Optimization of Degradation Conditions with PRG, a Polysaccharide from Phellinus ribis, by RSM and the Neuroprotective Activity in PC12 Cells Damaged by Aβ25–35

 ,
,
Abstract
1. Introduction
2. Results and Discussion
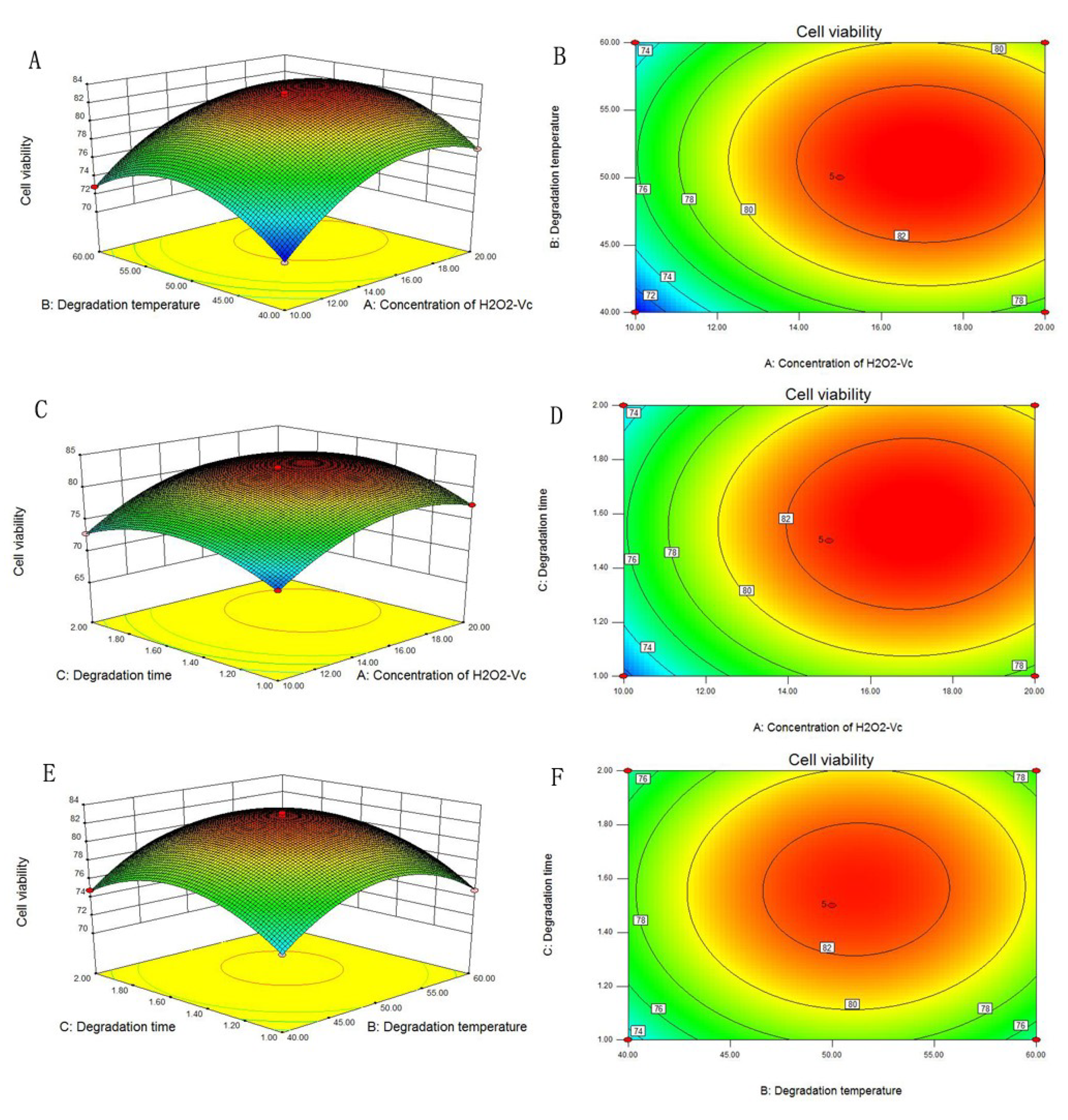
2.1. Optimization of the Degradation Conditions by RSM
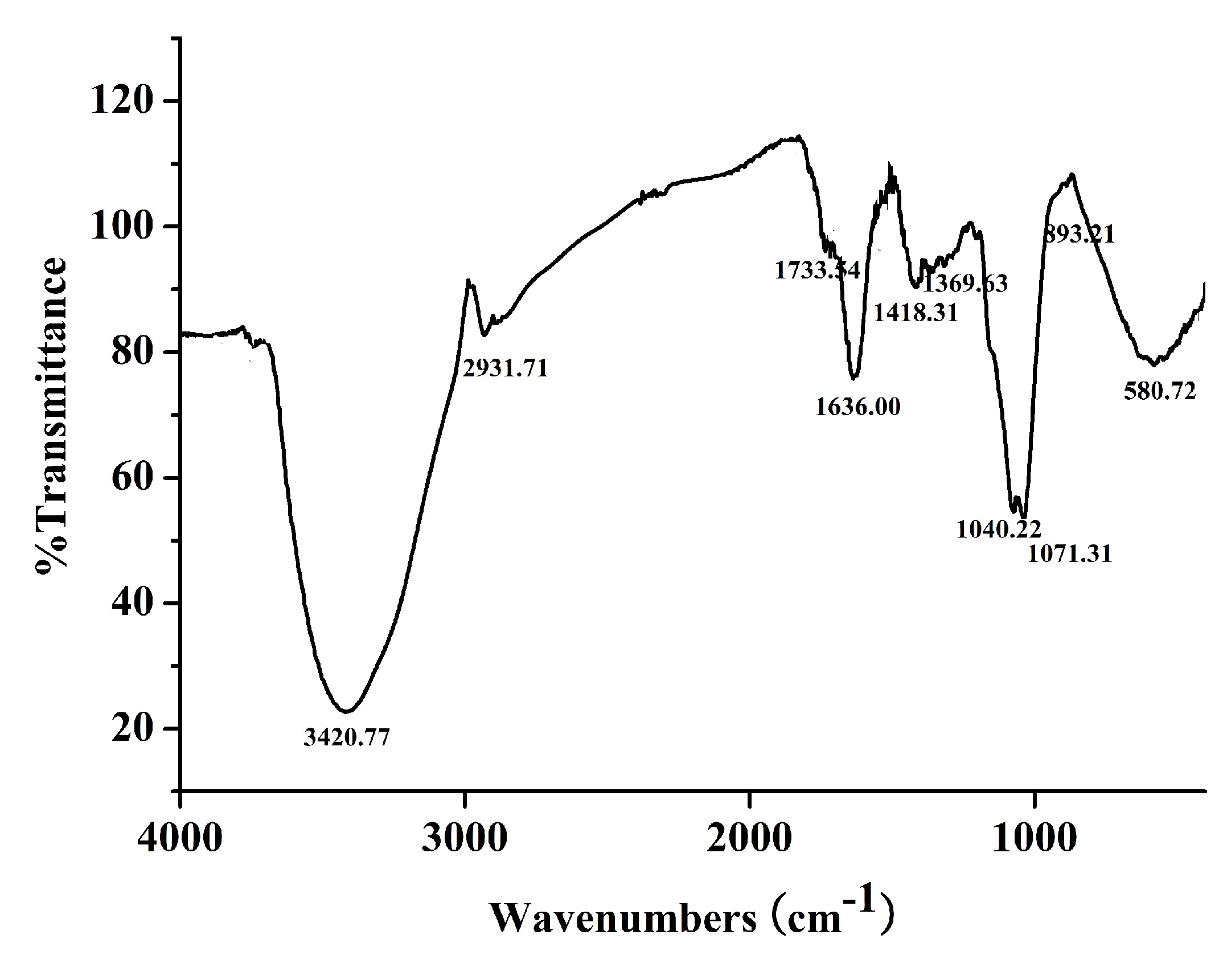
2.2. Characterization of DPRG
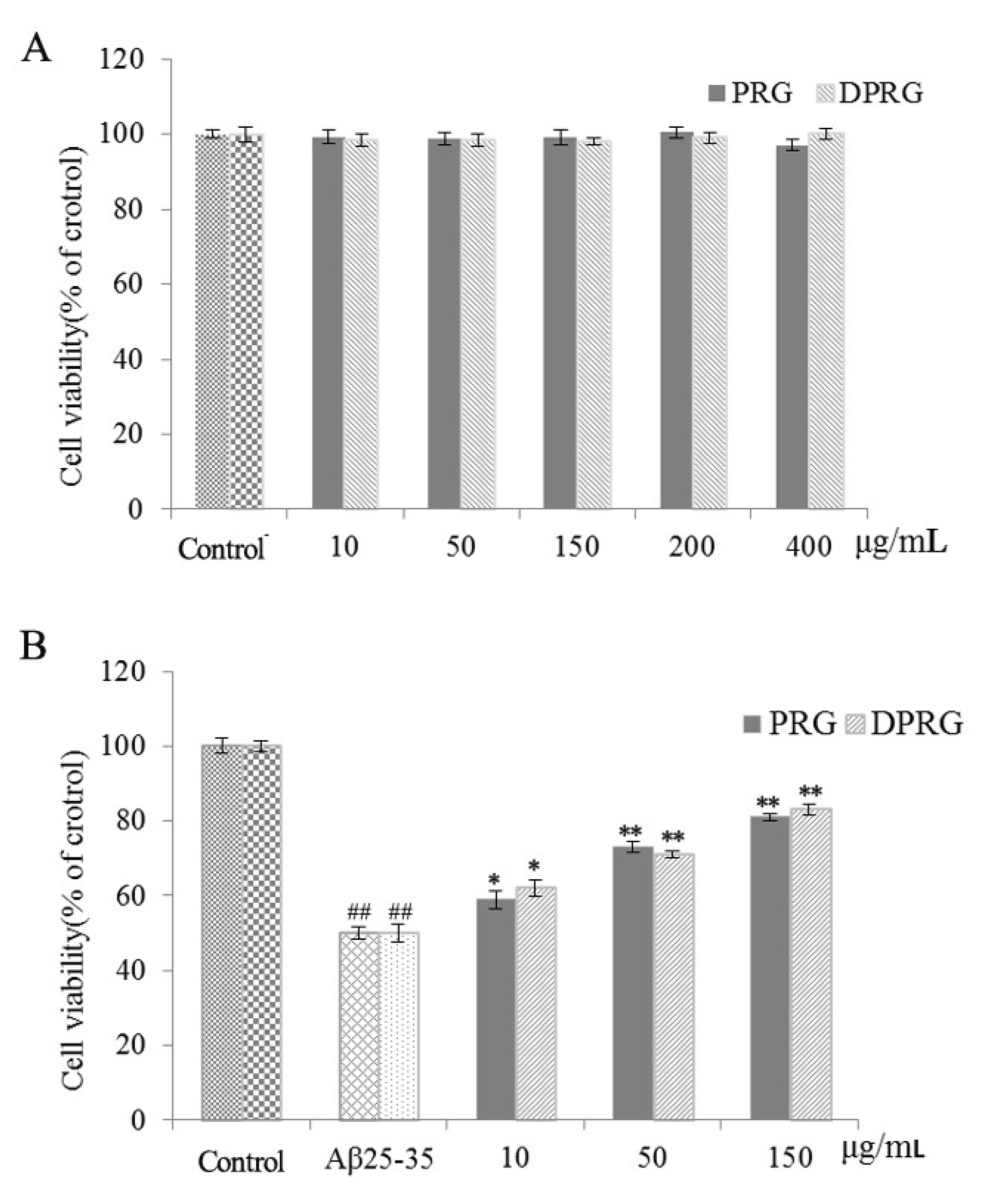
2.3. Effect of DPRG on Cell Viability in Aβ25–35-Damaged PC12 Cells
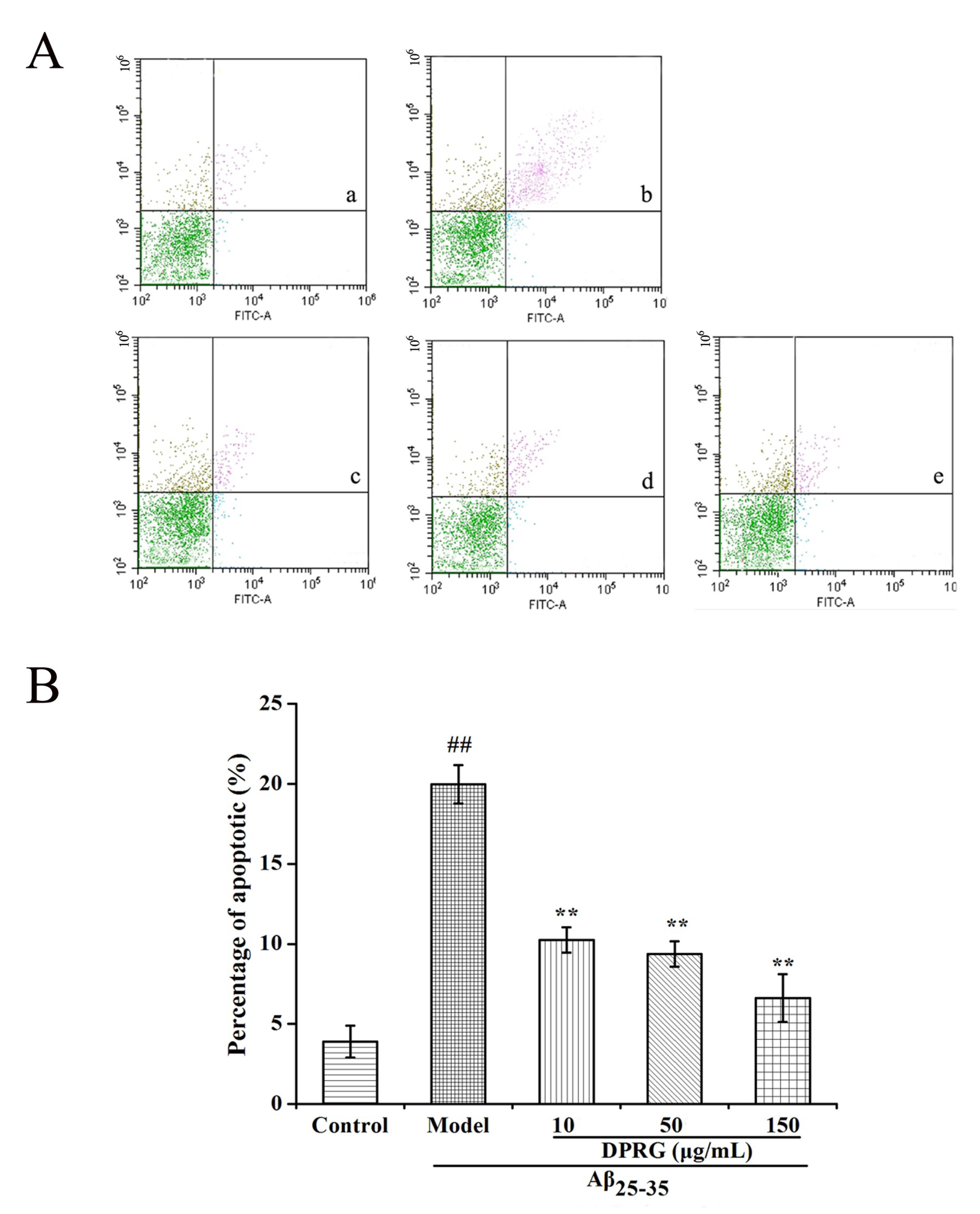
2.4. Effect of DPRG on Aβ25–35-Induced Apoptosis in PC 12 Cells
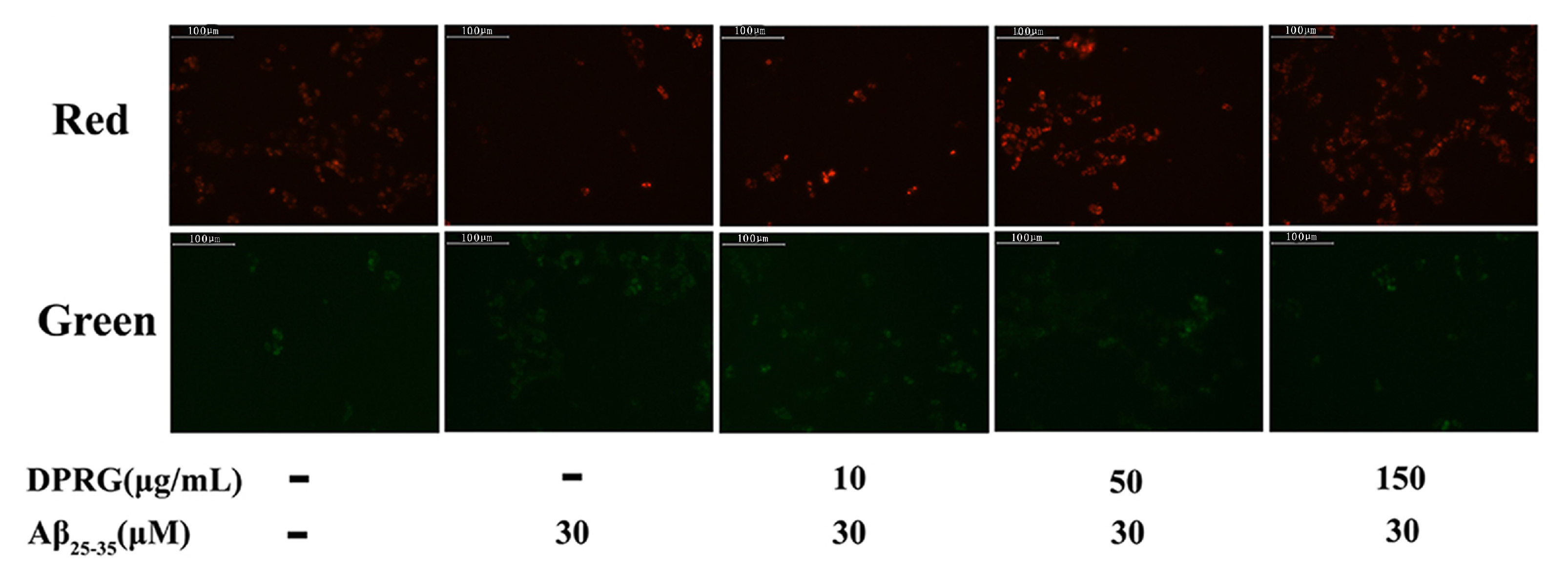
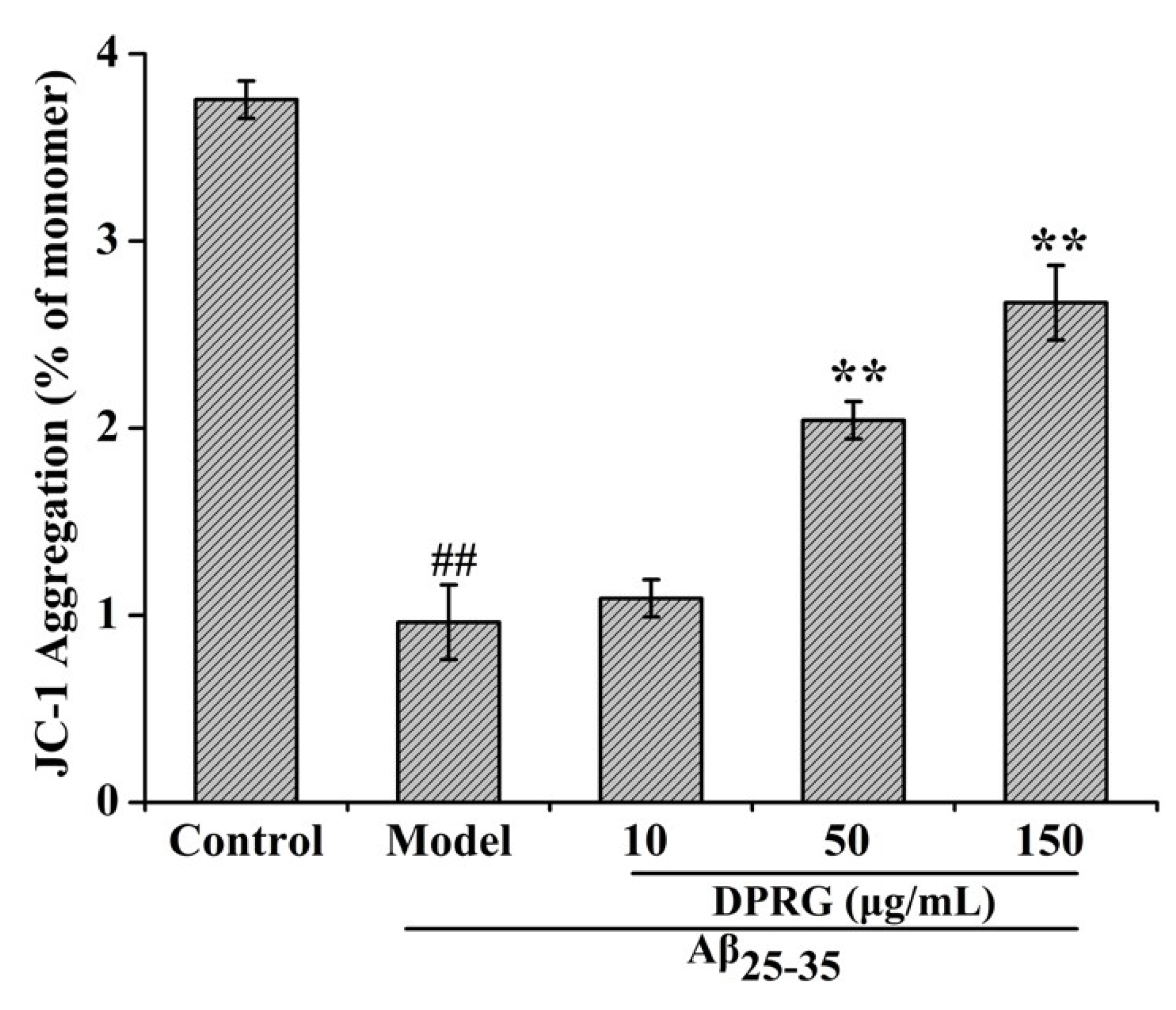
2.5. Effect of DPRG on Mitochondrial Dysfunction in Aβ25–35-treated PC12 Cells
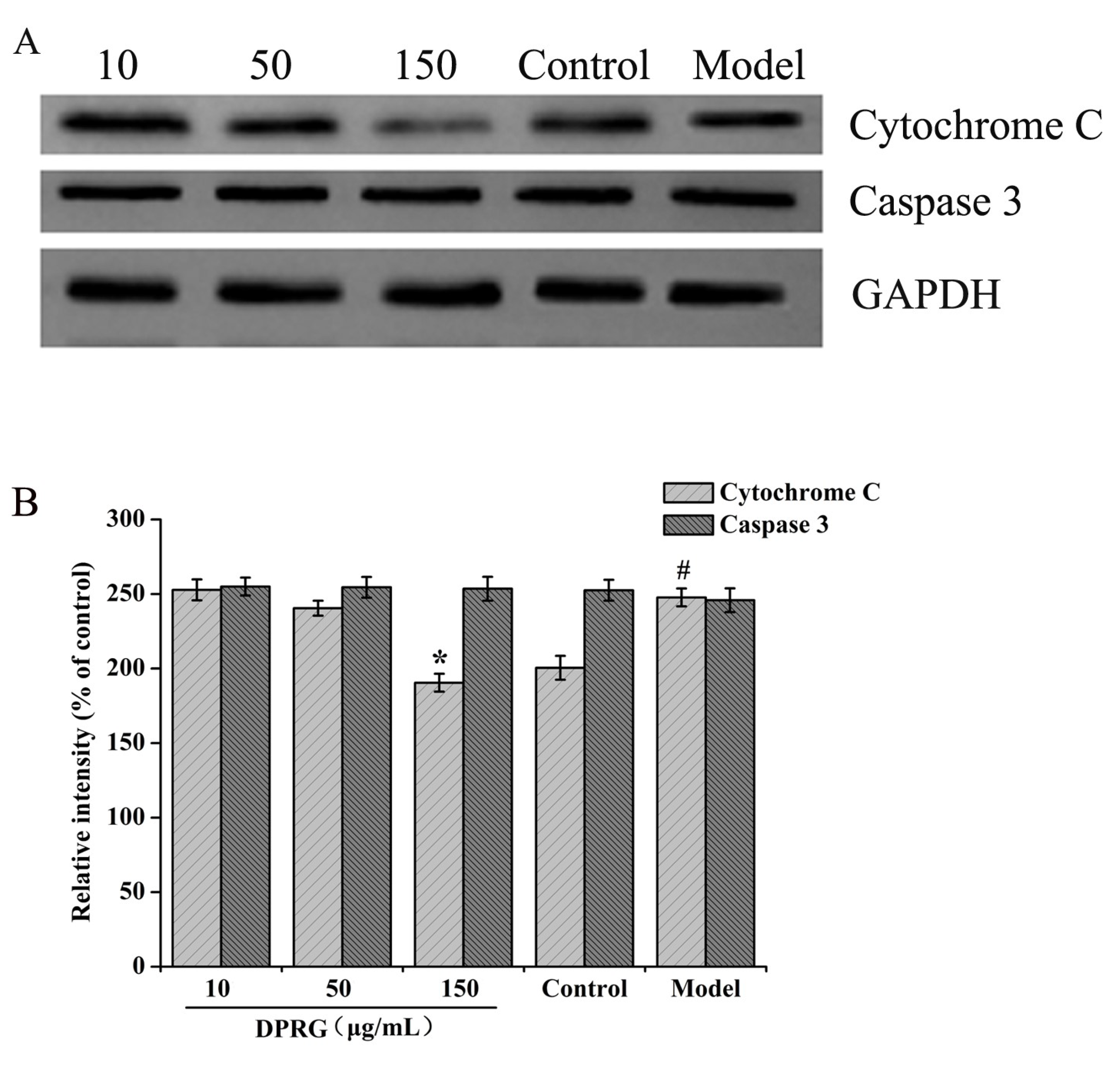
2.6. Effect of DPRG on Protein Expression in Aβ25–35-Treated PC12 Cells
3. Materials and Methods
3.1. Materials and Reagents
3.2. Preparation of PRG
3.3. Preparation of DPRG
3.4. Experimental Design of RSM
3.5. Molecular Weight Analysis
3.6. Characterization of DPRG
3.7. Cell Culture and Treatment
3.8. Preparation of Aβ25–35
3.9. Cell Viability Assay
3.10. Flow Cytometric Analysis
3.11. MMP Measurement
3.12. Western Blotting
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Diana, F.; Javier, D.; Lidia, A. A Study of Amyloid-β and Phosphotau in Plaques and Neurons in the Hippocampus of Alzheimer’s Disease Patients. J. Alzheimers Dis. 2018, 64, 417–435. [Google Scholar]
- Yang, S.; Zhang, R.; Wang, G.; Zhang, Y. The development prospection of HDAC inhibitors as a potential therapeutic direction in Alzheimer’s disease. Transl. Neurodegener. 2017, 6, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Mohammadhosseini, M.; Abadi, A.; Akbarzadeh, A.; Mellatyar, H. An Update on Biomedical Application of Nanotechnology for Alzheimer’s Disease Diagnosis and Therapy. Drug Res. 2016, 66, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, M.; Uchino, K.; Watanabe, Y.; Adachi, T.; Nakanishi, M.; Ichino, H.; Hongo, K.; Mizobata, T.; Kobayashi, S.; Nakashima, K.; et al. Anthocyanin suppresses the toxicity of Aβ deposits through diversion of molecular forms in in vitro and in vivo models of Alzheimer’s disease. Nutr. Neurosci. 2016, 19, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guan, S.; Liu, C.; Chen, X.; Zhu, Y.; Xie, Y.; Wang, J.; Ji, X.; Li, L.; Li, Z.; et al. Neuroprotective effects of Coptis chinensis Franch polysaccharide on amyloid-beta (Aβ)-induced toxicity in a transgenic Caenorhabditis elegans model of Alzheimer’s disease (AD). Int. J. Biol. Macromol. 2018, 113, 991–995. [Google Scholar] [CrossRef]
- Bag, S.; Chaudhury, S.; Pramanik, D.; DasGupta, S.; Dasgupta, S. Hydrophobic tail length plays a pivotal role in amyloid beta (25-35) fibril-surfactant interactions. Proteins 2016, 84, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Fresta, C.G.; Lazzarino, G.; Distefano, D.A.; Parlascino, P.; Lazzarino, G.; Caraci, F. Sub-Toxic Human Amylin Fragment Concentrations Promote the Survival and Proliferation of SH-SY5Y Cells via the Release of VEGF and HspB5 from Endothelial RBE4 Cells. Int. J. Mol. Sci. 2018, 19, 3659. [Google Scholar] [CrossRef]
- Zimbone, S.; Monaco, I.; Gianì, F.; Pandini, G.; Copani, A.G.; Giuffrida, M.L.; Rizzarelli, E. Amyloid Beta monomers regulate cyclic adenosine monophosphate response element binding protein functions by activating type-1 insulin-like growth factor receptors in neuronal cells. Aging Cell 2018, 17, e12684. [Google Scholar] [CrossRef]
- Caruso, G.; Distefano, D.A.; Parlascino, P.; Fresta, C.G.; Lazzarino, G.; Lunte, S.M.; Nicoletti, V.G. Receptor-mediated toxicity of human amylin fragment aggregated by short- and long-term incubations with copper ions. Mol. Cell. Biochem. 2017, 425, 85–93. [Google Scholar] [CrossRef]
- Ling, J. Radix Hedysari Polysaccharide Protects PC12 Cells against Aβ25–35-Induced Apoptosis via PRKCB/ERK-Dependent Pathways; Southern Medical University: GuangZhou, China, 2018. [Google Scholar]
- Chen, B.; Shi, M.; Cui, S.; Hao, S.; Hider, R.; Zhou, T. Improved antioxidant and anti-tyrosinase activity of polysaccharide from Sargassum fusiforme by degradation. Int. J. Biol. Macromol. 2016, 92, 715–722. [Google Scholar] [CrossRef]
- Lemieszek, M.; Nunes, F.; Cardoso, C.; Marques, G.; Rzeski, W. Neuroprotective properties of, Cantharellus cibarius, polysaccharide fractions in different, in vitro, models of neurodegeneration. Carbohydr. Polym. 2018, 197, 598–607. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, C.; Jiang, H.; Zhou, H.; Li, P.; Wang, F. Isolation, structural characterization and neurotrophic activity of a polysaccharide from Phellinus ribis. Carbohydr. Polym. 2015, 127, 145–151. [Google Scholar] [CrossRef]
- Zhao, X.; Li, B.; Xue, C.; Sun, L. Effect of molecular weight on the antioxidant property of low molecular weight alginate from Laminaria japonica. J. Appl. Phycol. 2012, 24, 295–300. [Google Scholar] [CrossRef]
- Zhao, Y.; Shi, Y.; Yang, H.; Mao, L. Extraction of Angelica sinensis polysaccharides using Ultrasound-assisted way and its bioactivity. Int. J. Biol. Macromol. 2016, 88, 44–50. [Google Scholar] [CrossRef]
- Pan, P.; Jin, W.; Li, X.; Chen, Y.; Jiang, J.; Wan, H.; Yu, D. Optimization of multiplex quantitative polymerase chain reaction based on response surface methodology and an artificial neural network-genetic algorithm approach. PLoS ONE 2018, 13, e0200962. [Google Scholar] [CrossRef]
- Gu, F.; Xu, F.; Tan, L.; Wu, H.; Chu, Z.; Wang, Q. Optimization of Enzymatic Process for Vanillin Extraction Using Response Surface Methodology. Molecules 2012, 17, 8753–8761. [Google Scholar] [CrossRef]
- Wang, R.; Chen, P.; Jia, F.; Tang, J.; Ma, F. Optimization of polysaccharides from Panax japonicus C.A. Meyer by RSM and its anti-oxidant activity. Int. J. Biol. Macromol. 2012, 50, 331–336. [Google Scholar] [CrossRef]
- González-Centeno, M.; Knoerzer, K.; Sabarez, H.; Simal, S.; Rosselló, C.; Femenia, A. Effect of acoustic frequency and power density on the aqueous ultrasonic-assisted extraction of grape pomace (Vitis vinifera L.)—A response surface approach. Ultrason Sonochem. 2014, 21, 2176–2184. [Google Scholar] [CrossRef]
- Mohammad, A.; Abdulhameed, A.; Jawad, A. Box-Behnken design to optimize the synthesis of new crosslinked chitosan-glyoxal/TiO2 nanocomposite: Methyl orange adsorption and mechanism studies. Int. J. Biol. Macromol. 2019, 129, 98–109. [Google Scholar] [CrossRef]
- Gupta, P.; Nayak, K. Optimization of keratin/alginate scaffold using RSM and its characterization for tissue engineering. Int. J. Biol. Macromol. 2016, 85, 141–149. [Google Scholar] [CrossRef]
- Maran, J.P.; Mekala, V.; Manikandan, S. Modeling and optimization of ultrasound-assisted extraction of polysaccharide from Cucurbita moschata. Carbohydr. Polym. 2013, 92, 2018–2026. [Google Scholar] [CrossRef]
- Gu, P.; Xu, S.; Zhou, S.; Liu, Z.; Sun, Y.; Ou, N.; Hu, Y.; Liu, J.; Wu, Y.; Wang, X.; et al. Optimization of angelica sinensis polysaccharide-loaded Poly (lactic-co-glycolicacid) nanoparticles by RSM and its immunological activity in vitro. Int. J. Biol. Macromol. 2018, 107, 222–229. [Google Scholar] [CrossRef]
- Kuo, C.; Hsiao, F.; Chen, J.; Hsieh, C.; Liu, Y.; Shieh, C. Kinetic aspects of ultrasound-accelerated lipase catalyzed acetylation and optimal synthesis of 4′-acetoxyresveratrol. Ultrason Sonochem. 2013, 20, 546–552. [Google Scholar] [CrossRef]
- Shan, H.; Chu, Y.; Chang, P.; Yang, L.; Wang, Y.; Zhu, S.; Zhang, M.; Tao, L. Neuroprotective effects of hydrogen sulfide on sodium azideinduced autophagic cell death in PC12 cells. Mol. Med. Rep. 2017, 16, 5938–5946. [Google Scholar] [CrossRef][Green Version]
- Sheng, Y.; Liu, G.; Wang, M.; Lv, Z.; Du, P. A selenium polysaccharide from, Platycodon grandiflorum, rescues PC12 cell death caused by H2O2, via inhibiting oxidative stress. Int. J. Biol. Macromol. 2017, 104, 393–399. [Google Scholar] [CrossRef]
- Hao, L.; Zhang, Q.; Yu, T.; Cheng, Y.; Ji, S. Antagonistic effects of ultra-low-molecular-weight heparin on Aβ25–35-induced apoptosis in cultured rat cortical neurons. Brain Res. 2011, 1368, 1–10. [Google Scholar] [CrossRef]
- Zhang, H.; Cao, Y.; Chen, L.; Wang, J.; Tian, Q.; Wang, N.; Liu, Z.; Li, J.; Wang, N.; Wang, X.; et al. A polysaccharide from Polygonatum sibiricum attenuates amyloid-β-induced neurotoxicity in PC12 cells. Carbohydr. Polym. 2015, 117, 879–886. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, J.; Liu, C.; Song, C.; Li, P.; Yin, F.; Xiao, Y.; Li, J.; Jiang, W.; Zong, A.; et al. Protective effects of low molecular weight chondroitin sulfate on amyloid beta (Aβ)-induced damage in vitro and in vivo. Neuroscience 2015, 305, 169–182. [Google Scholar] [CrossRef]
- Van Tonder, A.; Joubert, A.; Cromarty, A. Limitations of the 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyl-2H-tetrazolium bromide (MTT) assay when compared to three commonly used cell enumeration assays. BMC Res. Notes 2015, 8, 47–56. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Z.; Long, T.; Zhou, L.; Bao, Y. Immunomodulatory effects of herbal formula of astragalus polysaccharide (APS) and polysaccharopeptide (PSP) in mice with lung cancer. Int. J. Biol. Macromol. 2018, 106, 596–601. [Google Scholar] [CrossRef]
- Zapotocky, M.; Mejstrikova, E.; Smetana, K.; Stary, J.; Trka, J.; Starkova, J. Valproic acid triggers differentiation and apoptosis in AML1/ETO-positive leukemic cells specifically. Cancer Lett. 2012, 319, 144–153. [Google Scholar] [CrossRef]
- Menges, S.; Minakaki, G.; Schaefer, P.; Meixner, H.; Prots, I.; Schlötzer-Schrehardt, U.; Friedland, K.; Winner, B.; Outeiro, T.; Winklhofer, K.; et al. Alpha-synuclein prevents the formation of spherical mitochondria and apoptosis under oxidative stress. Sci. Rep. 2017, 7, 42942–42967. [Google Scholar] [CrossRef]
- Cheng, W.; Chen, W.; Wang, P. Asiatic acid protects differentiated PC12 cell from Aβ-induced apoptosis and tau hyperphosphorylation via regulating PI3K/Akt/GSK-3β signaling. Life Sci. 2018, 208, 96–101. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, T.; Wang, T.; Wang, B. Suppression of lncRNA-ATB prevents amyloid-β-induced neurotoxicity in PC12 cells via regulating miR-200/ZNF217 axis. Biomed. Pharmacother. 2018, 108, 707–715. [Google Scholar] [CrossRef]
- Hackenbeck, T.; Huber, R.; Schietke, R.; Knaup, K.; Monti, J.; Wu, X.; Klanke, B.; Frey, B.; Gaipl, U.; Wullich, B.; et al. The GTPase RAB20 is a HIF target with mitochondrial localization mediating apoptosis in hypoxia. Biochim. Biophys. Acta 2011, 1813, 1–13. [Google Scholar] [CrossRef]
- Xiong, S.; Mu, T.; Wang, G.; Jiang, X. Mitochondria-mediated apoptosis in mammals. Protein Cell 2014, 5, 737–749. [Google Scholar] [CrossRef]
- Wang, L.; Duan, Y.; Ma, Y.; Ding, H.; Li, E. Studies on extraction and antioxidant function of polysaccharides from Sophora japonica. J. Northwest A F Nat. Sci. Ed. 2008, 36, 213–218. [Google Scholar]
- Ru, Y.; Chen, X.; Xu, J.; Huang, L.; Jiang, M.; Guo, L.; Lin, Z.; Qiu, B.; Wong, K. Hypoglycemic effects of a polysaccharide from Tetrastigma hemsleyanum Diels & Gilg in alloxan-induced diabetic mice. Chem. Biodivers. 2018, 15, e1800070. [Google Scholar]
- Fujii, M.; Sato, Y.; Ito, H.; Masago, Y.; Omura, T. Monosaccharide composition of the outer membrane lipopolysaccharide and O-chain from the freshwater cyanobacterium Microcystis aeruginosa NIES-87. J. Appl. Microbiol. 2012, 113, 896–903. [Google Scholar] [CrossRef]
- Cao, C.; Huang, Q.; Zhang, B.; Li, C.; Fu, X. Physicochemical characterization and in vitro hypoglycemic activities of polysaccharides from Sargassum pallidum by microwave-assisted aqueous two-phase extraction. Int. J. Biol. Macromol. 2018, 109, 357–368. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, L.; Weng, Q.; Xiao, L.; Li, Q. Curcumin analogues attenuate Aβ-induced oxidative stress in PC12 cells via Keap1/Nrf2/HO-1 signaling pathways. Chem. Biol. Interact. 2019, 305, 171–179. [Google Scholar] [CrossRef]
- Deng, Q.; Yang, X. Protective effects of Gynostemma pentaphyllum polysaccharides on PC12 cells impaired by MPP+. Int. J. Biol. Macromol. 2014, 69, 171–175. [Google Scholar] [CrossRef]
- Lee, S.; Youn, K.; Jeong, W.; Ho, C.; Jun, M. Protective Effects of Red Ginseng Oil against Aβ25–35-Induced Neuronal Apoptosis and Inflammation in PC12 Cells. Int. J. Mol. Sci. 2017, 18, 2218–2237. [Google Scholar] [CrossRef]
- Silakhori, S.; Hosseinzadeh, H.; Behbahani, F.S.; Mehri, S. Neuroprotective effect of clavulanic acid on trimethyltin (TMT)-induced cytotoxicity in PC12 cells. Drug Chem. Toxicol. 2019, 42, 187–193. [Google Scholar] [CrossRef]
Sample Availability: Not available. |









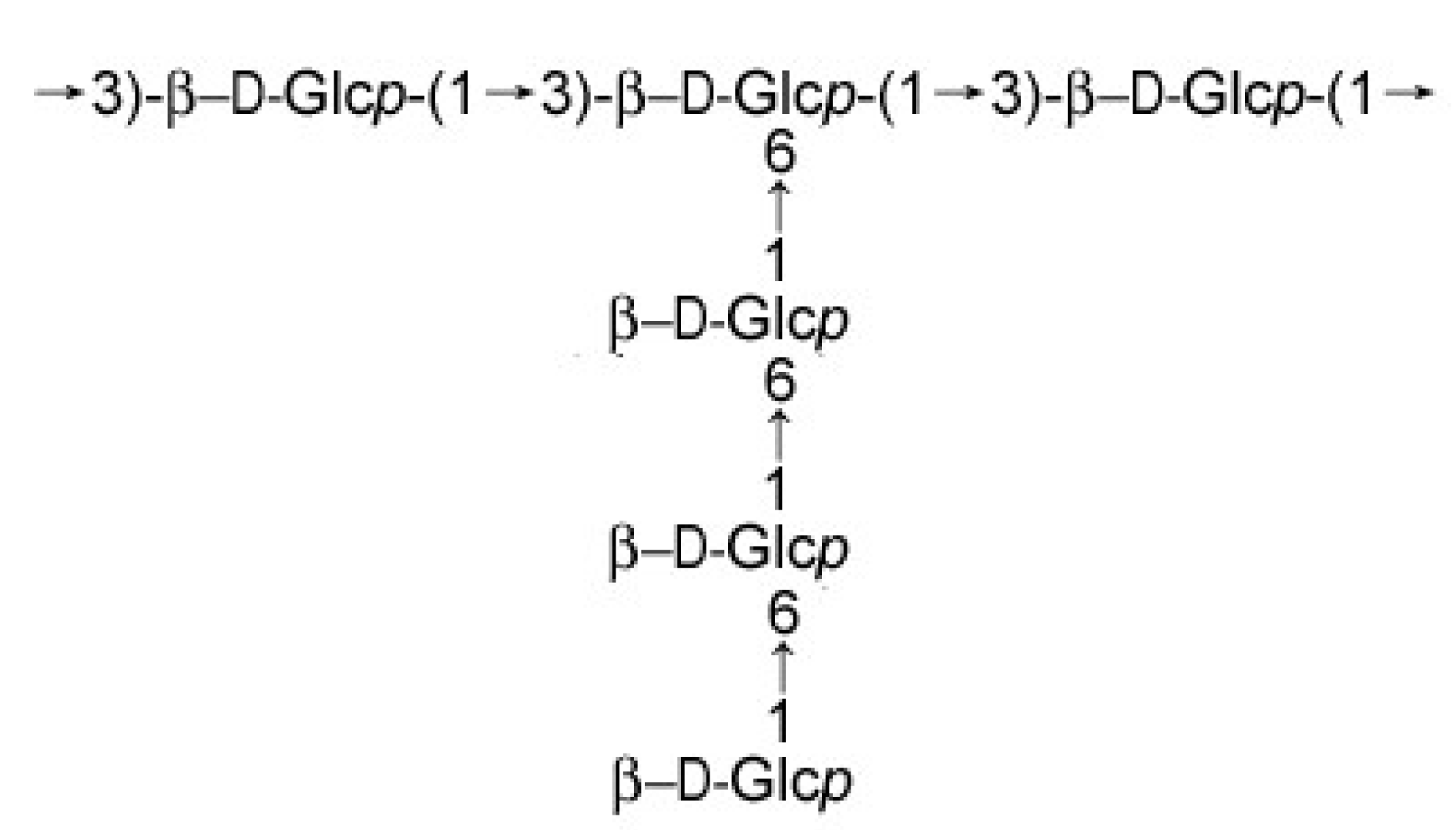{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | A | B | C | Y |
|---|---|---|---|---|
| 1 | 0 | −1 | 1 | 74.8 |
| 2 | 0 | 1 | 1 | 77.0 |
| 3 | 0 | −1 | −1 | 73.3 |
| 4 | 1 | 0 | −1 | 77.4 |
| 5 | −1 | −1 | 0 | 70.3 |
| 6 | −1 | 0 | 1 | 72.9 |
| 7 | −1 | 1 | 0 | 72.8 |
| 8 | 1 | 0 | 1 | 79.3 |
| 9 | 1 | −1 | 0 | 77.1 |
| 10 | 0 | 1 | −1 | 74.8 |
| 11 | 1 | 1 | 0 | 78.6 |
| 12 | −1 | 0 | −1 | 71.6 |
| 13 | 0 | 0 | 0 | 83.1 |
| 14 | 0 | 0 | 0 | 82.9 |
| 15 | 0 | 0 | 0 | 82.5 |
| 16 | 0 | 0 | 0 | 82.8 |
| 17 | 0 | 0 | 0 | 82.4 |
| Variables | Coded Symbols | Coded Levels | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| Concentration (mM) | A | 10 | 15 | 20 |
| Temperature (°C) | B | 40 | 50 | 60 |
| Time (h) | C | 1 | 1.5 | 2 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value Prob > F | |
|---|---|---|---|---|---|---|
| Model | 304.16 | 9 | 33.80 | 617.18 | <0.0001 | Significant |
| A | 75.89 | 1 | 75.89 | 1385.96 | <0.0001 | *** |
| B | 7.37 | 1 | 7.37 | 134.65 | <0.0001 | *** |
| C | 5.81 | 1 | 5.81 | 106.18 | <0.0001 | *** |
| AB | 0.26 | 1 | 0.26 | 4.66 | 0.0678 | |
| AC | 0.13 | 1 | 0.13 | 2.30 | 0.1730 | |
| BC | 0.12 | 1 | 0.12 | 2.17 | 0.1839 | |
| A2 | 63.67 | 1 | 63.67 | 1162.83 | <0.0001 | *** |
| B2 | 74.05 | 1 | 74.05 | 1352.39 | <0.0001 | *** |
| C2 | 54.38 | 1 | 54.38 | 993.10 | <0.0001 | *** |
| Residual | 0.38 | 7 | 0.055 | |||
| Lack of Fit | 0.057 | 3 | 0.019 | 0.23 | 0.8704 | not significant |
| Pure Error | 0.33 | 4 | 0.082 | |||
| Cor Total | 304.54 | 16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Jin, J.; Liu, Q.; Ma, D.; Li, J.; Zhang, Y.; Liu, Y. Optimization of Degradation Conditions with PRG, a Polysaccharide from Phellinus ribis, by RSM and the Neuroprotective Activity in PC12 Cells Damaged by Aβ25–35. Molecules 2019, 24, 3010. https://doi.org/10.3390/molecules24163010
Yang P, Jin J, Liu Q, Ma D, Li J, Zhang Y, Liu Y. Optimization of Degradation Conditions with PRG, a Polysaccharide from Phellinus ribis, by RSM and the Neuroprotective Activity in PC12 Cells Damaged by Aβ25–35. Molecules. 2019; 24(16):3010. https://doi.org/10.3390/molecules24163010
Chicago/Turabian StyleYang, Pei, Juan Jin, Qian Liu, Dongmei Ma, Jia Li, Yongqing Zhang, and Yuhong Liu. 2019. "Optimization of Degradation Conditions with PRG, a Polysaccharide from Phellinus ribis, by RSM and the Neuroprotective Activity in PC12 Cells Damaged by Aβ25–35" Molecules 24, no. 16: 3010. https://doi.org/10.3390/molecules24163010
APA StyleYang, P., Jin, J., Liu, Q., Ma, D., Li, J., Zhang, Y., & Liu, Y. (2019). Optimization of Degradation Conditions with PRG, a Polysaccharide from Phellinus ribis, by RSM and the Neuroprotective Activity in PC12 Cells Damaged by Aβ25–35. Molecules, 24(16), 3010. https://doi.org/10.3390/molecules24163010