
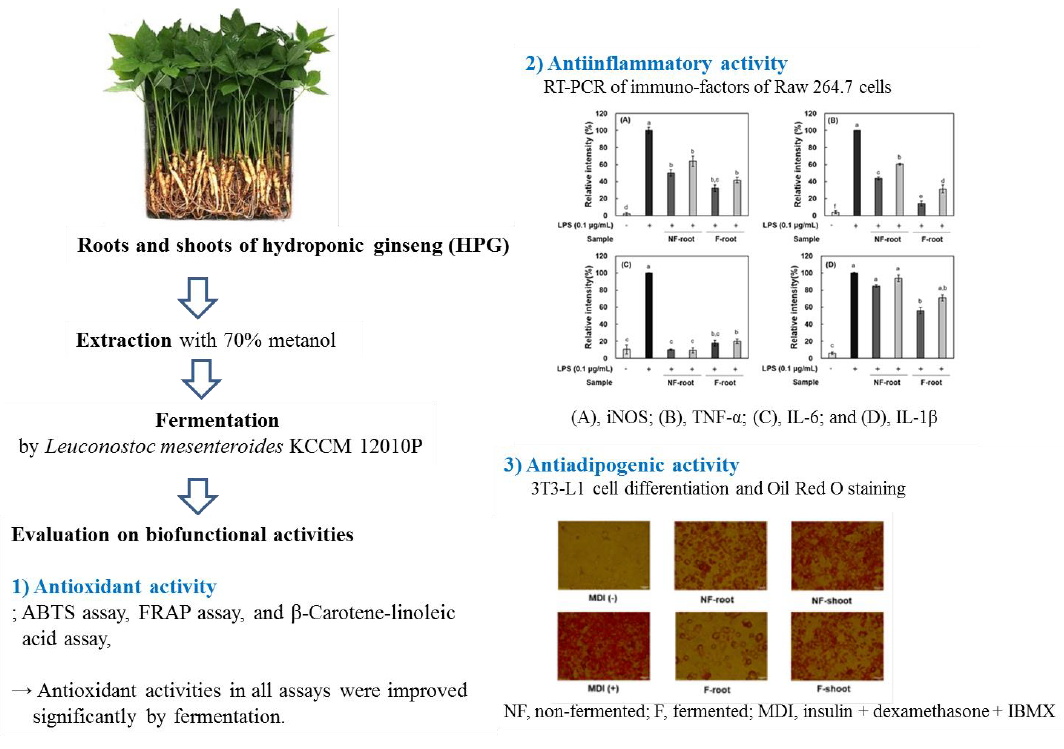
Improved Antioxidant, Anti-inflammatory, and Anti-adipogenic Properties of Hydroponic Ginseng Fermented by Leuconostoc mesenteroides KCCM 12010P


Abstract

1. Introduction
2. Results and Discussion
2.1. Enzymatic Activity of Leuconostoc Mesenteroides KCCM 12010P
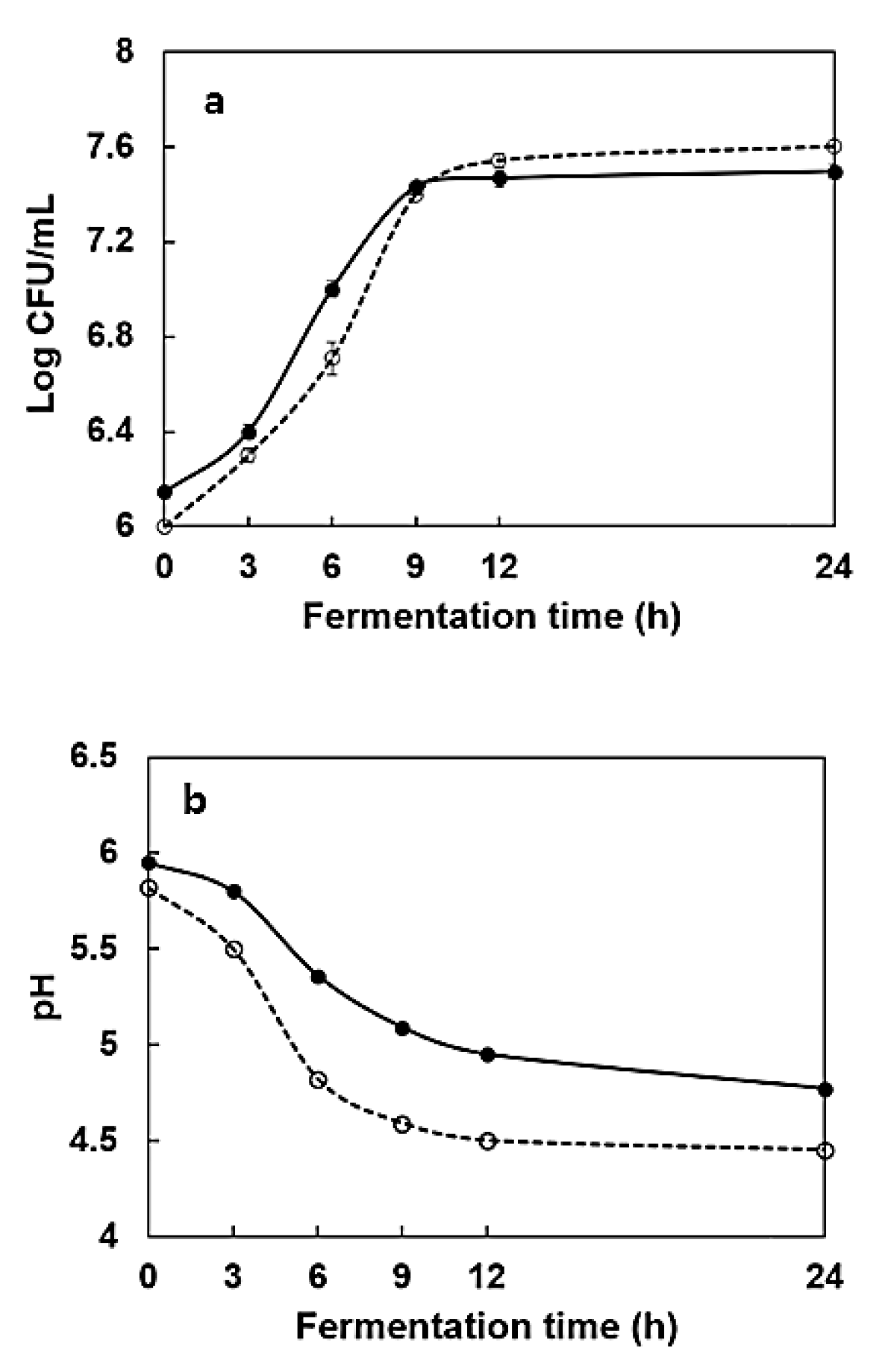
2.2. Changes in the Number of Cells and pH During Fermentation
2.3. Total Polyphenol and Flavonoid Content
2.4. Antioxidant Activity of Fermented Hydroponic Ginseng
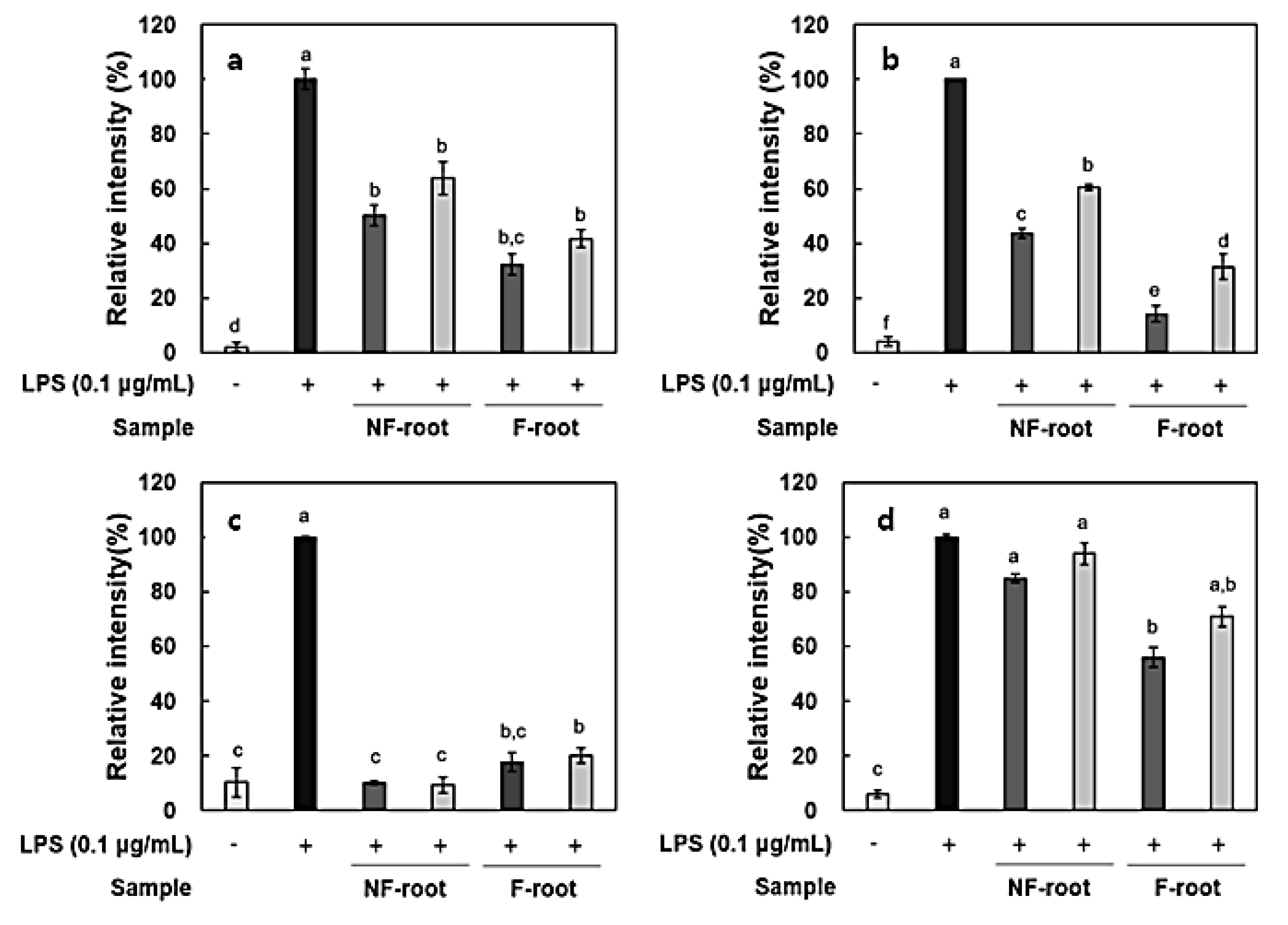
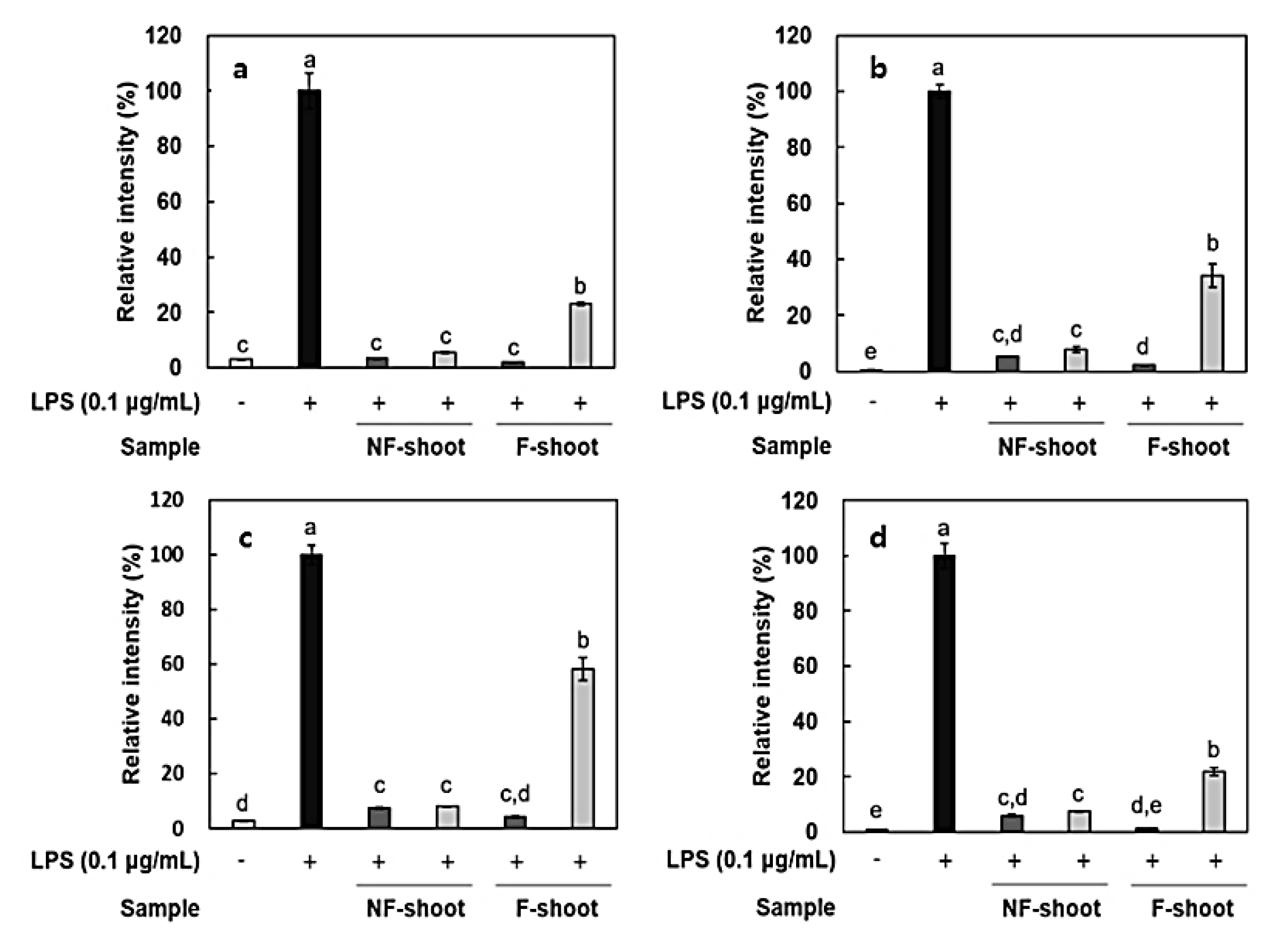
2.5. Anti-Inflammatory Effect of Non-fermented and Fermented Hydroponic Ginseng
2.6. Anti-Adipogenic Activity of Non-fermented and Fermented Hydroponic Ginseng
3. Materials and Methods
3.1. Materials
3.2. Microorganisms
3.3. Determination of Bacterial Production of Enzymes
3.4. Preparation for Hydroponic Ginseng Fermentation
3.5. Changes in Viable Number of Cells and pH during Fermentation
3.6. Measurement of Total Flavonoid Content
3.7. Measurement of Total Polyphenol Content
3.8. 2,2′-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS) Radical Scavenging Assay
3.9. Ferric Reducing Antioxidant Power (FRAP) Assay
3.10. β-Carotene-linoleic Acid Assay
3.11. Measurement of Nitric Oxide (NO) Production in RAW 264.7 Macrophage Cells
3.12. Measurement of Expression of Inflammation-Related Mediators
3.13. Cell Viability Assay
3.14. Cell Culture and Pre-Adipocyte Differentiation
3.15. Oil Red O Staining
3.16. Statistical Analysis
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Kim, J.; Byeon, H.; Im, K.; Min, H. Effects of ginsenosides on regulatory T cell differentiation. Food Sci. Biotechnol. 2018, 27, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.E.; Suh, D.H.; Kim, K.T.; Paik, H.D. Comparative study on anti-oxidative and anti-inflammatory properties of hydroponic ginseng and soil-cultured ginseng. Food Sci. Biotechnol. 2019, 28, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Garruti, D.S.; Franco, M.R.B.; da Silva, M.A.A.P.; Janzantti, N.S.; Alves, G.L. Assessment of aroma impact compounds in a cashew apple-based alcoholic beverage by GC–MS and GC-olfactometry. LWT- Food Sci. Technol. 2006, 39, 373–378. [Google Scholar] [CrossRef]
- Jung, J.; Jang, H.J.; Eom, S.J.; Choi, N.S.; Lee, N.K.; Paik, H.D. Fermentation of red ginseng extract by the probiotic Lactobacillus plantarum KCCM 11613P: ginsenoside conversion and antioxidant effects. J. Ginseng Res. 2019, 43, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.J.; Hwang, J.E.; Kim, H.S.; Kim, K.-T.; Paik, H.-D. Anti-inflammatory and cytotoxic effects of ginseng extract bioconverted by Leuconostoc mesenteroides KCCM 12010P isolated from kimchi. Int. J. Food Sci. Technol. 2018, 53, 1331–1337. [Google Scholar] [CrossRef]
- Gan, R.Y.; Shah, N.P.; Wang, M.F.; Lui, W.Y.; Corke, H. Lactobacillus plantarum WCFS1 fermentation differentially affects antioxidant capacity and polyphenol content in mung bean (Vigna radiata) and soya bean (Glycine max) milks. J. Food Process Preserv. 2017, 41, e12944. [Google Scholar] [CrossRef]
- Reyes-Gordillo, K.; Shat, R.; Muriel, P. Oxidative stress and inflammation in hepatic diseases: current and future therapy. Oxid. Med. Cell Longev. 2017. [Google Scholar] [CrossRef] [PubMed]
- Sim, M.O.; Jang, J.H.; Lee, H.E.; Jung, H.K.; Cho, H.W. Antioxidant effects of Geranium nepalense ethanol extract on H2O2-induced cytotoxicity in H9c2, SH-SY5Y, BEAS-2B, and HEK293. Food Sci. Biotechnol. 2017, 26, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Chae, G.N.; Kwank, S.J. NF-κB is involved in the TNF-α induced inhibition of the differentiation of 3T3-L1 cells by reducing PPARγ expression. Exp. Mol. Med. 2003, 35, 431–437. [Google Scholar] [CrossRef]
- Fernández-Sánchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, Á.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar]
- Song, Y.; Oh, G.H.; Kim, M.B.; Hwang, J.K. Fucosterol inhibits adipogenesis through the activation of AMPK and Wnt/beta-catenin signaling pathways. Food Sci. Biotechnol. 2017, 26, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Moller, D.E. Potential role of TNF-alpha in the pathogenesis of insulin resistance and type 2 diabetes. Trends Endocrinol. Metab. 2000, 11, 212–217. [Google Scholar] [CrossRef]
- Seo, J.Y.; Lee, J.H.; Kim, N.W.; Kim, Y.J.; Chan, S.H.; Ko, N.Y.; Her, E.; Yoo, Y.H.; Kim, J.W.; Lee, B.Y.; et al. Inhibitory effects of a fermented ginseng extract, BST204, on the expression of inducible nitric oxide synthase and nitric oxide production in lipopolysaccharide-activated murine macrophages. J. Pharm. Pharmacol. 2005, 57, 911–918. [Google Scholar] [CrossRef] [PubMed]
- Jeon, W.J.; Oh, J.S.; Park, M.S.; Ji, G.E. Anti-hyperglycemic effect of fermented ginseng in type 2 diabetes mellitus mouse model. Phytotherapy Res. 2013, 27, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.J.; Hwang, J.E.; Kim, K.-T.; Paik, H.-D. Increased antioxidative and nitric oxide scavenging activity of ginseng marc fermented by Pediococcus acidilactici KCCM11614P. Food Sci. Biotechnol. 2017, 27, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Lim, J.J.; Ahn, M.S.; Jeong, H.N.; An, T.J.; Kim, S.H. Comparative phenolic compound profiles and antioxidative activity of the fruit, leaves, and roots of Korean ginseng (Panax Ginseng Meyer) according to cultivation years. J. Ginseng Res. 2016, 40, 68–75. [Google Scholar] [CrossRef]
- Lee, K.S.; Seong, B.J.; Kim, G.H.; Kim, S.I.; Han, S.H.; Kim, H.H.; Baik, N.D. Ginsenoside, phenolic acid composition and physiological significances of fermented ginseng leaf. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1194–1200. [Google Scholar]
- Young, R.; Bush, S.J.; Lefevre, L.; McCulloch, M.E.B.; Lisowski, Z.M.; Muriuki, C.; Waddell, L.A.; Sauter, K.A.; Pridans, C.; Clark, E.L.; et al. Species-specific transcriptional regulation of genes involved in nitric oxide production and arginine metabolism in macrophages. ImmunoHorizons 2018, 2, 27–37. [Google Scholar] [CrossRef]
- Cho, J.Y.; Yoo, E.S.; Baik, K.U.; Park, M.H.; Han, B.H. In vitro inhibitory effect of protopanaxadiol ginsenosides on tumor necrosis factor (TNF)-α production and its modulation by known TNF-α antagonists. Planta Med. 2001, 67, 213–218. [Google Scholar] [CrossRef]
- Ye, R.; Yang, Q.; Kong, X.; Han, J.; Zhang, X.; Zhang, Y.; Li, P.; Liu, J.; Shi, M.; Xiong, L.; et al. Ginsenoside Rd attenuates early oxidative damage and sequential inflammatory response after transient focal ischemia in rats. Neurochem. Int. 2011, 58, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.W.; Do, H.J.; Kim, O.Y.; Chung, J.H.; Lee, J.Y.; Park, Y.S.; Hwang, K.Y.; Seong, S.I.; Shin, M.J. Fermented soy bean extract suppresses differentiation of 3T3-L1 preadipocytes and facilitates its glucose utilization. J. Funct. Food 2015, 15, 516–524. [Google Scholar] [CrossRef]
- Song, B.H.; Chan, Y.G.; Lee, K.A.; Lee, S.W.; Kang, S.W.; Cha, S.W. Studies on analysis of growth characteristics, ability of dry matter production, and yield of Panax ginseng C. A. Meyer at different growth stages with different cultivars and shading nets in paddy field. Korean J. Medicin Crop. Sci. 2011, 19, 90–96. [Google Scholar] [CrossRef]
- Jang, J.G.; Lee, K.S.; Kwon, D.W.; Oh, H.K. Chemical compositions of Korean ginseng with special reference to the part of ginseng plant. J. Ginseng Res. 1987, 6, 84–89. [Google Scholar]
- Oh, Y.J.; Kwon, S.H.; Choi, K.B.; Kim, T.S.; Yeo, I.H. Effect of vinegar made with hydroponic-cultured Panax ginseng C. A. Meyer on body weight and lipid metabolism in high-fat diet-fed mice. Korean Food Sci. Biotechnol. 2014, 46, 743–749. [Google Scholar] [CrossRef]
- Gülçin, İ.; Huyut, Z.; Elmastaş, M.; Aboul-Enein, H.Y. Radical scavenging and antioxidant activity of tannic acid. Arab. J. Chem. 2010, 3, 43–53. [Google Scholar] [CrossRef]
- Cha, B.J.; Park, J.H.; Shrestha, S.; Baek, N.I.; Lee, S.M.; Lee, T.H.; Kim, J.; Kim, G.S.; Kim, S.Y.; Lee, D.Y. Glycosyl glycerides from hydroponic Panax ginseng inhibited NO production in lipopolysaccharide-stimulated RAW264.7 cells. J. Ginseng Res. 2015, 39, 162–168. [Google Scholar] [CrossRef][Green Version]
- Liu, H.; Wang, J.; Liu, M.; Zhao, H.; Yaqoob, S.; Zheng, M.; Cai, D.; Liu, J. Antiobesity effectsof ginsenoside Rg1 on 3T3-L1 preadipocytes and high fat diet-induced obese mice mediated by AMPK. Nutrients 2018, 10, 830. [Google Scholar] [CrossRef]
- Nam, H.; Jung, H.; Kim, Y.; Kim, B.; Kim, K.H.; Park, S.J.; Suh, J.G. Aged black garlic extract regulates lipid metabolism by inhibiting lipogenesis and promoting lipolysis in mature 3T3-L1 adipocytes. Food Sci. Biotechnol. 2018, 27, 575–579. [Google Scholar] [CrossRef]
- Hsu, C.L.; Yen, G.C. Effects of flavonoids and phenolic acids on the inhibition of adipogenesis in 3T3-L1 adipocytes. J. Agric. Food Chem. 2007, 55, 8404–8410. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Activity 1) | Enzymes | Activity |
|---|---|---|---|
| Control | 0 | Acid phosphatase | 5 |
| Alkaline phosphatase | 5 | Naphthol-ASBI-phosphohydrolase | 10 |
| Esterase | 5 | α-Galactosidase | 20 |
| Esterase lipase | 0 | β-Galactosidase | ≥40 |
| Lipase | 0 | β-Glucuronidase | 0 |
| Leucine arylamidase | 10 | α-Glucosidase | 30 |
| Valine arylamidase | 0 | β-Glucosidase | ≥40 |
| Cystine arylamidase | 0 | N-Acetyl-β-glucosaminidase | 0 |
| Trypsin | 0 | α-Mannosidase | 0 |
| α-Chymotrypsin | 0 | α-Fucosidase | 0 |
| Sample | Total Polyphenol Content (GAE1)) | Total Flavonoid Content (KPE2)) | Antioxidant Activity | |||
|---|---|---|---|---|---|---|
| ABTS Radical Scavenging Activity (%) | FRAP Assay (μM Fe2+/mg Solid Sample) | Inhibition of β-carotene Oxidation (%) | ||||
| Non- fermented | Roots | 27.96 ± 0.35d | 0.68 ± 0.18d | 34.21 ± 0.36d | 89.47 ± 0.865d | 84.72 ± 0.72b |
| Shoots | 32.56 ± 0.40b | 13.74 ± 0.24b | 54.69 ± 1.20b | 214.76 ± 0.28b | 74.76 ± 0.67c | |
| Fermented | Roots | 29.97 ± 0.57c | 4.39 ± 0.10c | 46.41 ± 1.38c | 110.09 ± 1.27c | 88.20 ± 0.47a |
| Shoots | 40.11 ± 0.98a | 29.95 ± 0.46a | 62.30 ± 1.20a | 222.06 ± 0.22a | 76.76 ± 0.41d | |
| Forward | Reverse | |
|---|---|---|
| β-actin | 5′-GTCGGCCTAGGCACCAG-3′ | 5′-GGAGGAAGAGGATGCGGCAGT-3′ |
| iNOS | 5′-CCCTTCCGAACTTTCTGGCAGCAGC-3′ | 5′-GGCTGTCAGAGTCTCGTGGCTTTGG-3′ |
| TNF-α | 5′-GCAGAAGAGGCACTCCCCCA-3′ | 5′-GATCCATGCCGTTGGCCAGG-3′ |
| IL-1β | 5′-CAGGATGAGGACATGAGCACC-3′ | 5′-CTCTGCAGACTCAAACTCCAC-3′ |
| IL-6 | 5′-AGTTGCCTTCTTGGGACTGA-3′ | 5′-TTCTGCAAGTGCATCGT-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, J.E.; Kim, K.-T.; Paik, H.-D. Improved Antioxidant, Anti-inflammatory, and Anti-adipogenic Properties of Hydroponic Ginseng Fermented by Leuconostoc mesenteroides KCCM 12010P. Molecules 2019, 24, 3359. https://doi.org/10.3390/molecules24183359
Hwang JE, Kim K-T, Paik H-D. Improved Antioxidant, Anti-inflammatory, and Anti-adipogenic Properties of Hydroponic Ginseng Fermented by Leuconostoc mesenteroides KCCM 12010P. Molecules. 2019; 24(18):3359. https://doi.org/10.3390/molecules24183359
Chicago/Turabian StyleHwang, Ji Eun, Kee-Tae Kim, and Hyun-Dong Paik. 2019. "Improved Antioxidant, Anti-inflammatory, and Anti-adipogenic Properties of Hydroponic Ginseng Fermented by Leuconostoc mesenteroides KCCM 12010P" Molecules 24, no. 18: 3359. https://doi.org/10.3390/molecules24183359
APA StyleHwang, J. E., Kim, K.-T., & Paik, H.-D. (2019). Improved Antioxidant, Anti-inflammatory, and Anti-adipogenic Properties of Hydroponic Ginseng Fermented by Leuconostoc mesenteroides KCCM 12010P. Molecules, 24(18), 3359. https://doi.org/10.3390/molecules24183359