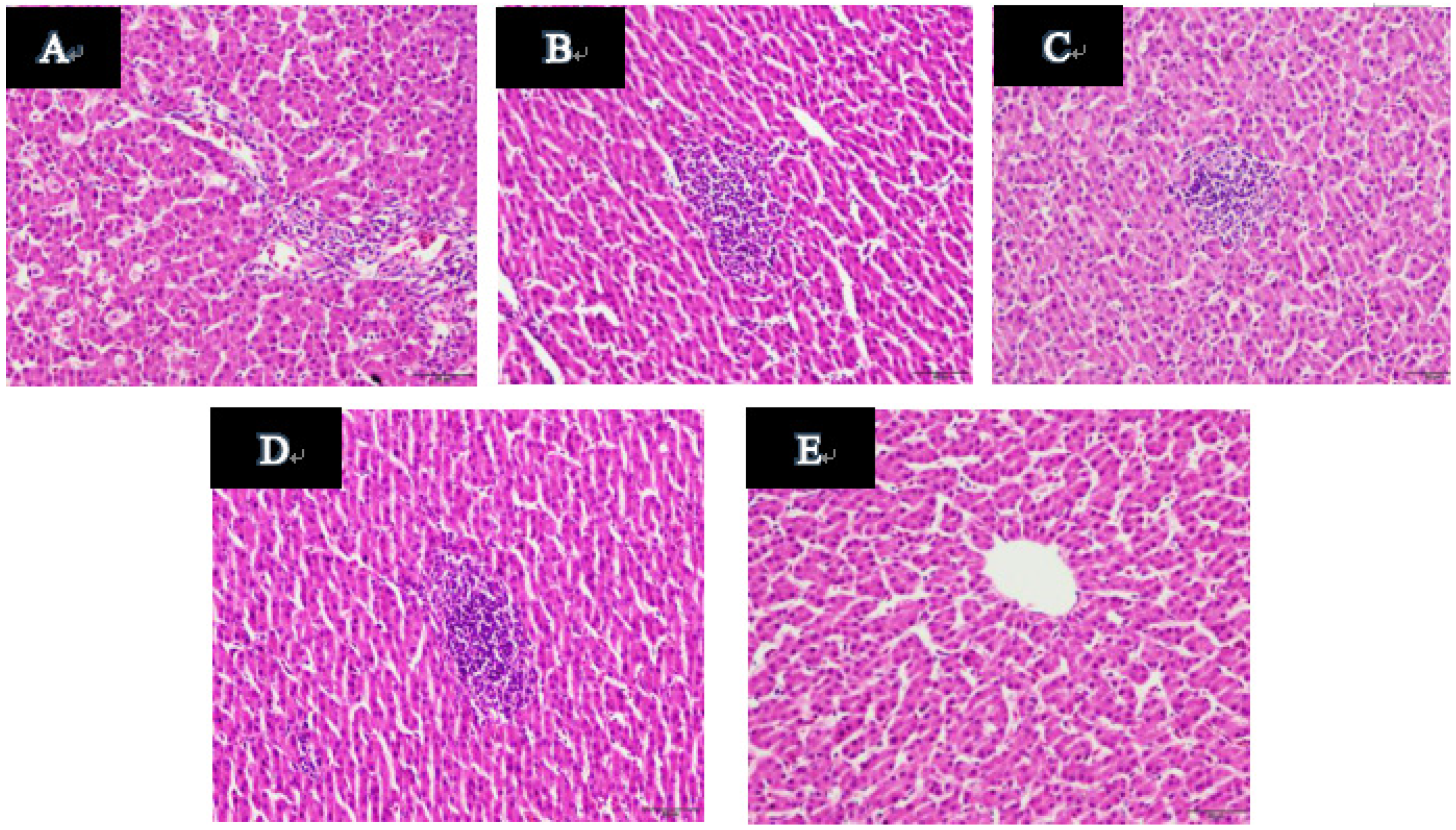
:
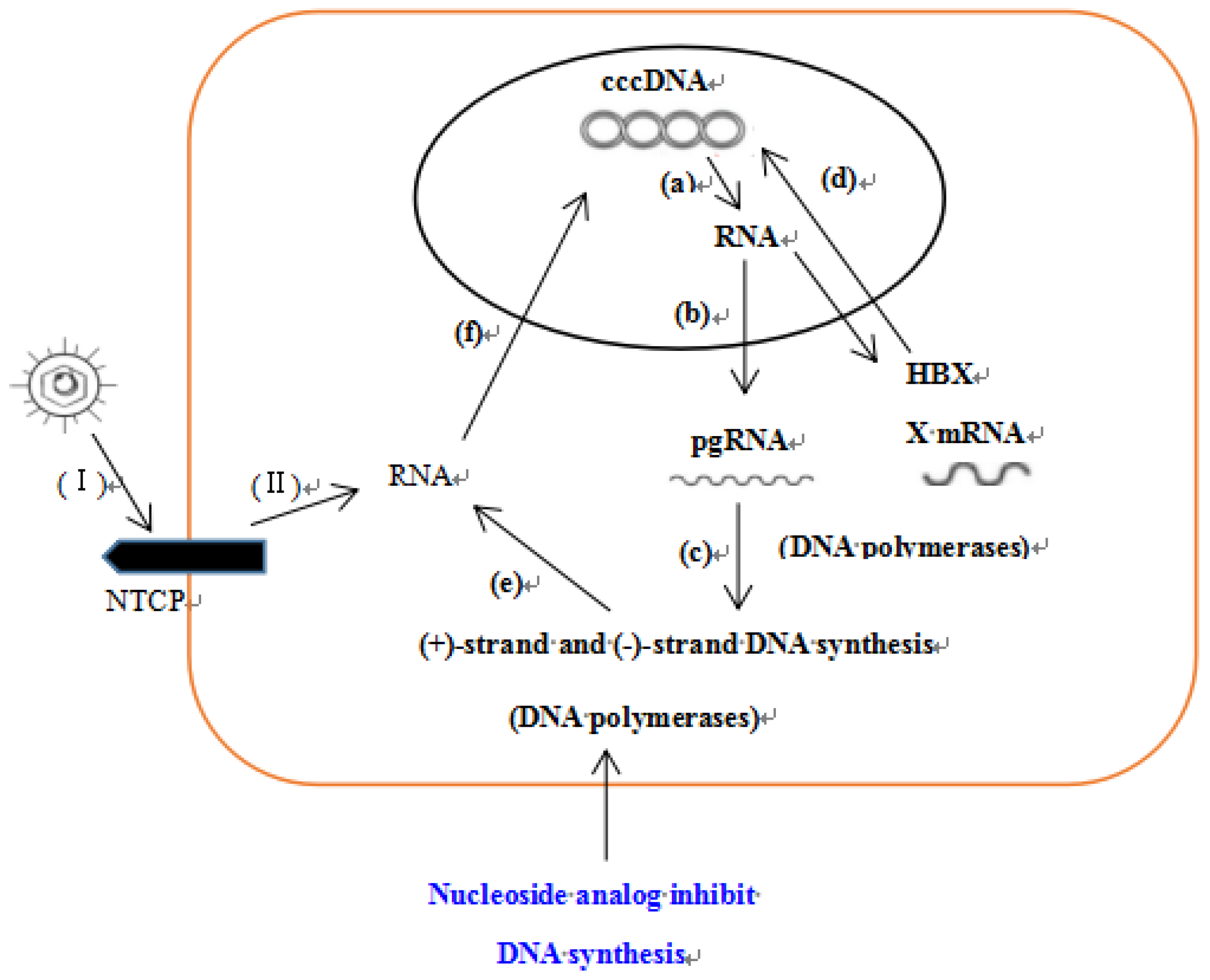
1. Introduction
2. Results
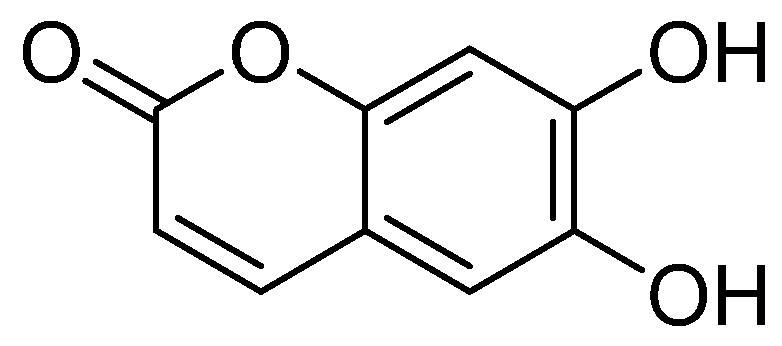
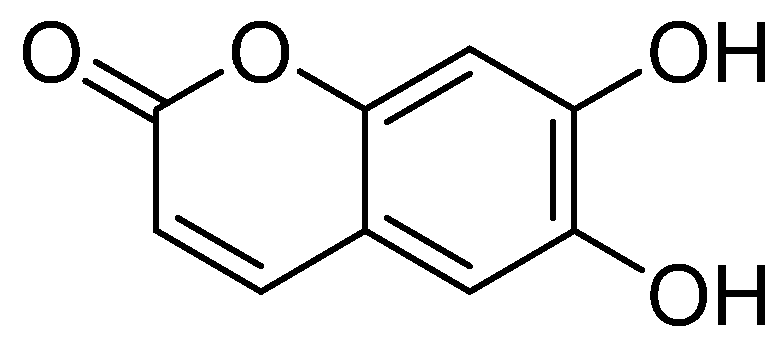
2.1. Structural Identification
2.2. Cytotoxic Effect of Esculetin for HL-7702 Cells
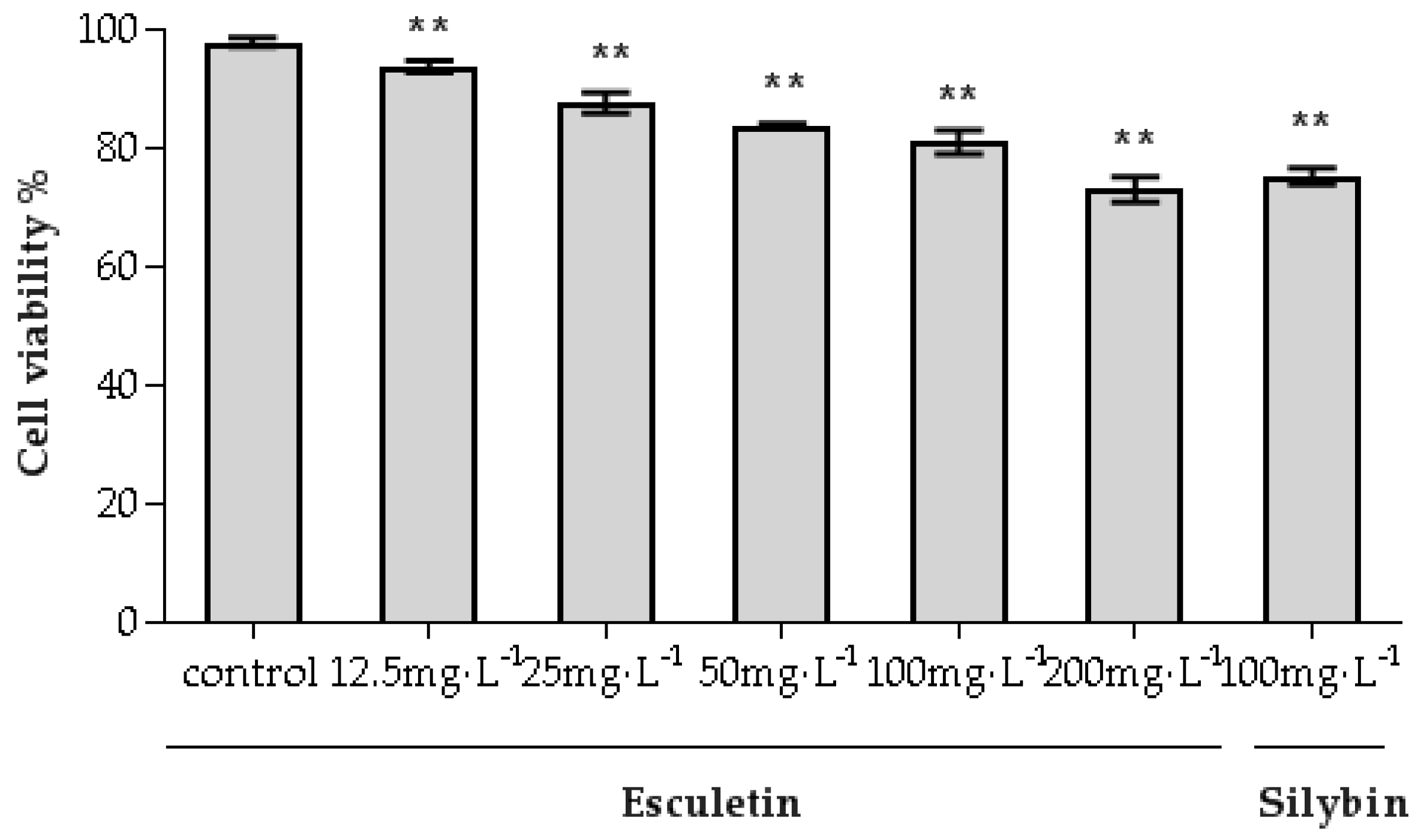
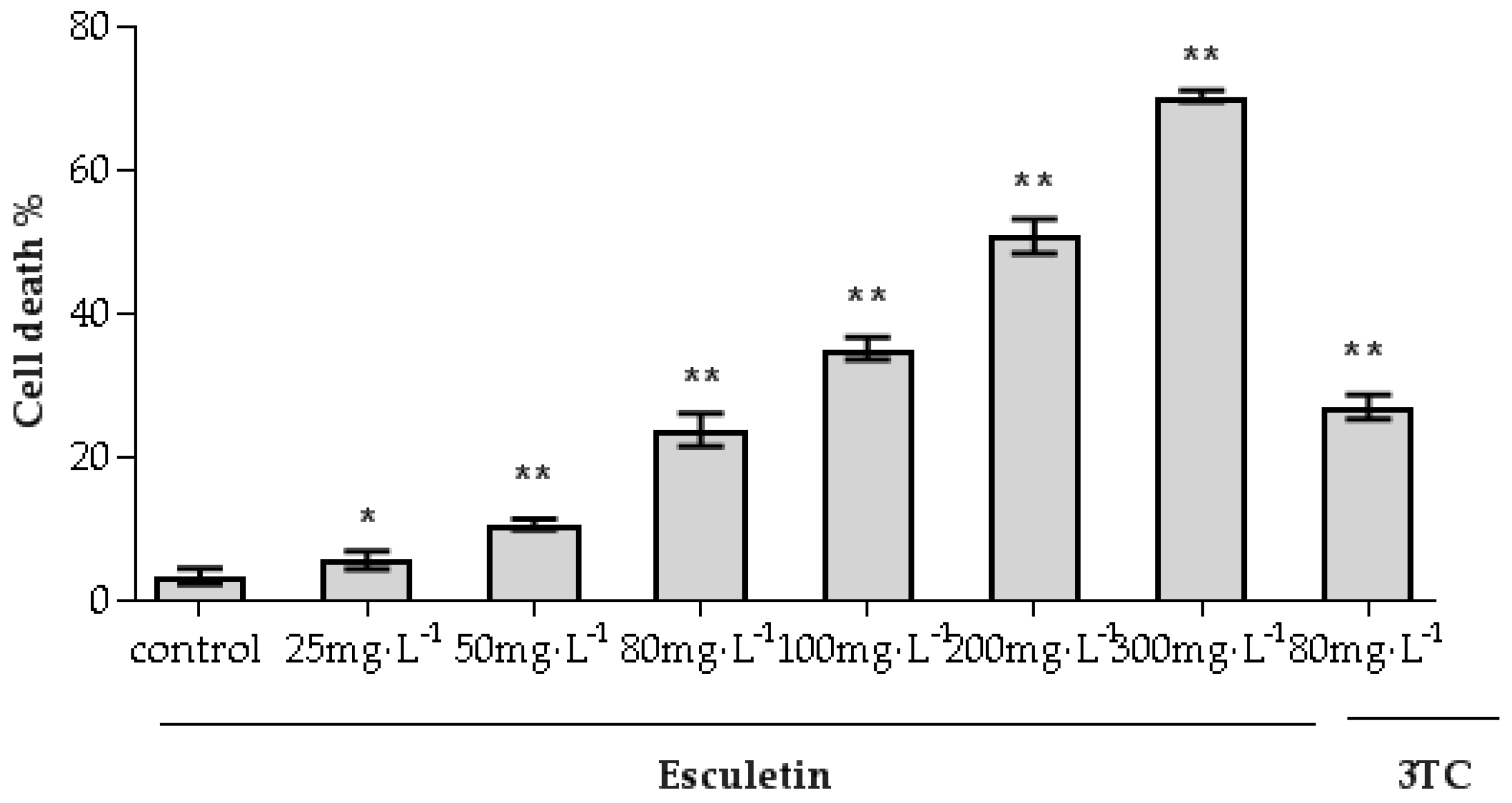
2.3. Cytotoxic Effect of Esculetin for HepG2.2.15 Cells
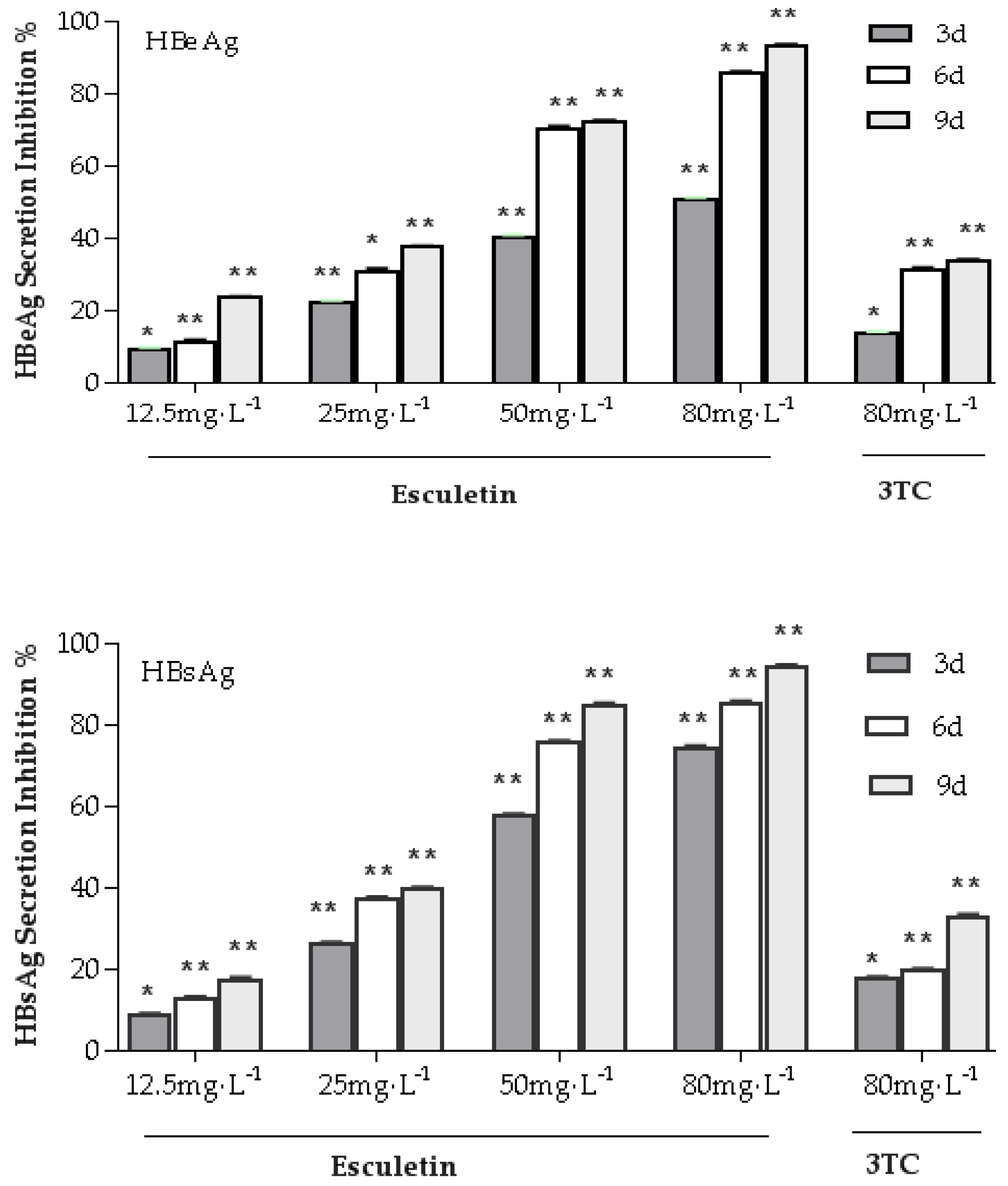
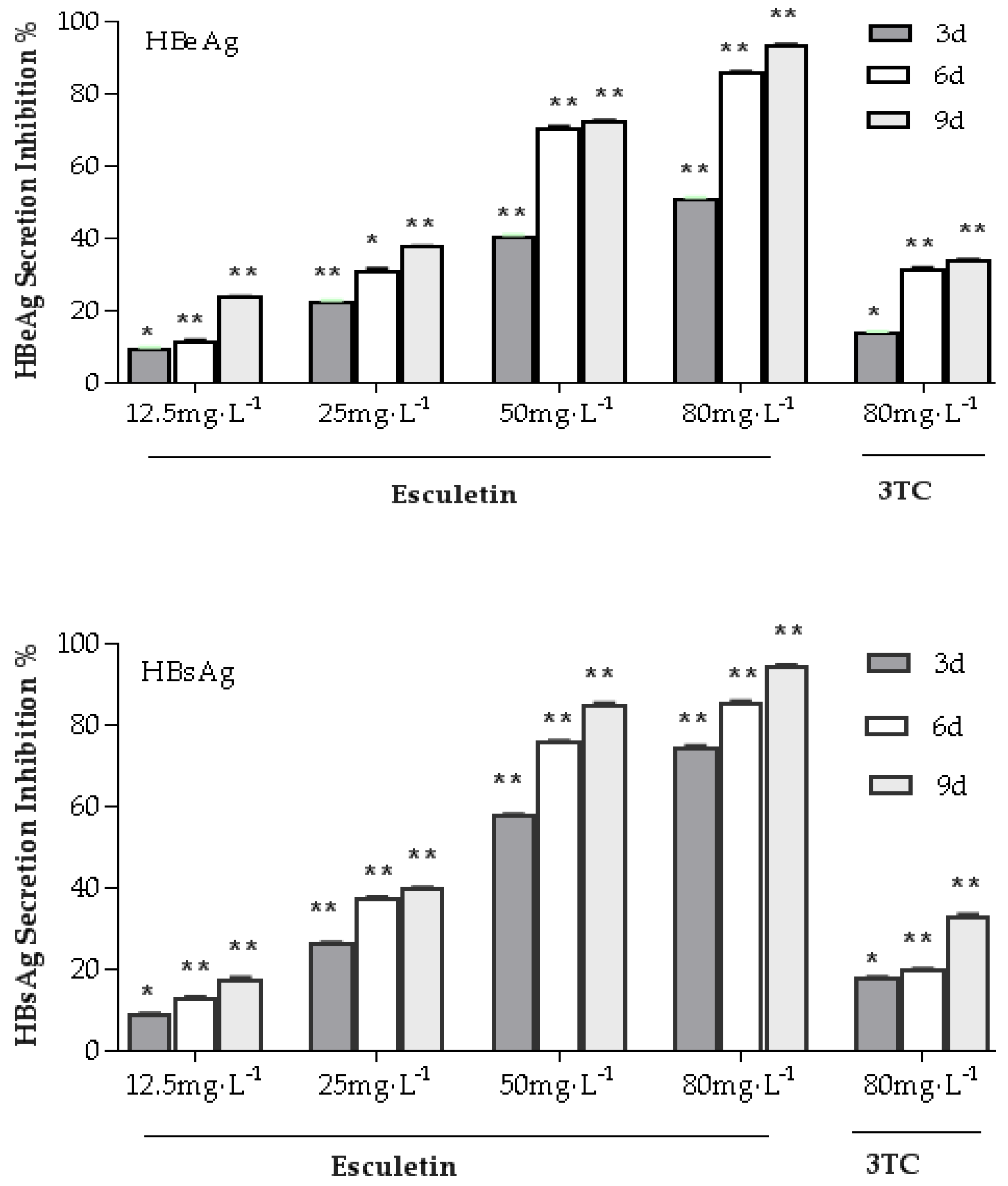
2.4. Effects of Esculetin on the Secretion of the HBsAg and HBeAg in the HepG2.2.15 Cell Line
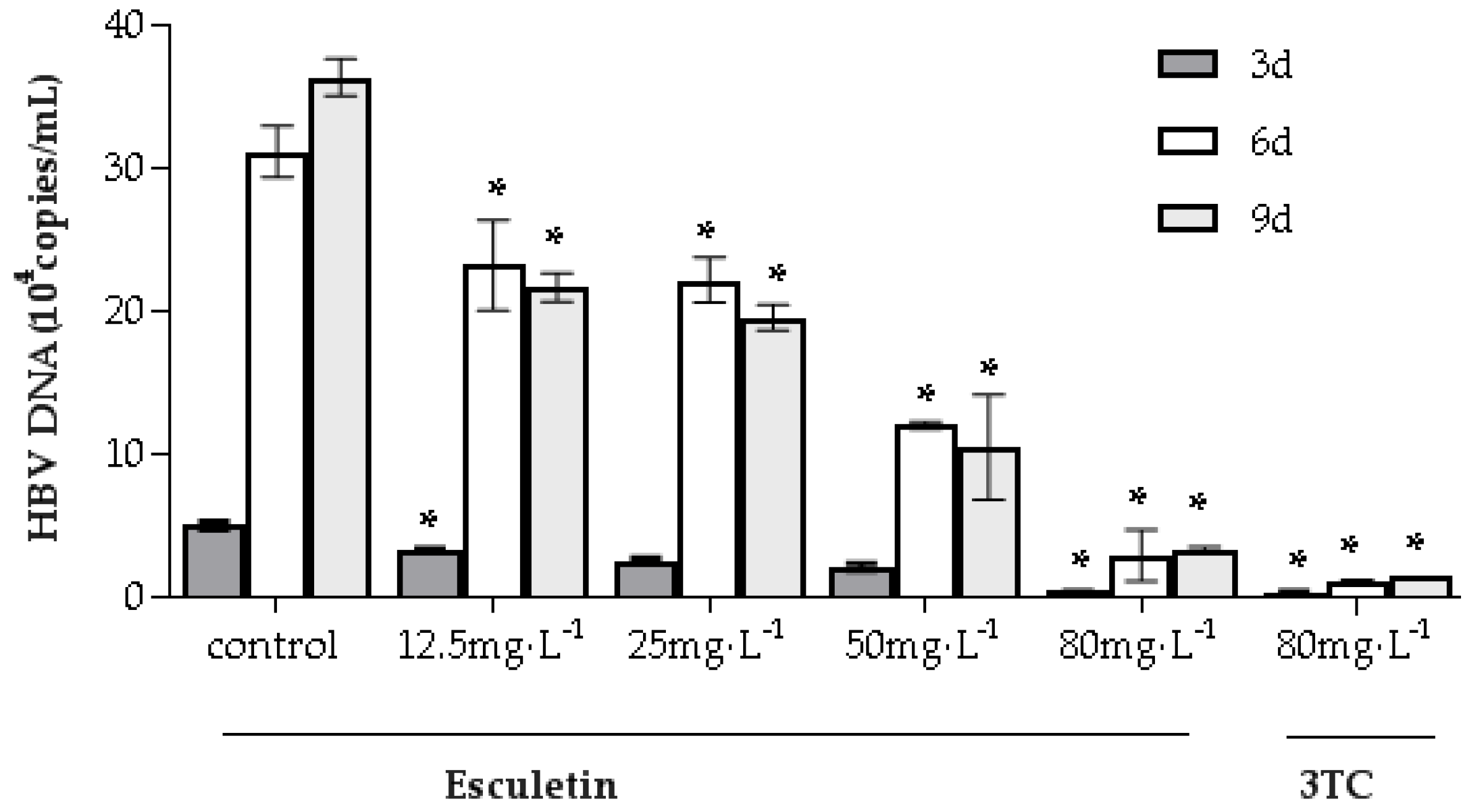
2.5. Effects of Esculetin on Secretion of Supernatant HBV DNA
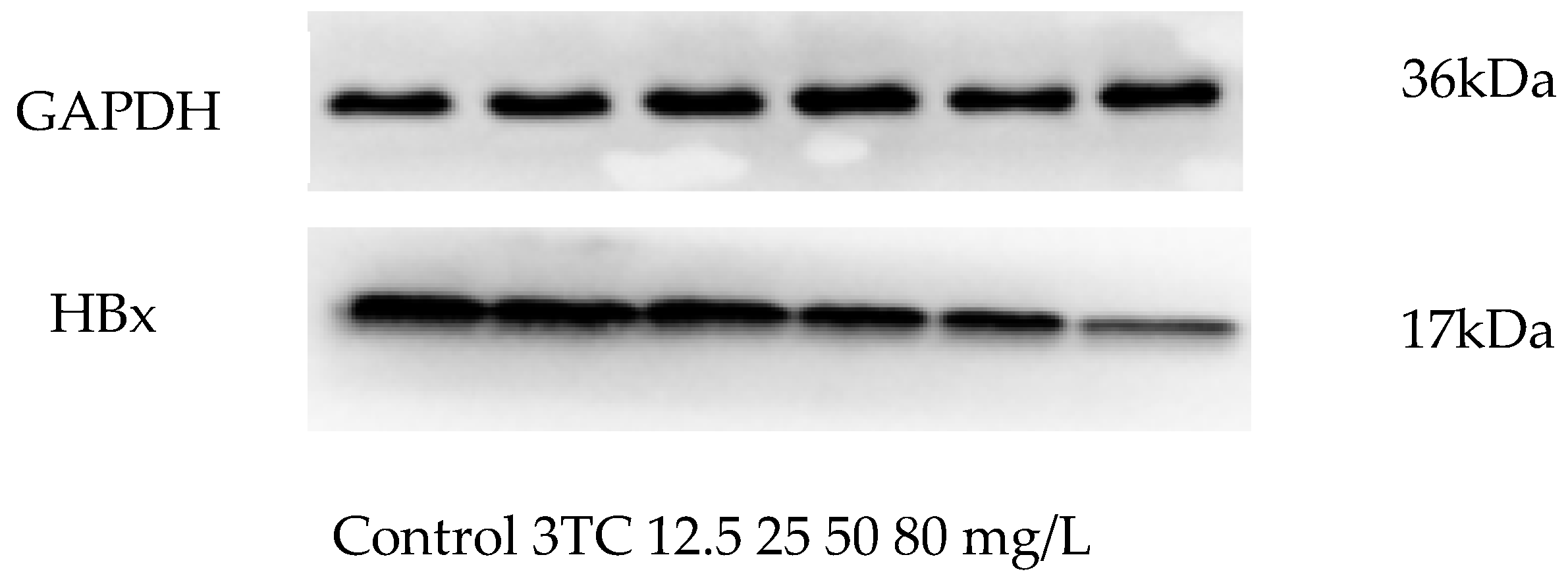
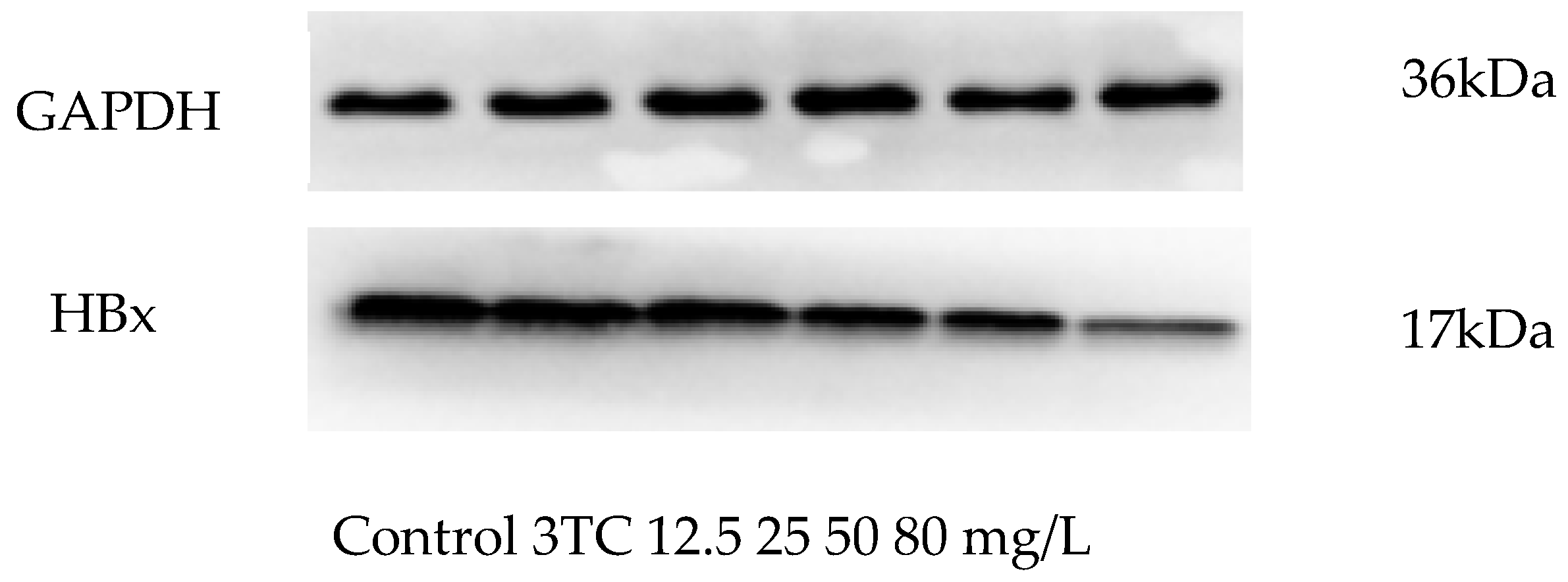
2.6. Effect of Esculetin on the Expression of HBx
2.7. Short-Term Toxicity of Esculetin on Ducklings
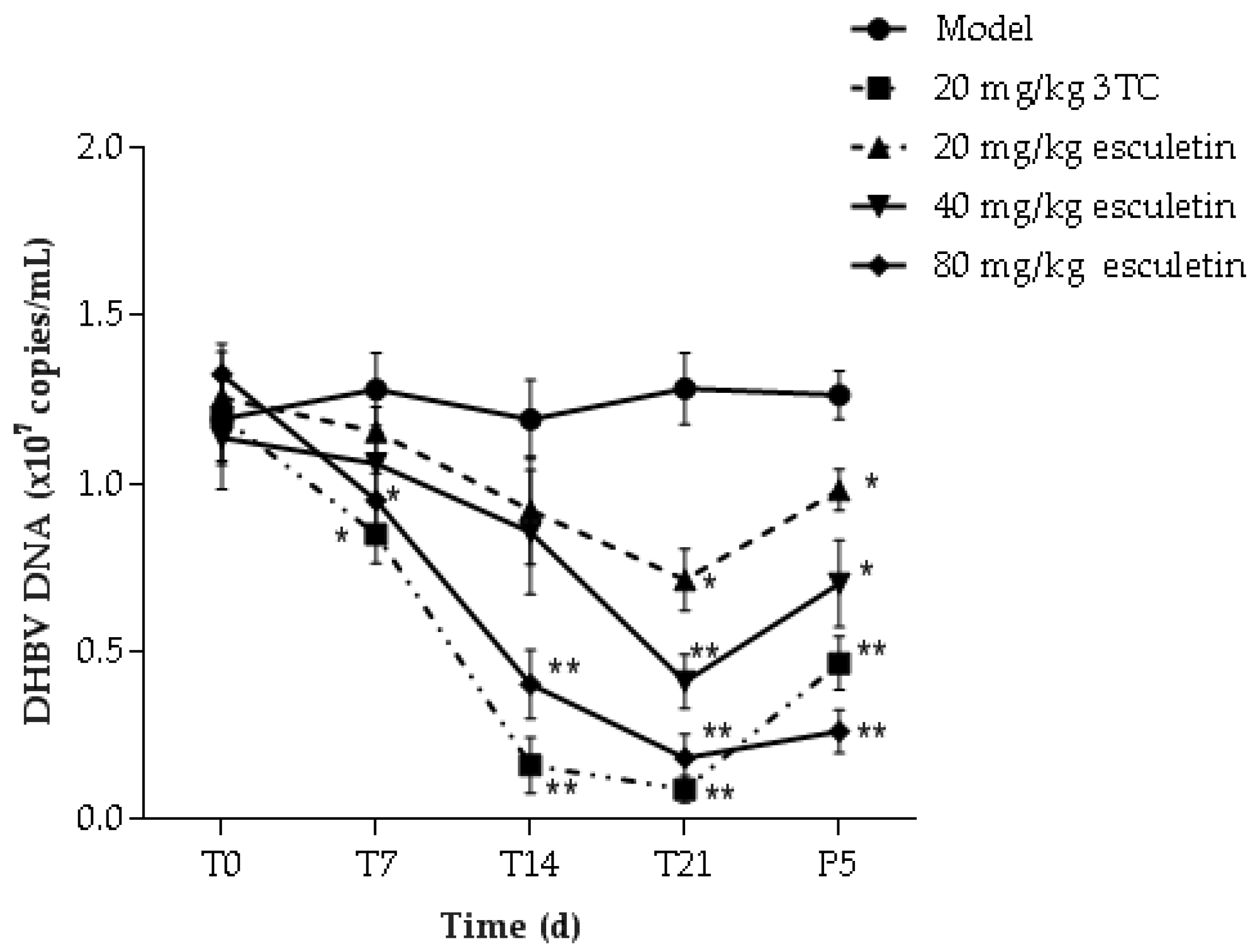
2.8. Effect of Esculetin on Serum Content of DHBV DNA
2.9. Effects of Esculetin on Serum DHBsAg and DHBeAg Levels
2.10. Analysis of ALT and AST Levels in Serum
2.11. Histopathological Examination of Duck Livers
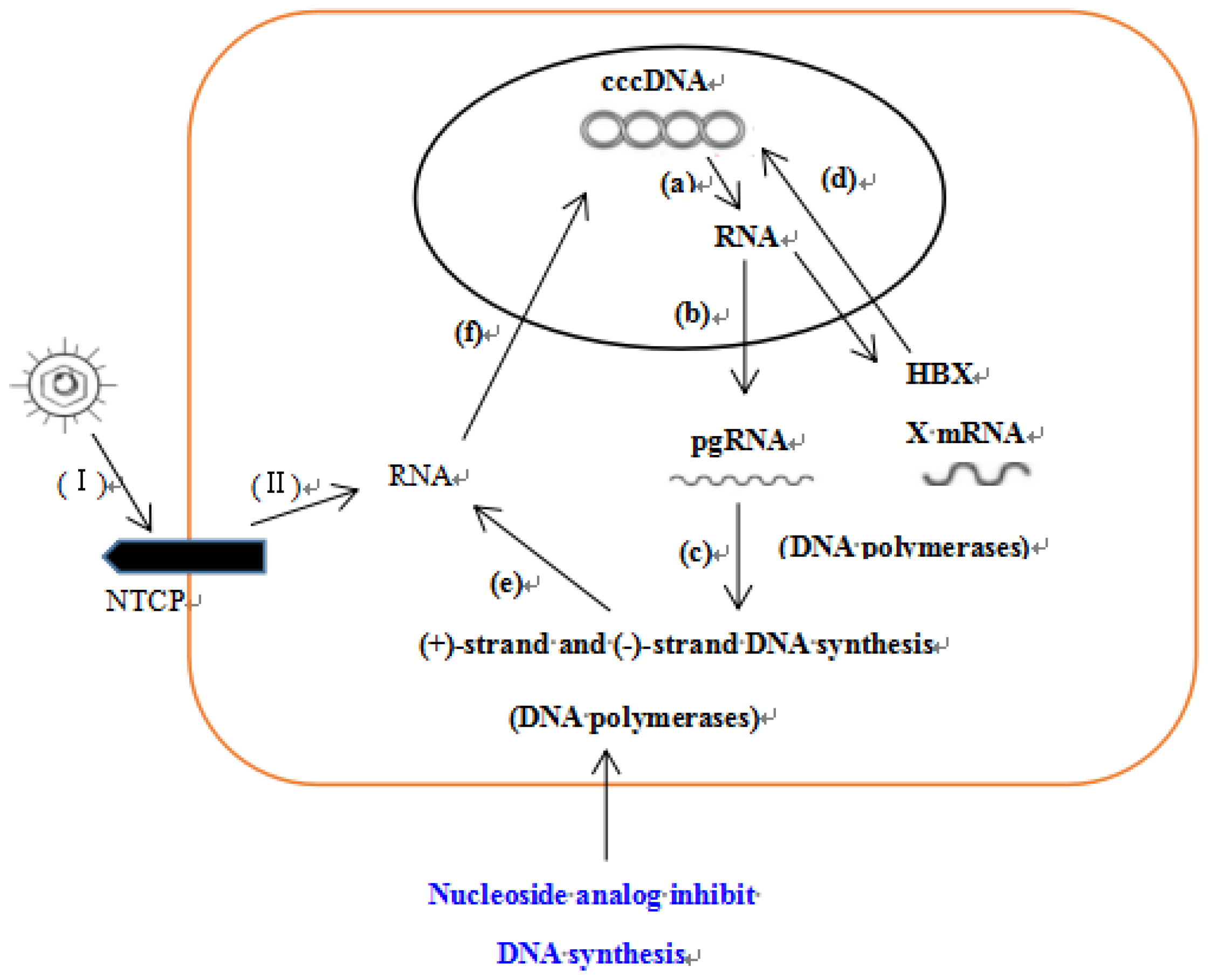
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. General Experimental Instruments
4.3. Plant Material
4.4. Extraction and Isolation
4.5. Cell Culture
4.6. Cytotoxicity Assay on HL-7702 Cells
4.7. Cytotoxicity Assay on HepG2.2.15 Cells
4.8. Detection of HBsAg and HBeAg in the Supernatant by ELISA
4.9. Determination of HBV DNA in the Supernatant by FQ-PCR (Fluorescent Probe Method)
4.10. Determination of HBx Protein by Western Blot
4.11. Short-Term Toxic Reaction of Esculetin on Ducklings
4.12. Animals and Treatments
4.13. Measurement of Serum DHBV DNA by FQ-PCR Assay
4.14. Measurement of Serum DHBsAg and DHBeAg
4.15. Analysis of Serum ALT and AST
4.16. Histopathological Examination of the Duck Livers
4.17. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ishtiaq, Q.; Kaneez, F.; Hany, A.H. Hepatitis B virus X protein impedes the DNA repair via its association with transcription factor, TFIIH. BMC Microbiol. 2011, 11, 1–15. [Google Scholar]
- Torres, C.; Leone, F.G.P.Y.; Pezzano, S.C.; Mbayed, V.A.; Campos, R.H. New perspectives on the evolutionary history of hepatitis B virus genotype F. Molecular Phylogenet Evol. 2011, 59, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.N.; Feng, M.; Gong, S.J.; Dai, Z.K.; Zhou, Y.Z.; Qing, X.V. The effects of extracts from Arenaria kansuensis on the levels of HBsAg and HBeAg in HepG2.2.15 cells. J. Chin. Med. Mater. 2008, 31, 1212–1216. [Google Scholar]
- Caviglia, G.P.; Abate, M.L.; Pellicano, R.; Smedile, A. Chronic hepatitis B therapy: Available drugs and treatment guidelines. MinervaI Gastroenterol Dietol. 2015, 61, 61–70. [Google Scholar]
- Zhang, G.M.; Sun, X.J.; Wang, F.Z. Analysis of epidemiological characteristics of hepatitis B among the population of 18–59 year old and HepB immunization strategies. Chin. J. Vaccines Immun. 2013, 19, 266–270. [Google Scholar]
- Hou, J.; Wang, G.; Wang, F.; Cheng, J.; Hong, R.; Hui, Z.; Jian, S.; Li, L.; Jie, L.; Meng, Q. Guideline of prevention and treatment for chronic hepatitis B. J. Clin. Transl. Hepatol. 2017, 5, 297–318. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.P.; Zhou, D.Q.; Xiong, Y.Q.; Gao, H.; He, J.S.; Chen, Y.J.; Wei, C.S.; Chen, L.; Xu, W.J.; Li, Q. Effect of dendritic cells and HBV-DNA to chronic hepatitis B patients with Liangxue Zishen Yiqi Method. Chin. J. Etmf. 2015, 21, 160–164. [Google Scholar]
- Wei, D.S.; Zeng, L.L.; Wang, Y.P.; Liang, K. Experiment on the propagation of spores of Microsorum fortunei. Chin. Herb. Med. 1999, 3, 224–225. [Google Scholar]
- Yao, Z.S.; Xu, X.R.; Chen, J.; Ge, F. Medicinal plant resources of Guanshan nature reserve in Jiangxi province. Subtrop. Plant Sci. 2005, 34, 43–47. [Google Scholar]
- Li, C.Y.; Ding, Y.J.; Li, J.; Zhao, A.G. Anti - tumor effect and mechanism of esculetin. Chin. Arch. Tradit. Chin. Med. 2019, 37, 1620–1623. [Google Scholar]
- Kim, W.K.; Byun, W.S.; Chung, H.J.; Oh, J.; Park, H.J.; Choi, J.S.; Lee, S.K. Esculetin suppresses tumor growth and metastasis by targeting Axin2/E-cadherin axis in colorectal cancer. Biochem. Pharm. 2018, 152, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Selvasundaram, R.; Manoharan, S.; Buddhan, R.; Neelakandan, M.; Murali, R. Chemopreventive potential of esculetin in 7,12-dimethylbenz(a)anthracene-induced hamster buccal pouch carcinogenesis. Mol. Cell. Biochem. 2018, 448, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Jeong, N.H.; Yang, E.J.; Jin, M.; Lee, J.Y.; Choi, Y.A.; Park, P.H.; Lee, S.R.; Kim, S.U.; Shin, T.Y.; Kwon, T.K.; et al. Esculetin from Fraxinus rhynchophylla attenuates atopic skin inflammation by inhibiting the expression of inflammatory cytokines. Int. Immunopharmacol. 2018, 59, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Raj, P.; Goru, S.K.; Kadakol, A.; Malek, V.; Sharma, N.; Gaikwad, A.B. Esculetin ameliorates hepatic fibrosis in high fat diet induced non-alcoholic fatty liver disease by regulation of FoxO1 mediated pathway. Pharmacol Rep. 2017, 69, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Yang, J.; Jeon, J.; Jeong, H.S.; Lee, J.; Sung, J. Hepatoprotective effect of esculetin on ethanol-induced liver injury in human HepG2 cells and C57BL/6J mice. J. Funct. Foods 2018, 40, 536–543. [Google Scholar] [CrossRef]
- Dhamodaran, P.; Natarajan, A. Protective effect of esculetin on hyperglycemia-mediated oxidative damage in the hepatic and renal tissues of experimental diabetic rats. Biochimie 2013, 95, 366–373. [Google Scholar]
- Tien, Y.C.; Liao, J.C.; Chiu, C.S.; Huang, T.H.; Huang, C.Y.; Chang, W.T.; Peng, W.H. Esculetin ameliorates carbon tetrachloride-mediated hepatic apoptosis in rats. Int. J. Mol Sci. 2011, 12, 4053–4067. [Google Scholar] [CrossRef]
- Wang, Q.; Dong, J.X. Functional progress in X gene of heptitis B virus and protein HBX. Lett Biotech. 2006, 17, 267–268. [Google Scholar]
- Cho, H.K.; Kim, S.Y.; Kyaw, Y.Y.; Win, A.A.; Koo, S.H.; Kim, H.H.; Cheong, J. HBx induces the proliferation of hepatocellular carcinoma cells via AP1 over-expressed as a result of ER stress. Biochem. J. 2015, 466, 115–121. [Google Scholar] [CrossRef]
- Lee, S.H.; Cha, E.J.; Lim, J.E.; Kwon, S.H.; Kim, D.H.; Cho, H.; Han, K.H. Structural characterization of an intrinsically unfolded mini-HBX protein from hepatitis B virus. Mol. Cells 2012, 34, 165–169. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Wang, L.W.; Yan, S.N.; Gong, Z.J. Effects of cell cycle on telomerase activity and on hepatitis B virus replication in HepG2 2.2.15 cells. Hepatobiliary Pancreat. Dis. Int. 2004, 3, 543–547. [Google Scholar] [PubMed]
- Gearhart, T.L.; Bouchard, M.J. The hepatitis B virus X protein modulates hepatocyte proliferation path-ways to stimulate viral replication. J. Virol. 2010, 84, 2675–2686. [Google Scholar] [CrossRef] [PubMed]
- Su, H.L.; Huang, R.D.; He, S.Q.; Xu, Q.; Zhu, H.; Mo, Z.J.; Liu, Q.B.; Liu, Y.M. Cloning and sequence analysis of the DHBV genome of the brown ducks in guilin region and establishment of the quantitative method for detecting DHBV. Chin. J. Virol. 2013, 29, 180. [Google Scholar]
- Lu, X.L.; Qiao, Y.; Zhang, X.M.; Ma, B.L.; Qiu, M.H. Chemical Constituents from Ceratophyllum demersum (Ceratophyllaceae). Acta Bot. Yunnanica 2007, 29, 263–264. [Google Scholar]
- Louis, K.S.; Siegel, A.C. Cell viability analysis using trypan blue: Manual and automated methods. Methods Mol. Biol. 2011, 740, 7–12. [Google Scholar]
- Tan, G.X.; Usonlopez, R.A.; Rahman, M.M.; Hosokawa, T.; Saito, T.; Kurasaki, M. Myricetin enhances on apoptosis induced by serum deprivation in PC12 cells mediated by mitochondrial signaling pathway. Environ. Toxicol. Pharm. 2018, 57, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.Y.; Liu, J.H.; Chang, Y. Fetal bovine serum can reduce toxicity of indocyanine green, brilliant blue G and trypan blue in ARPE-19 cellular model that suggests new surgical staining protocols for internal limiting membrane peeling procedure. Clin. Exp. Ophthalmol. 2018. [Google Scholar] [CrossRef]
- Ulrike, P. Hepatitis: Epigenetic control of HBV by HBx protein-releasing the break? Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 558–559. [Google Scholar]
- Sheng, L.; Wei, W.; Shi, K.; Cao, X.; Zhou, M.; Liu, Z. In vitro and in vivo anti-hepatitis B virus activities of the lignan niranthin isolated from Phyllanthus niruri L. J. Ethnopharmacol. 2014, 157, 62–68. [Google Scholar]
- Zhang, X.W.; Li, Y.J.; Fang, J.; Dou, J.J. The experimental research on SYR-SLN for the resistance of DHBV. Tradit. Chin. Med. 2011, 28, 107–110. [Google Scholar]
- Zhang, G.; Zeng, F.D.; Wang, T.C. Recent research progress in lamivudine, a new anti-HBV agent. Chin. J. New Drugs 2001, 10, 96–99. [Google Scholar]
- Chen, Y.P. Recent research on anti-hepatitis B virus therapy with lamivudine. Int. J. Virol. 1999, 6, 74–77. [Google Scholar]
- Ma, F.F.; Hu, Z.X.; Chen, J.; Liang, G.Y.; Xu, B.X. Chemical constituents of the leaves of magnolia grandiflora linn and their antitumor activities. Chin. Pharm. J. 2016, 51, 967–971. [Google Scholar]
- Yan, L.; Yu, H.H.; Liu, Y.S.; Wang, Y.S.; Zhao, W.H. Esculetin enhances the inhibitory effect of 5-Fluorouracil on the proliferation, migration and epithelial-mesenchymal transition of colorectal cancer. Cancer Biomark. Sect. A Dis. Markers 2019, 24, 231–240. [Google Scholar] [CrossRef]
- Choi, Y.J.; Lee, C.M.; Park, S.H.; Nam, M.J. Esculetin induces cell cycle arrest and apoptosis in human colon cancer LoVo cells. Environ. Toxicol. 2019. [Google Scholar] [CrossRef]
- Liang, M. Study on the isolation and activity of antioxidant components from cortex anserinal. Sci. Technol. Food Ind. 2006, 27, 64–66. [Google Scholar]
- Sun, Y.F.; Du, L.L.; Zhou, L.; Zhang, W.G.; Miao, F.; Yang, X.J.; Geng, H.L. Study on antibacterial active components of radix rehmanniae. China J. Chin. Mater. Med. 2011, 34, 88–90. [Google Scholar]
- Liang, C.Q.; Zhou, X.L.; Wang, P.C.; Tan, X.D.; Luo, Q.; Chen, X.; Pan, Z.H. Chemical constituents from stems and leaves of Microsorium fortunei. J. Chin. Med. Mater. 2017, 40, 2089–2092. [Google Scholar]
- Zhao, Y.; Geng, C.A.; Chen, H.; Ma, Y.B.; Huang, X.Y.; Cao, T.W.; He, K.; Wang, H.; Zhang, X.M.; Chen, J.J. Isolation, synthesis and anti-hepatitis B virus evaluation of p-hydroxyacetophenone derivatives from Artemisia capillaris. Bioorg Med. Chem. Lett. 2015, 25, 1509–1514. [Google Scholar] [CrossRef]
- An, R.B.; Sohn, D.H.; Kim, Y.C. Hepatoprotective compounds of the roots of Cudrania tricuspidata on tacrine-induced cytotoxicity in Hep G2 cells. Biol. Pharm. Bull. 2006, 29, 838–840. [Google Scholar] [CrossRef]
- Song, X.Y.; Wu, Y.H.; Hu, S.Q.; Hao, B.J.; Meng, Q.L. Inhibitory effect of aqueous extract of Leontopodium leontopodioides against d-galactosamine-induced hepatocyte injury. Biol. Pharm. Bull. 2006, 29, 838–840. [Google Scholar]
- Duflot, A.; Mehrotra, R.; Yu, S.Z.; Barraud, L.; Trepo, C.; Cova, L. Spectrum of liver disease and duck hepatitis B virus infection in a large series of chinese ducks with hepatocellular carcinoma. Hepatology 2010, 21, 1483–1491. [Google Scholar]
- Zhou, X.L.; Li, Y.X.; Zhang, S.J.; Liu, L.; Huang, R.B. Effect of taraffinisoside A on the expressions of HBsAg, HBeAg and HBV-DNA in HepG2.2.15 cells. Chin. Tradit. Pat. Med. 2015, 37, 1897–1900. [Google Scholar]
- Peng, Z.; Fang, G.; Peng, F.; Pan, Z.; Su, Z.; Tian, W.; Li, D.; Hou, H. Effects of Rubiadin isolated from Prismatomeris connata on anti-hepatitis B virus activity in vitro. Phytotherapy Research. 2017, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Huang, R.; Wei, L.; Chen, Y.; Lv, S.; Liang, C.; Zhang, X.; Yin, F.; Li, H.; Zhuo, L. Antiviral activity of methyl helicterate isolated from Helicteres angustifolia (Sterculiaceae) against hepatitis B virus. Antivir. Res. 2013, 100, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Bieche, I.; Olivi, M.; Champeme, M.; Vidaud, D.; Lidereau, R.; Vidaud, M. Novel approach to quantitative polymerase chain reaction using real-time detection: Application to the detection of gene amplification in breast cancer. Int. J. Cancer 1998, 78, 661–666. [Google Scholar] [CrossRef]
- Wei, P.H.; Wu, S.Z.; Mu, X.M.; Xu, B.; Su, Q.J.; Wei, J.L.; Yang, Y.; Qin, B.; Xie, Z.C. Effect of alcohol extract of Acanthus ilicifolius L. on anti-duck hepatitis B virus and protection of liver. J. Ethnopharmacol. 2015, 160, 1–5. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Sample of the compound esculetin is available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Concentration (mg/kg) | ALT | ||||
|---|---|---|---|---|---|---|
| T0 | T7 | T14 | T21 | P5 | ||
| Model | 89.67 ± 4.28 | 83.53 ± 5.5 | 85.25 ± 6.83 | 79.13 ± 3.35 | 77.00 ± 7.72 | |
| 3TC | 20 | 86.35 ± 5.39 | 62.95 ± 7.36 ** | 47.88 ± 4.47 ** | 39.45 ± 6.00 ** | 65.65 ± 5.42 |
| 80 | 77.97 ± 6.63 | 56.76 ± 4.16 ** | 45.56 ± 3.60 ** | 36.08 ± 7.59 ** | 49.99 ± 8.56 ** | |
| esculetin | 40 | 85.57 ± 5.92 | 68.73 ± 7.23 * | 58.88 ± 7.82 ** | 48.85 ± 6.31 ** | 60.87 ± 4.91 ** |
| 20 | 78.88 ± 4.91 | 75.59 ± 6.53 | 69.31 ± 9.06 * | 65.20 ± 7.09 * | 72.02 ± 7.02 | |
| Group | Concentration (mg/kg) | AST | ||||
|---|---|---|---|---|---|---|
| T0 | T7 | T14 | T21 | P5 | ||
| Model | 160.65 ± 17.38 | 163.18 ± 14.49 | 162.07 ± 13.14 | 167.24 ± 15.05 | 150.44 ± 11.20 | |
| 3TC | 20 | 175.98 ± 13.79 | 120.79 ± 12.92 ** | 94.86 ± 13.05 ** | 80.70 ± 12.11 ** | 110.40± 12.12 ** |
| 80 | 156.93 ± 14.72 | 114.04 ± 13.34 ** | 89.19 ± 11.74 ** | 70.52 ± 13.32 ** | 89.72 ± 14.55 ** | |
| esculetin | 40 | 165.83 ± 13.58 | 137.95 ± 10.88 * | 118.48 ± 15.59 ** | 109.74 ± 13.88 ** | 126.94 ± 15.92 ** |
| 20 | 169.19 ± 14.00 | 146.01 ± 14.36 * | 130.77 ± 14.54 ** | 117.08 ± 10.48 ** | 142.44 ± 12.43 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.-X.; Mou, J.-F.; Luo, Q.; Mo, Q.-H.; Zhou, X.-L.; Huang, X.; Xu, Q.; Tan, X.-D.; Chen, X.; Liang, C.-Q. Anti-Hepatitis B Virus Activity of Esculetin from Microsorium fortunei In Vitro and In Vivo. Molecules 2019, 24, 3475. https://doi.org/10.3390/molecules24193475
Huang S-X, Mou J-F, Luo Q, Mo Q-H, Zhou X-L, Huang X, Xu Q, Tan X-D, Chen X, Liang C-Q. Anti-Hepatitis B Virus Activity of Esculetin from Microsorium fortunei In Vitro and In Vivo. Molecules. 2019; 24(19):3475. https://doi.org/10.3390/molecules24193475
Chicago/Turabian StyleHuang, Si-Xin, Jun-Fei Mou, Qin Luo, Qing-Hu Mo, Xian-Li Zhou, Xiao Huang, Qing Xu, Xiang-Duan Tan, Xu Chen, and Cheng-Qin Liang. 2019. "Anti-Hepatitis B Virus Activity of Esculetin from Microsorium fortunei In Vitro and In Vivo" Molecules 24, no. 19: 3475. https://doi.org/10.3390/molecules24193475
APA StyleHuang, S.-X., Mou, J.-F., Luo, Q., Mo, Q.-H., Zhou, X.-L., Huang, X., Xu, Q., Tan, X.-D., Chen, X., & Liang, C.-Q. (2019). Anti-Hepatitis B Virus Activity of Esculetin from Microsorium fortunei In Vitro and In Vivo. Molecules, 24(19), 3475. https://doi.org/10.3390/molecules24193475