
Involvement of CB2 Receptors in the Neurobehavioral Effects of Catha Edulis (Vahl) Endl. (Khat) in Mice

 , ,
, ,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
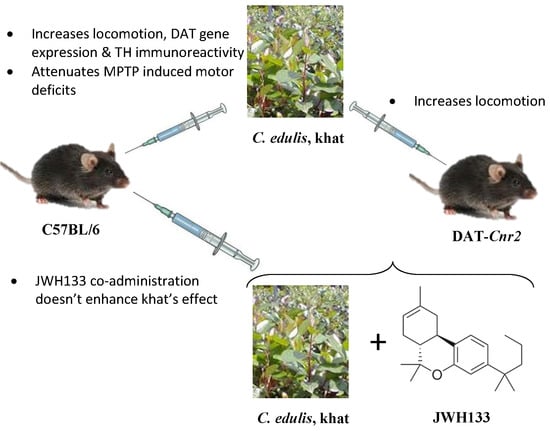
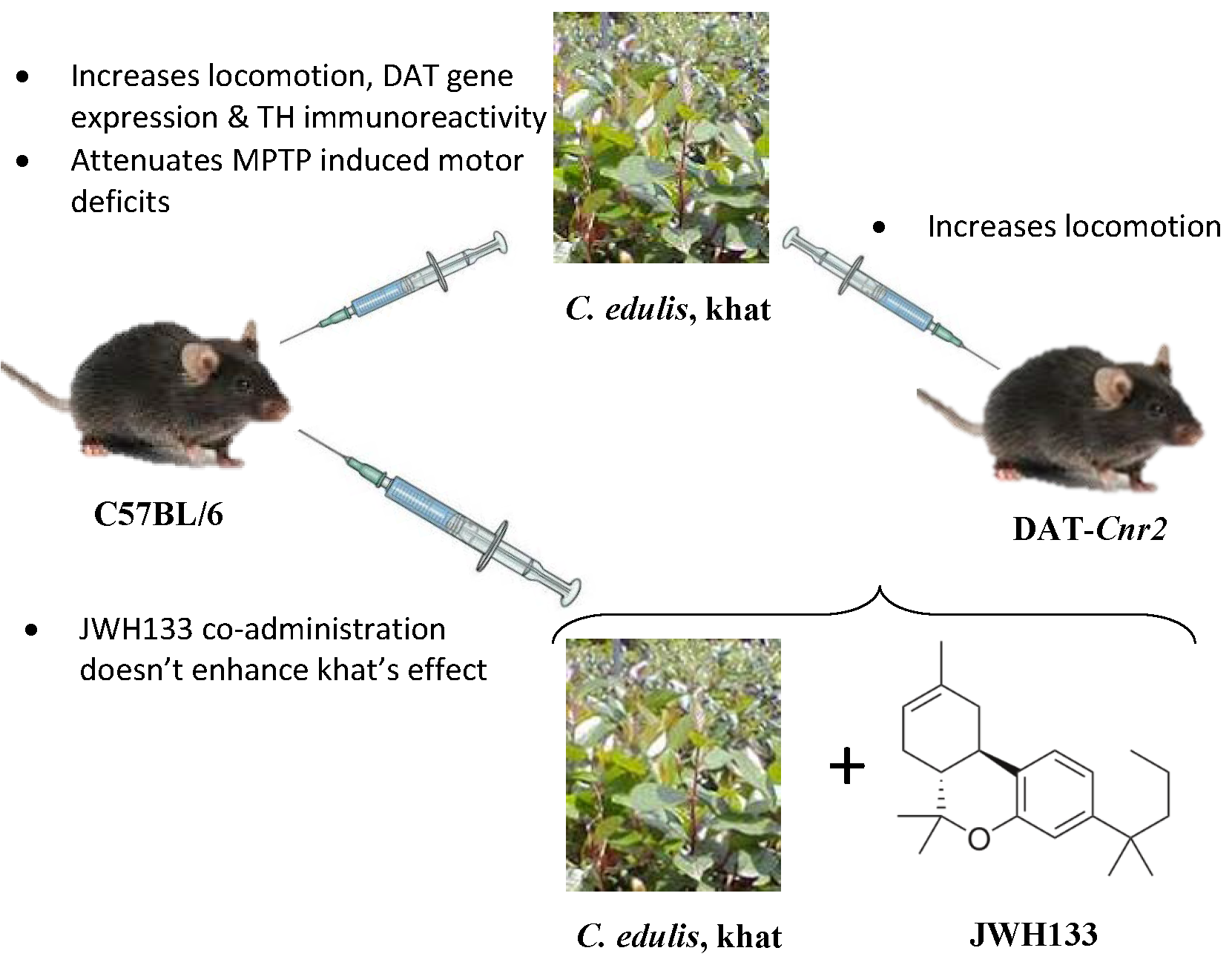
2.1. Khat Extract Increases Locomotor Activity in WT Mice
2.2. Deletion of CB2Rs in DA Neurons Augments Locomotor Activity of Khat Extract
2.3. Effect of JWH133 on Khat Extract′s Locomotor Activity
2.4. Khat Extract Attenuates MPTP-Induced Motor Deficits
2.5. JWH133 Increases Hyperlocomotor Effect of Khat Extract in MPTP Lesioned Mice
2.6. Effect of Khat Extract on Immunohistochemical Staining for TH-Positive Neurons in WT Mice
2.7. JWH133 Did Not Alter TH Immunoreactivity of Khat Extract
2.8. Effect of Khat Extract on DAT Mrna Gene Expression in WT Mice
2.9. Effect of Co-Administration of JWH133 on DAT Mrna Gene Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Chemicals
4.3. Collection and Extraction of Khat
4.4. Grouping and Dosing of Animals
4.5. MPTP-Induced Dopaminergic Lesion in Mice
4.6. Spontaneous Locomotor Activity Test
4.7. Immunohistochemical Staining for TH-Positive Neurons
4.8. Quantitative Reverse-Transcription (Qrt) PCR for Mrna Quantification
4.9. Sampling of VTA Tissue from Mice Midbrain
4.10. cDNA Synthesis
4.11. qRT-PCR Analysis of Relative mRNA Levels
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carlini, E.A. Plants and the central nervous system. Pharmacol. Biochem. Behav. 2003, 75, 501–512. [Google Scholar] [CrossRef]
- Connor, J.D.; Rostom, A.; Makonnen, E. Comparison of effects of khat extract and amphetamine on motor behaviors in mice. J. Ethnopharmacol. 2002, 81, 65–71. [Google Scholar] [CrossRef]
- Al-Hebshi, N.N.; Skaug, N. Khat (Catha edulis)-An updated review. Addict. Biol. 2005, 10, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, A.E.; Haughey, H.M.; Metzger, R.R.; Kokoshka, J.M.; Riddle, E.L.; Hanson, J.E.; Gibb, J.W.; Hanson, G.R. Differential effects of psychostimulants and related agents on dopaminergic and serotonergic transporter function. Eur. J. Pharmacol. 1999, 382, 45–49. [Google Scholar] [CrossRef]
- Feyissa, A.M.; Kelly, J.P. A review of the neuropharmacological properties of khat. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 1147–1166. [Google Scholar] [CrossRef] [PubMed]
- Engidawork, E. Pharmacological and Toxicological Effects of Catha edulis F. (Khat). Phyther. Res. 2017, 31, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Oyungu, E.; Kioy, P.G.; Patel, N.B. Effect of Catha edulis (khat) on behaviour and its potential to induce seizures in sprague Dawley rats. East. Afr. Med. J. 2007, 84, 219–225. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Geresu, B.; Onaivi, E.; Engidawork, E. Behavioral evidence for the interaction between cannabinoids and Catha edulis F. (Khat) in mice. Brain Res. 2016, 1648, 333–338. [Google Scholar] [CrossRef]
- Bedada, W.; Engidawork, E. The neuropsychopharmacological effects of Catha edulis in mice offspring born to mothers exposed during pregnancy and lactation. Phyther. Res. 2010, 24, 268–276. [Google Scholar]
- Onaivi, E.S.; Ishiguro, H.; Gu, S.; Liu, Q.R. CNS effects of CB2 cannabinoid receptors: Beyond neuro-immuno-cannabinoid activity. J. Psychopharmacol. 2012, 26, 92–103. [Google Scholar] [CrossRef]
- Little, P.J.; Compton, D.R.; Johnson, M.R.; Melvin, L.S.; Martin, B.R. Pharmacology and stereoselectivity of structurally novel cannabinoids in mice. J. Pharmacol. Exp. Ther. 1988, 247, 1046–1051. [Google Scholar]
- Burston, J.J.; Leggett, D.C.; Alekseeva, O.O.; Martin, B.R.; Wiley, J.L.; Mahadevan, A.; Razdan, R.K. CB 1 cannabinoid receptor-mediated modulation of food intake in mice. Br. J. Pharmacol. 2005, 145, 293–300. [Google Scholar]
- Ward, S.J.; Dykstra, L.A. The role of CB1 receptors in sweet versus fat reinforcement: Effect of CB1 receptor deletion, CB1 receptor antagonism (SR141716A) and CB1 receptor agonism (CP-55940). Behav. Pharmacol. 2005, 16, 381–388. [Google Scholar] [CrossRef]
- Riedel, G.; Davies, S.N. Cannabinoid function in learning, memory and plasticity. Handb. Exp. Pharmacol. 2005, 168, 445–477. [Google Scholar]
- Monory, K.; Blaudzun, H.; Massa, F.; Kaiser, N.; Lemberger, T.; Schütz, G.; Wotjak, C.T.; Lutz, B.; Marsicano, G. Genetic dissection of behavioural and autonomic effects of Δ 9-tetrahydrocannabinol in mice. PLoS Biol. 2007, 5. [Google Scholar] [CrossRef] [PubMed]
- von Rüden, E.L.; Jafari, M.; Bogdanovic, R.M.; Wotjak, C.T.; Potschka, H. Analysis in conditional cannabinoid 1 receptor-knockout mice reveals neuronal subpopulation-specific effects on epileptogenesis in the kindling paradigm. Neurobiol. Dis. 2015, 73, 334–347. [Google Scholar] [CrossRef] [PubMed]
- Monory, K.; Massa, F.; Egertová, M.; Eder, M.; Blaudzun, H.; Westenbroek, R.; Kelsch, W.W.; Jacob, W.; Marsch, R.; Ekker, M.; et al. The Endocannabinoid System Controls Key Epileptogenic Circuits in the Hippocampus. Neuron 2006, 51, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Steiner, M.A.; Marsicano, G.; Wotjak, C.T.; Lutz, B. Conditional cannabinoid receptor type 1 mutants reveal neuron subpopulation-specific effects on behavioral and neuroendocrine stress responses. Psychoneuroendocrinology 2008, 33, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Stempel, A.V.; Stumpf, A.; Zhang, H.Y.; Özdoğan, T.; Pannasch, U.; Theis, A.K.; Otte, D.M.; Wojtalla, A.; Rácz, I.; Ponomarenko, A.; et al. Cannabinoid Type 2 Receptors Mediate a Cell Type-Specific Plasticity in the Hippocampus. Neuron 2016, 90, 795–809. [Google Scholar] [CrossRef]
- Liu, Q.R.; Canseco-Alba, A.; Zhang, H.Y.; Tagliaferro, P.; Chung, M.; Dennis, E.; Sanabria, B.; Schanz, N.; Escosteguy-Neto, J.C.; Ishiguro, H.; et al. Cannabinoid type 2 receptors in dopamine neurons inhibits psychomotor behaviors, alters anxiety, depression and alcohol preference. Sci. Rep. 2017, 7, 17410. [Google Scholar] [CrossRef] [PubMed]
- Raitio, K.; Salo, O.; Nevalainen, T.; Poso, A.; Jarvinen, T. Targeting the cannabinoid CB2 receptor: Mutations, modeling and development of CB2 selective ligands. Curr. Med. Chem. 2005, 12, 1217–1237. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, A.; Parthier, D.; Sammons, R.P.; Stempel, A.V.; Breustedt, J.; Rost, B.R.; Schmitz, D. Cannabinoid type 2 receptors mediate a cell type-specific self-inhibition in cortical neurons. Neuropharmacology 2018, 139, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Canseco-Alba, A.; Schanz, N.; Sanabria, B.; Zhao, J.; Lin, Z.; Liu, Q.R.; Onaivi, E.S. Behavioral effects of psychostimulants in mutant mice with cell-type specific deletion of CB2 cannabinoid receptors in dopamine neurons. Behav. Brain Res. 2019, 360, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.J.; Xi, Z.-X. Progress in brain cannabinoid CB2 receptor research: From genes to behavior. Neurosci. Biobehav. Rev. 2019. [Google Scholar] [CrossRef] [PubMed]
- Brusco, A.; Tagliaferro, P.; Saez, T.; Onaivi, E.S. Postsynaptic localization of CB2 cannabinoid receptors in the rat hippocampus. Synapse 2008, 62, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; et al. Neuroscience: Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 2005, 310, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Brain neuronal CB2 cannabinoid receptors in drug abuse and depression: From mice to human subjects. PLoS ONE 2008, 3, e1640. [Google Scholar] [CrossRef]
- Fitzgerald, M.L.; Shobin, E.; Pickel, V.M. Cannabinoid modulation of the dopaminergic circuitry: Implications for limbic and striatal output. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 38, 21–29. [Google Scholar] [CrossRef]
- Banjaw, M.Y.; Schmidt, W.J. Behavioural sensitisation following repeated intermittent oral administration of Catha edulis in rats. Behav. Brain Res. 2005, 156, 181–189. [Google Scholar] [CrossRef]
- Kimani, S.T.; Nyongesa, A.W. Effects of single daily khat (Catha edulis) extract on spatial learning and memory in CBA mice. Behav. Brain Res. 2008, 195, 192–197. [Google Scholar] [CrossRef]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G. Dopamine receptors: From structure to function. Physiol Rev. 1998, 78, 189–225. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Gainetdinov, R.R.; Fumagalli, F.; Xu, F.; Jones, S.R.; Bock, C.B.; Miller, G.W.; Wightman, R.M.; Caron, M.G. Knockout of the vesicular monoamine transporter 2 gene results in neonatal death and supersensitivity to cocaine and amphetamine. Neuron 1997, 19, 1285–1296. [Google Scholar] [CrossRef]
- Amara, S.G.; Sonders, M.S. Neurotransmitter transporters as molecular targets for addictive drugs. Drug Alcohol Depend. 1998, 51, 87–96. [Google Scholar] [CrossRef]
- Javitch, J.A.; Uhl, G.R.; Snyder, S.H. Parkinsonism-inducing neurotoxin, N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine: Characterization and localization of receptor binding sites in rat and human brain. Proc. Natl. Acad. Sci. USA 1984, 81, 4591–4595. [Google Scholar] [CrossRef]
- Scavone, J.L.; Sterling, R.C.; Van Bockstaele, E.J. Cannabinoid and opioid interactions: Implications for opiate dependence and withdrawal. Neuroscience 2013, 248, 637–654. [Google Scholar] [CrossRef] [PubMed]
- Werling, L.L.; Reed, S.C.; Wade, D.; Izenwasser, S. Chronic nicotine alters cannabinoid-mediated locomotor activity and receptor density in periadolescent but not adult male rats. Int. J. Dev. Neurosci. 2009, 27, 263–269. [Google Scholar] [CrossRef]
- Viveros, M.P.; Marco, E.M.; File, S.E. Nicotine and cannabinoids: Parallels, contrasts and interactions. Neurosci. Biobehav. Rev. 2006, 30, 1161–1181. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Peng, X.; Li, X.; Song, R.; Zhang, H. Brain Cannabinoid CB2 Receptors Modulate Cocaine′s Actions in Mice. Nat. Neurosci. 2015, 8, 1699–1712. [Google Scholar]
- Peters, D.W.A. Khat: Its history, botany, chemistry and toxicology. Pharm. J. 1952, 169, 16–18. [Google Scholar]
- Al-Habori, M. The potential adverse effects of habitual use of Catha edulis (khat). Expert Opin. Drug Saf. 2005, 4, 1145–1154. [Google Scholar] [CrossRef]
- Cox, G.; Rampes, H. Adverse effects of khat: A review. Adv. Psychiatr. Treat. 2003, 9, 456–463. [Google Scholar] [CrossRef]
- Warfa, N.; Klein, A.; Bhui, K.; Leavey, G.; Craig, T.; Alfred Stansfeld, S. Khat use and mental illness: A critical review. Soc. Sci. Med. 2007, 65, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M. Chronic khat use and psychotic disorders: A review of the literature and. Sucht 2007, 53, 9–22. [Google Scholar] [CrossRef]
- Kalix, P. Hypermotility of the Amphetamine Type Induced by a Constituent of Khat Leaves. Br. J. Pharmacol. 1980, 68, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Geresu, B.; Engidawork, E. Catha edulis F. (Khat) Reverses Haloperidol but Not Morphine Induced Motor Deficits Following Acute and Subacute Administration in Mice. Ethiop. Pharm. J. 2012, 28, 117–130. [Google Scholar] [CrossRef]
- Hassan, N.; Gunaid, A.; Murray-Lyon, I. Khat (Catha edulis): Health aspects of khat chewing. East. Mediterr. Health J. 2007, 13, 706–718. [Google Scholar]
- Chaperon, F.; Thiebot, M.-H. Behavioral Effects of Cannabinoid Agents in Animals. Crit. Rev. Neurobiol. 2017, 13, 243–281. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Lastres-Becker, I.; Cabranes, A.; González, S.; Ramos, J.A. Endocannabinoids and basal ganglia functionality. Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 257–267. [Google Scholar] [CrossRef]
- Stella, N. Cannabinoid and cannabinoid-like receptors in microglia, astrocytes, and astrocytomas. Glia 2010, 58, 1017–1030. [Google Scholar] [CrossRef]
- Gong, J.P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 receptors: Immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef]
- Baek, J.H.; Zheng, Y.; Darlington, C.L.; Smith, P.F. Cannabinoid CB2 receptor expression in the rat brainstem cochlear and vestibular nuclei. Acta Otolaryngol. 2008, 128, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, M.B.; Suetterlin, P.; Yip, P.; Molina-Holgado, F.; Walker, D.J.; Oudin, M.J.; Zentar, M.P.; Pollard, S.; Yáñez-Muñoz, R.J.; Williams, G.; et al. A diacylglycerol lipase-CB2 cannabinoid pathway regulates adult subventricular zone neurogenesis in an age-dependent manner. Mol. Cell. Neurosci. 2008, 38, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Viscomi, M.T.; Oddi, S.; Latini, L.; Pasquariello, N.; Florenzano, F.; Bernardi, G.; Molinari, M.; Maccarrone, M. Selective CB2 Receptor Agonism Protects Central Neurons from Remote Axotomy-Induced Apoptosis through the PI3K/Akt Pathway. J. Neurosci. 2009, 29, 4564–4570. [Google Scholar] [CrossRef] [PubMed]
- Sagredo, O.; González, S.; Aroyo, I.; Pazos, M.R.; Benito, C.; Lastres-Becker, I.; Romero, J.P.; Tolón, R.M.; Mechoulam, R.; Brouillet, E.; et al. Cannabinoid CB 2 receptor agonists protect the striatum against malonate toxicity: Relevance for Huntington′s disease. Glia 2009, 57, 1154–1167. [Google Scholar] [CrossRef] [PubMed]
- den Boon, F.S.; Chameau, P.; Schaafsma-Zhao, Q.; van Aken, W.; Bari, M.; Oddi, S.; Kruse, C.G.; Maccarrone, M.; Wadman, W.J.; Werkman, T.R. Excitability of prefrontal cortical pyramidal neurons is modulated by activation of intracellular type-2 cannabinoid receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 3534–3539. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Vincent, S.R. Histochemistry of MPTP oxidation in the rat brain: Sites of synthesis of the parkinsonism-inducing toxin MPP+. Neurosci. Lett. 1986, 65, 321–325. [Google Scholar] [CrossRef]
- Kopin, I.; Markey, S. MPTP toxicity: Implications for research in Parkinson′s disease. Annu. Rev. Neurosci. 1988, 11, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Shin, W.H.; Baek, J.Y.; Cho, E.J.; Baik, H.H.; Kim, S.R.; Won, S.Y.; Jin, B.K. CB2 receptor activation prevents glial-derived neurotoxic mediator production, BBB leakage and peripheral immune cell infiltration and rescues dopamine neurons in the MPTP model of Parkinson′s disease. Exp. Mol. Med. 2016, 48. [Google Scholar] [CrossRef]
- Shi, J.; Cai, Q.; Zhu, R.; Zhang, J.; Liu, Y.; He, X.; Jin, L. AM1241 alleviates MPTP-induced Parkinson′s disease and promotes the regeneration of DA neurons in PD mice. Oncotarget 2017, 8. [Google Scholar] [CrossRef]
- Walter, L.; Franklin, A.; Witting, A.; Wade, C.; Xie, Y.; Kunos, G.; Mackie, K.; Stella, N. Nonpsychotropic Cannabinoid Receptors Regulate Microglial Cell Migration. J. Neurosci. 2003, 23, 1398–1405. [Google Scholar] [CrossRef]
- Ramirez, B.G. Prevention of Alzheimer′s Disease Pathology by Cannabinoids: Neuroprotection Mediated by Blockade of Microglial Activation. J. Neurosci. 2005, 25, 1904–1913. [Google Scholar] [CrossRef] [PubMed]
- Price, D.A.; Martinez, A.A.; Seillier, A.; Koek, W.; Acosta, Y.; Fernandez, E.; Strong, R.; Lutz, B.; Marsicano, G.; Roberts, J.L.; et al. WIN55,212-2, a cannabinoid receptor agonist, protects against nigrostriatal cell loss in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson′s disease. Eur. J. Neurosci. 2009, 29, 2177–2186. [Google Scholar] [CrossRef] [PubMed]
- Grace, A.A.; Onn, S.P. Morphology and electrophysiological properties of immunocytochemically identified rat dopamine neurons recorded in vitro. J. Neurosci. 1989, 9, 3463–3481. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.W.; North, R.A. Two types of neurone in the rat ventral tegmental area and their synaptic inputs. J. Physiol. 1992, 450, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Lacey, M.G.; Mercuri, N.B.; North, R.A. Two cell types in rat substantia nigra zona compacta distinguished by membrane properties and the actions of dopamine and opioids. J. Neurosci. 1989, 9, 1233–1241. [Google Scholar] [CrossRef]
- Keath, J.R.; Iacoviello, M.P.; Barrett, L.E.; Mansvelder, H.D.; McGehee, D.S. Differential Modulation by Nicotine of Substantia Nigra Versus Ventral Tegmental Area Dopamine Neurons. J. Neurophysiol. 2007, 98, 3388–3396. [Google Scholar] [CrossRef] [PubMed]
- Daubner, S.C.; Le, T.; Wang, S. Tyrosine hydroxylase and regulation of dopamine synthesis. Arch. Biochem. Biophys. 2011, 508, 1–12. [Google Scholar] [CrossRef]
- Gordon, S.L.; Quinsey, N.S.; Dunkley, P.R.; Dickson, P.W. Tyrosine hydroxylase activity is regulated by two distinct dopamine-binding sites. J. Neurochem. 2008, 106, 1614–1623. [Google Scholar] [CrossRef]
- Zelger, J.L.; Schorno, X.; Carlini, E.A. Behavioural effects of cathinone, an amine obtained from Catha edulis Forsk.: Comparisons with amphetamine, norpseudoephedrine, apomorphine and nomifensine. Bull. Narc. 1980, 32, 67–81. [Google Scholar]
- Siciliano, C.A.; Calipari, E.S.; Ferris, M.J.; Jones, S.R. Adaptations of presynaptic dopamine terminals induced by psychostimulant self-administration. ACS Chem. Neurosci. 2015, 6, 27–36. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Canseco-Alba, A.; Sanabria, B.D. Identification of cannabinoid CB2 receptor neuro-immune crosstalk following conditional deletion of type 2 cannabinoid receptors in microglia and dopamine neurons. In Proceedings of the Society for Neuroscience-Abstract, San Diego, CA, USA, 3–7 November 2018. [Google Scholar]
- Furman, C.A.; Chen, R.; Guptaroy, B.; Zhang, M.; Holz, R.W.; Gnegy, M. Dopamine and amphetamine rapidly increase dopamine transporter trafficking to the surface: Live-cell imaging using total internal reflection fluorescence microscopy. J. Neurosci. 2009, 29, 3328–3336. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Guptaroy, B.; Lund, D.; Shamban, S.; Gnegy, M.E. Regulation of amphetamine-stimulated dopamine efflux by protein kinase C β. J. Biol. Chem. 2005, 280, 10914–10919. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Furman, C.A.; Zhang, M.; Guptaroy, B.; Gnegy, M.E. Rapid delivery of the dopamine transporter to the plasmalemmal membrane upon amphetamine stimulation. Neuropharmacology 2005, 49, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Rayport, S. Amphetamine and other psychostimulants reduce pH gradients in midbrain dopaminergic neurons and chromaffin granules: A mechanism of action. Neuron 1990, 5, 797–808. [Google Scholar] [CrossRef]
- Zhu, J.; Reith, M. Role of the Dopamine Transporter in the Action of Psychostimulants, Nicotine, and Other Drugs of Abuse. CNS Neurol. Disord. Drug Targets 2008, 7, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Admassie, E.; Engidawork, E. Subchronic administration of Catha edulis F. (khat) extract is marked by elevation of cardiac biomarkers and subendocardial necrosis besides blood pressure alteration in rats. J. Ethnopharmacol. 2011, 136, 246–253. [Google Scholar] [CrossRef]
- Meredith, G.E.; Rademacher, D.J. MPTP mouse models of Parkinson′s disease: An update. J. Parkinsons. Dis. 2011, 1, 19–33. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geresu, B.; Canseco-Alba, A.; Sanabria, B.; Lin, Z.; Liu, Q.-R.; Onaivi, E.S.; Engidawork, E. Involvement of CB2 Receptors in the Neurobehavioral Effects of Catha Edulis (Vahl) Endl. (Khat) in Mice. Molecules 2019, 24, 3164. https://doi.org/10.3390/molecules24173164
Geresu B, Canseco-Alba A, Sanabria B, Lin Z, Liu Q-R, Onaivi ES, Engidawork E. Involvement of CB2 Receptors in the Neurobehavioral Effects of Catha Edulis (Vahl) Endl. (Khat) in Mice. Molecules. 2019; 24(17):3164. https://doi.org/10.3390/molecules24173164
Chicago/Turabian StyleGeresu, Berhanu, Ana Canseco-Alba, Branden Sanabria, Zhicheng Lin, Qing-Rong Liu, Emmanuel S. Onaivi, and Ephrem Engidawork. 2019. "Involvement of CB2 Receptors in the Neurobehavioral Effects of Catha Edulis (Vahl) Endl. (Khat) in Mice" Molecules 24, no. 17: 3164. https://doi.org/10.3390/molecules24173164
APA StyleGeresu, B., Canseco-Alba, A., Sanabria, B., Lin, Z., Liu, Q.-R., Onaivi, E. S., & Engidawork, E. (2019). Involvement of CB2 Receptors in the Neurobehavioral Effects of Catha Edulis (Vahl) Endl. (Khat) in Mice. Molecules, 24(17), 3164. https://doi.org/10.3390/molecules24173164