β-Peltoboykinolic Acid from Astilbe rubra Attenuates TGF-β1-Induced Epithelial-to-Mesenchymal Transitions in Lung Alveolar Epithelial Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
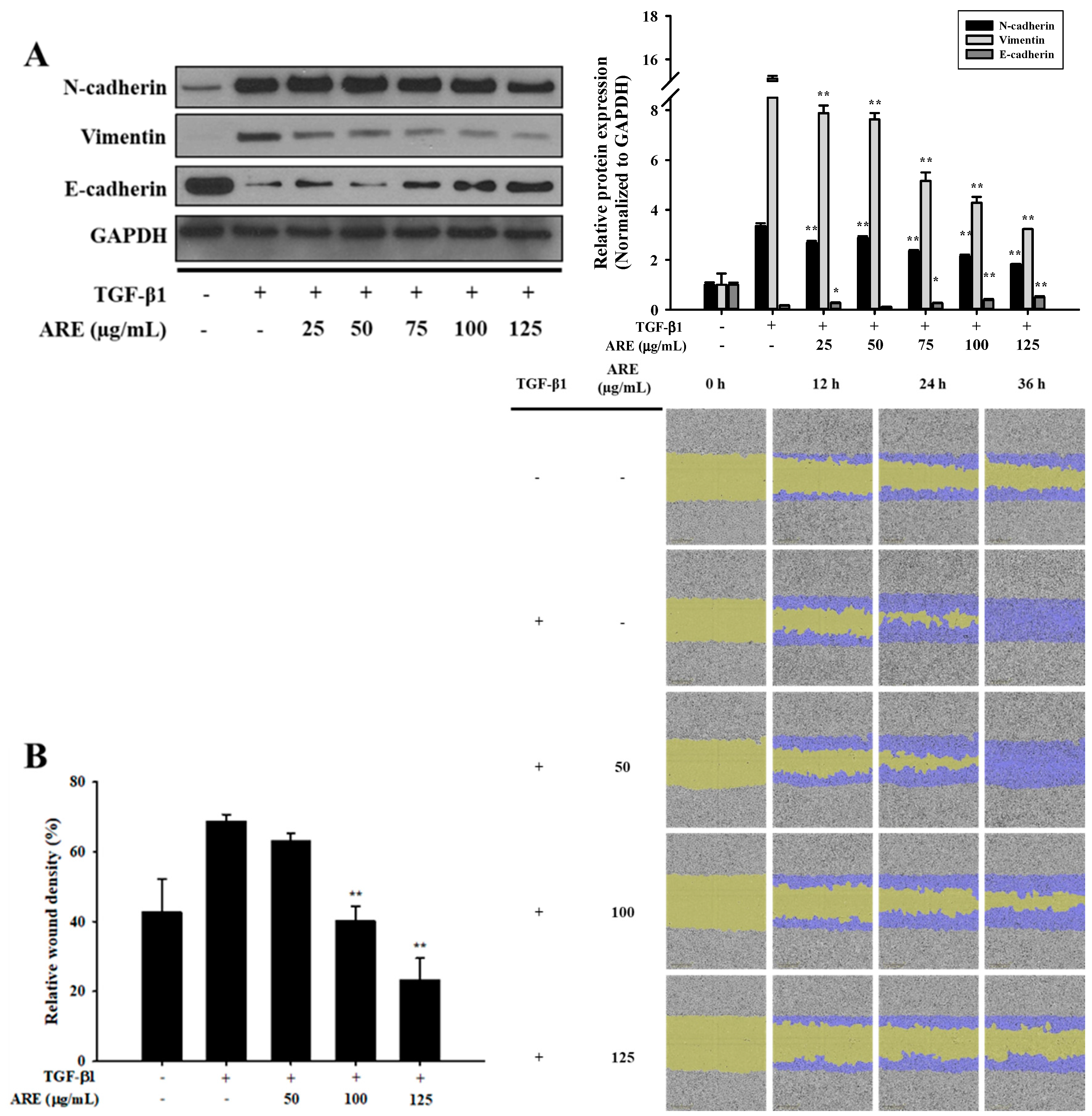
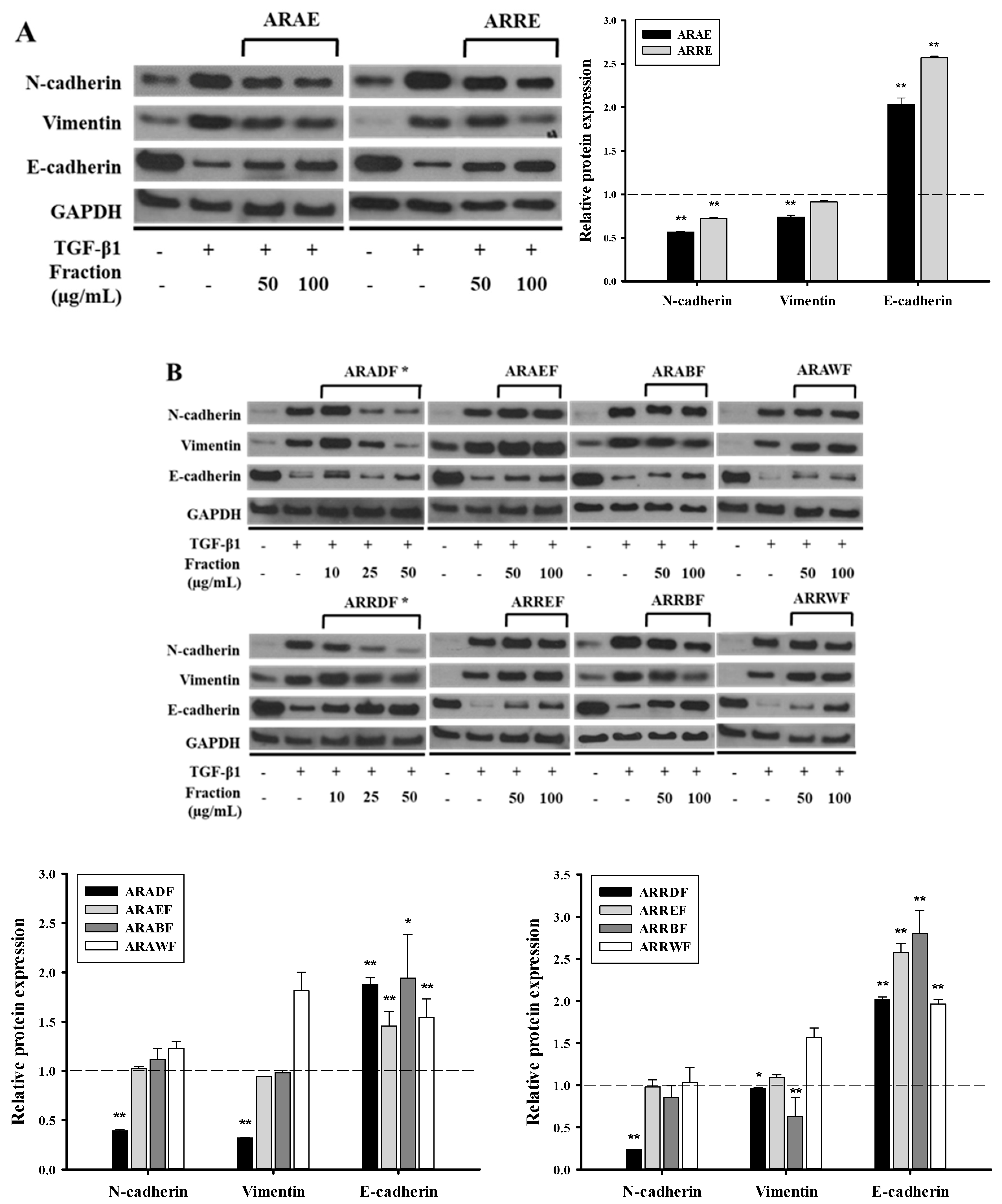
2.1. Ethanol Extract of A. rubra Whole Plant Inhibits TGF-β1-Induced EMT in A549 Cells
2.2. Dichloromethane Fractions Show the Strongest Inhibitory Effect on TGF-β1-Induced EMT
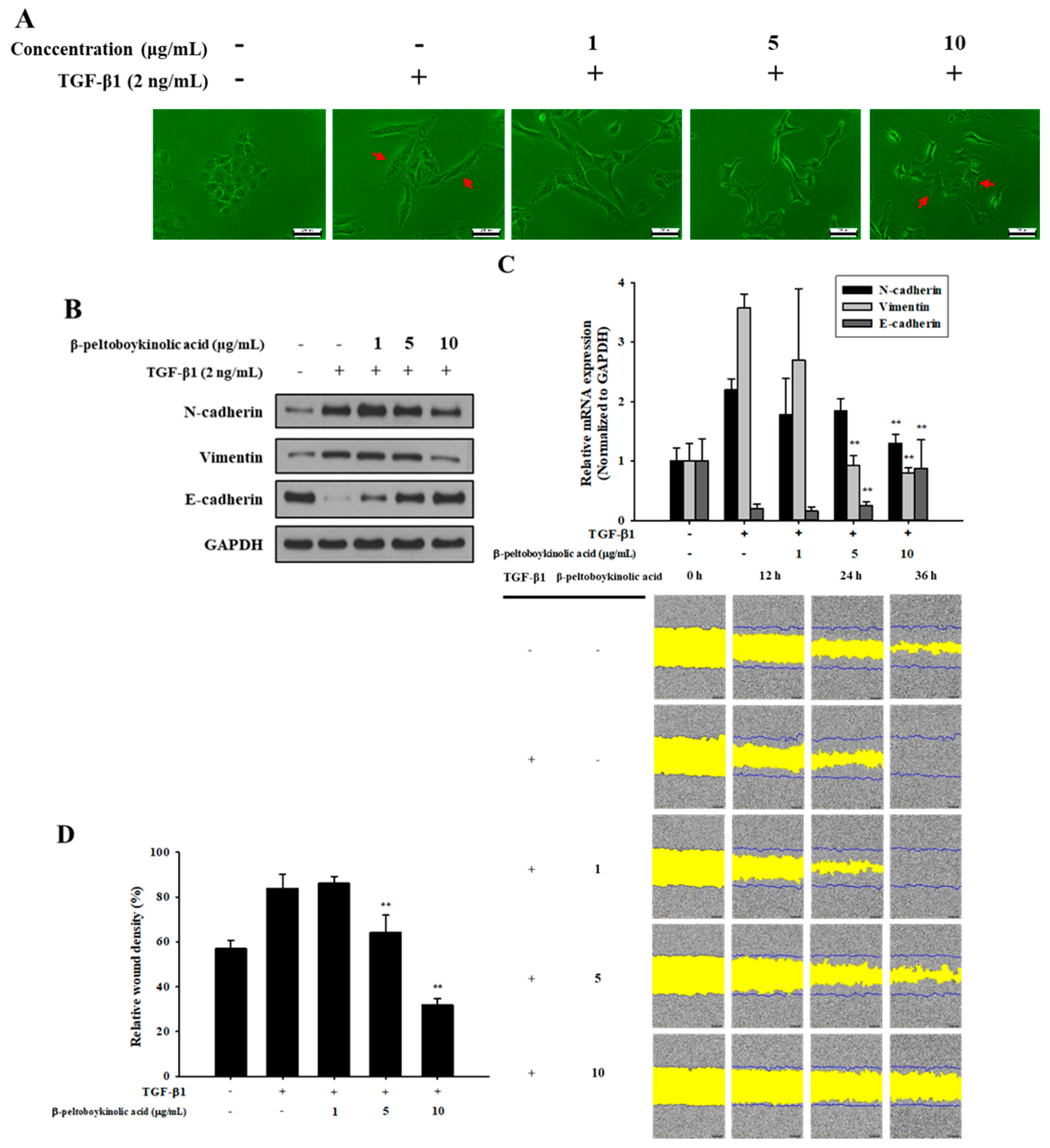
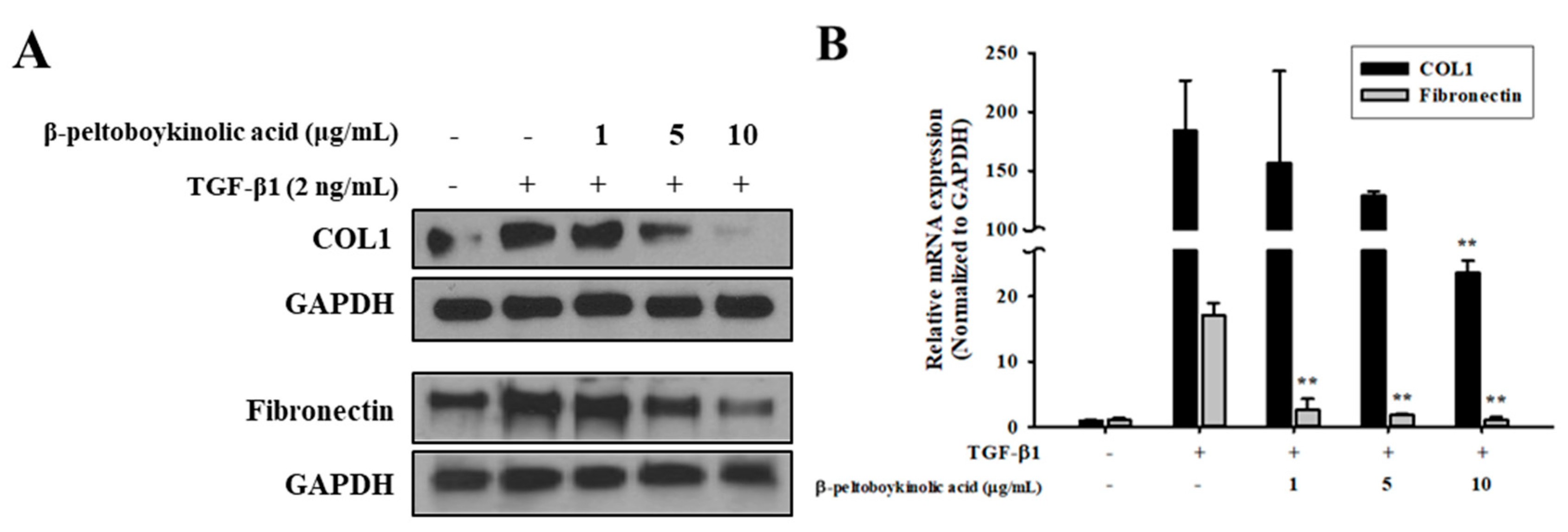
2.3. β-Peltoboykinolic Acid Attenuates TGF-β1-Induced EMT
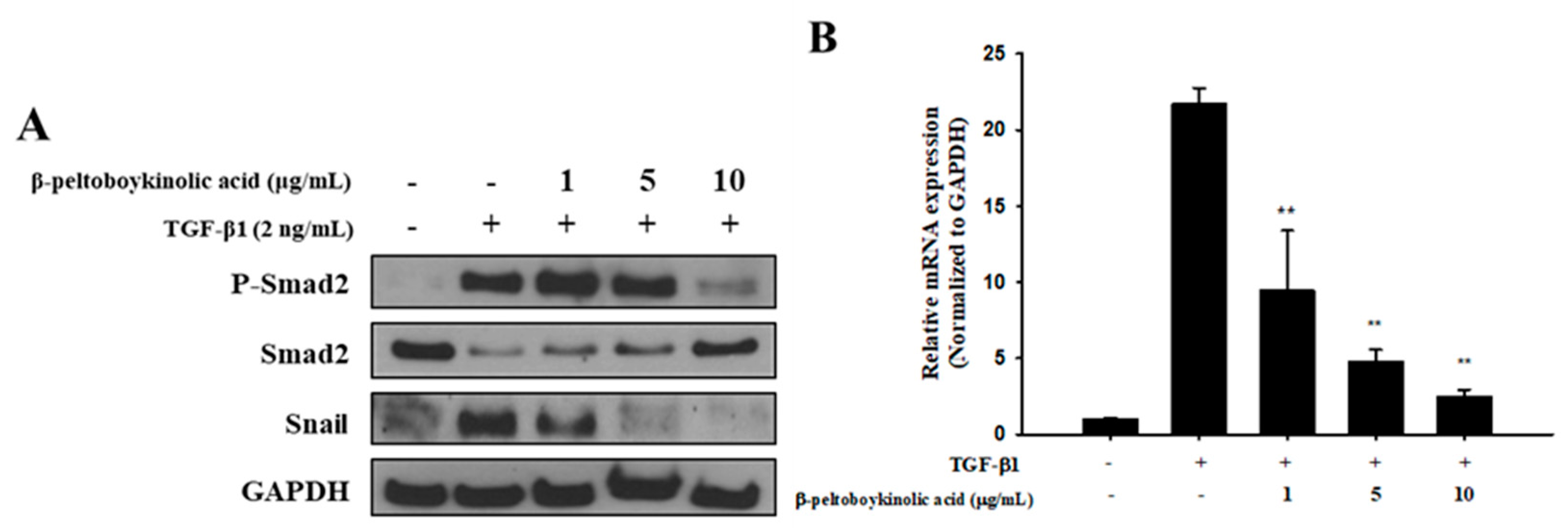
2.4. β-Peltoboykinolic Acid Interrupts the Activation of the Smad Pathway by TGF-β1
3. Discussion
4. Materials and Methods
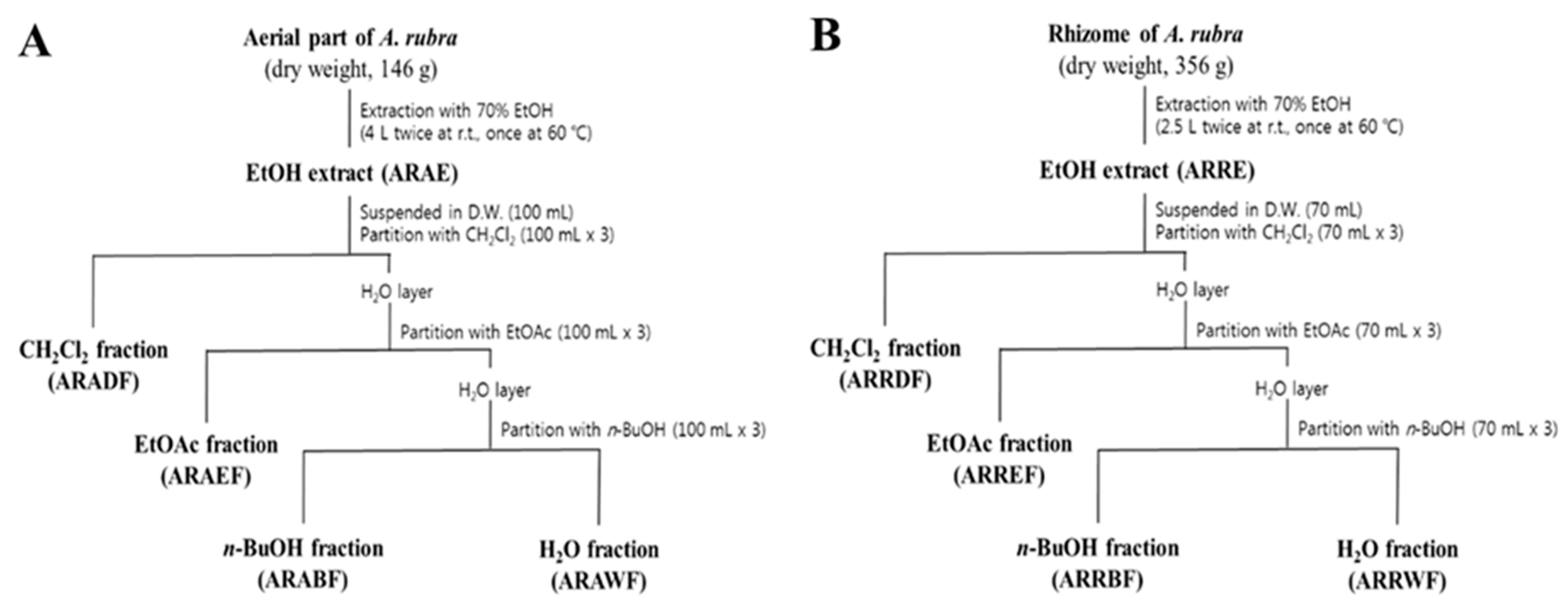
4.1. Plant Material
4.2. Extraction and Fractionation
4.3. Isolation of β-Peltoboykinolic Acid
4.4. Cell Culture and Reagents
4.5. Western Blot Analysis
4.6. COL1 Western Blot Analysis
4.7. Migration Assay
4.8. Quantitative Real-Time PCR (qRT-PCR)
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wells, A.U.; Rosas, I.O. Broad therapeutic efficacy of nintedanib in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care. Med. 2016, 193, 112–113. [Google Scholar] [CrossRef]
- King, T.E., Jr.; Pardo, A.; Selman, M. Idiopathic pulmonary fibrosis. Lancet 2011, 378, 1949–1961. [Google Scholar] [CrossRef]
- Chang, C.C.; Tsai, M.L.; Huang, H.C.; Chen, C.Y.; Dai, S.X. Epithelial-mesenchymal transition contributes to SWCNT-induced pulmonary fibrosis. Nanotoxicology 2012, 6, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Selman, M.; Pardo, A. Alveolar epithelial cell disintegrity and subsequent activation: A key process in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2012, 186, 119–121. [Google Scholar] [CrossRef]
- Bartis, D.; Mise, N.; Mahida, R.Y.; Eickelberg, O.; Thickett, D.R. Epithelial-mesenchymal transition in lung development and disease: Does it exist and is it important? Thorax 2014, 69, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Willis, B.C.; Borok, Z. TGF-beta-induced EMT: Mechanisms and implications for fibrotic lung disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L525–L534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.; Lai, M. Modulation of epithelial-to-mesenchymal cancerous transition by natural products. Fitoterapia 2015, 106, 247–255. [Google Scholar] [CrossRef]
- Kim, T.J. Korean Resources Plants, Vol. II; Seoul National University Press: Seoul, Korea, 1996; p. 82. [Google Scholar]
- Kim, H.Y.; Lim, S.H.; Park, Y.H.; Ham, H.J.; Lee, K.J.; Park, D.S.; Kim, K.H.; Kim, S.M. Screening of α-amylase, α-glucosidase and lipase inhibitory activity with Gangwon-do wild plants extracts. J. Korean Soc. Food Sci. Nutr. 2011, 40, 308–315. [Google Scholar] [CrossRef]
- Yang, Y.J.; Kim, H.J.; Kang, S.H.; Kang, S.C. Screening of natural herb resources for antioxidative effects in Korea. Korean J. Plant Res. 2011, 24, 1–9. [Google Scholar] [CrossRef][Green Version]
- Hong, J.Y.; Pakr, H.R.; Kwon, Y.G.; Kwon, W.Y.; Kim, S.Y.; Park, R.D.; Shin, S.Y. Anticancer properties of Astilbe rubra extracts on AsPC-1 human pancreatic cancer cells. J. Basic Sci. 2015, 32, 21–28. [Google Scholar]
- Sun, H.; Zhang, J.; Ye, Y.; Pan, Y.; Shen, Y. Cytotoxic pentacyclic triterpenoids from the rhizome of Astilbe chinensis. Helv. Chim. Acta 2003, 86, 2414–2422. [Google Scholar] [CrossRef]
- Raghu, G.; Rochwerg, B.; Zhang, Y.; Garcia, C.A.; Azuma, A.; Behr, J.; Brozek, J.L.; Collard, H.R.; Cunningham, W.; Homma, S.; et al. An Official ATS/ERS/JRS/ALAT Clinical Practice Guideline: Treatment of Idiopathic Pulmonary Fibrosis. An Update of the 2011 Clinical Practice Guideline. Am. J. Respir. Crit. Care Med. 2015, 192, e3–e19. [Google Scholar] [PubMed]
- Yang, Z.C.; Yi, M.J.; Ran, N.; Jin, L.Y.; Fu, P.; Feng, X.Y.; Lei, X.; Qu, Z.H. Astragalus extract inhibits TGF-β1-induced EMT of bronchial epithelial cells and airway remodeling in asthmatic mice. Int. J. Clin. Exp. Med. 2016, 9, 1281–1289. [Google Scholar]
- Li, R.; Wang, Y.; Liu, Y.; Chen, Q.; Fu, W.; Wang, H.; Cai, H.; Peng, W.; Zhang, X. Curcumin inhibits transforming growth factor-β1-induced EMT via PPARγ pathway, not Smad pathway in renal tubular epithelial cells. PLoS ONE 2013, 8, e58848. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Liu, X.; Han, Z.; Zhou, L.; Sui, H.; Yan, L.; Jiang, H.; Ren, J.; Cai, J.; Li, Q. Resveratrol suppresses epithelial-to-mesenchymal transition in colorectal cancer through TGF-β1/Smads signaling pathway mediated Snail/E-cadherin expression. BMC Cancer 2015, 15, 97. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Ko, J.H.; Lee, J.H.; Kim, C.; Lee, H.; Nam, D.; Lee, J.; Lee, S.G.; Yang, W.M.; Um, J.Y.; et al. Ginkgolic acid inhibits invasion and migration and TGF-β-induced EMT of lung cancer cells through PI3K/Akt/mTOR inactivation. J. Cell Physiol. 2017, 232, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Pattarayan, D.; Sivanantham, A.; Krishnaswami, V.; Loganathan, L.; Palanichamy, R.; Natesan, S.; Muthusamy, K.; Rajasekaran, S. Tannic acid attenuates TGF-β1-induced epithelial-to-mesenchymal transition by effectively intervening TGF-β signaling in lung epithelial cells. J. Cell. Physiol. 2018, 233, 2513–2525. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Lee, I.; Chang, M.J.; Yoo, J.K.; Na, M.; Hung, T.M.; Thuong, P.T.; Lee, J.; Kim, J.H.; Kim, J.C.; et al. Anticomplementary activity of triterpenoids from the whole plant of Aceriphyllum rossii against the classical pathway. Planta Med. 2008, 74, 726–729. [Google Scholar] [CrossRef]
- Arisawa, M.; Bai, H.; Shimizu, S.; Koshimura, S.; Tanaka, M.; Sasaki, T.; Morita, N. Isolation and identification of a cytotoxic principle from Chrysosplenium grayanum Maxim. (Saxifragaceae) and its antitumor activities. Chem. Pharm. Bull. (Tokyo). 1992, 40, 3274–3276. [Google Scholar] [CrossRef][Green Version]
- Na, M.; Cui, L.; Min, B.S.; Bae, K.; Yoo, J.K.; Kim, B.Y.; Oh, W.K.; Ahn, J.S. Protein tyrosine phosphatase 1B inhibitory activity of triterpenes isolated from Astilbe koreana. Bioorg. Med. Chem. Lett. 2006, 16, 3273–3276. [Google Scholar] [CrossRef]
- Lee, I.; Yoo, J.K.; Na, M.; Min, B.S.; Lee, J.; Yun, B.S.; Jin, W.; Kim, H.; Youn, U.; Chen, Q.C.; et al. Cytotoxicity of triterpenes isolated from Aceriphyllum rossii. Chem. Pharm. Bull. (Tokyo) 2007, 55, 1376–1378. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhao, J.; Shi, W.; Wang, Y.L.; Chen, H.; Bringas, P., Jr.; Datto, M.B.; Frederick, J.P.; Wang, X.F.; Warburton, D. Smad3 deficiency attenuates bleomycin-induced pulmonary fibrosis in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, L585–L593. [Google Scholar] [CrossRef] [PubMed]
- Raghow, R. The role of extracellular matrix in postinflammatory wound healing and fibrosis. FASEB J. 1994, 8, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Imamichi, Y.; König, A.; Gress, T.; Menke, A. Collagen type I-induced Smad-interacting protein 1 expression downregulates E-cadherin in pancreatic cancer. Oncogene 2007, 26, 2381–2385. [Google Scholar] [CrossRef] [PubMed]
- Muro, A.F.; Moretti, F.A.; Moore, B.B.; Yan, M.; Atrasz, R.G.; Wilke, C.A.; Flaherty, K.R.; Martinez, F.J.; Tsui, J.L.; Sheppard, D.; et al. An essential role for fibronectin extra type III domain A in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2008, 177, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.J.; Yuan, W.; Mori, Y.; Levenson, A.; Trojanowska, M.; Varga, J. Stimulation of type I collagen transcription in human skin fibroblasts by TGF-beta: Involvement of Smad 3. J. Investig. Dermatol. 1999, 112, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Isono, M.; Chen, S.; Hong, S.W.; Iglesias-de la Cruz, M.C.; Ziyadeh, F.N. Smad pathway is activated in the diabetic mouse kidney and Smad3 mediates TGF-beta-induced fibronectin in mesangial cells. Biochem. Biophys. Res. Commun. 2002, 296, 1356–1365. [Google Scholar] [CrossRef]
- Batlle, E.; Sancho, E.; Francí, C.; Domínguez, D.; Monfar, M.; Baulida, J.; García De Herreros, A. The transcription factor snail is a repressor of E-cadherin gene expression in epithelial tumour cells. Nat. Cell Biol. 2000, 2, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Walton, K.L.; Johnson, K.E.; Harrison, C.A. Targeting TGF-β Mediated SMAD Signaling for the Prevention of Fibrosis. Front. Pharmacol. 2017, 8, 461. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Q.; Feng, Y.L.; Cao, G.; Zhao, Y.Y. Natural Products as a Source for Antifibrosis Therapy. Trends Pharmacol. Sci. 2018, 39, 937–952. [Google Scholar] [CrossRef]
- Yoon, J.H.; Sudo, K.; Kuroda, M.; Kato, M.; Lee, I.K.; Han, J.S.; Nakae, S.; Imamura, T.; Kim, J.; Ju, J.H.; et al. Phosphorylation status determines the opposing functions of Smad2/Smad3 as STAT3 cofactors in TH17 differentiation. Nat. Commun. 2015, 6, 7600. [Google Scholar] [CrossRef] [PubMed]
- Ooshima, A.; Park, J.; Kim, S.J. Phosphorylation status at Smad3 linker region modulates transforming growth factor-β-induced epithelial-mesenchymal transition and cancer progression. Cancer Sci. 2019, 110, 481. [Google Scholar] [CrossRef] [PubMed]
- Nacif, M.; Shaker, O. Targeting transforming growth factor-β (TGF-β) in cancer and non-neoplastic diseases. J. Cancer Ther. 2014, 5, 735–747. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bang, I.J.; Kim, H.R.; Jeon, Y.; Jeong, M.H.; Park, Y.J.; Kwak, J.H.; Chung, K.H. β-Peltoboykinolic Acid from Astilbe rubra Attenuates TGF-β1-Induced Epithelial-to-Mesenchymal Transitions in Lung Alveolar Epithelial Cells. Molecules 2019, 24, 2573. https://doi.org/10.3390/molecules24142573
Bang IJ, Kim HR, Jeon Y, Jeong MH, Park YJ, Kwak JH, Chung KH. β-Peltoboykinolic Acid from Astilbe rubra Attenuates TGF-β1-Induced Epithelial-to-Mesenchymal Transitions in Lung Alveolar Epithelial Cells. Molecules. 2019; 24(14):2573. https://doi.org/10.3390/molecules24142573
Chicago/Turabian StyleBang, In Jae, Ha Ryong Kim, Yukyoung Jeon, Mi Ho Jeong, Yong Joo Park, Jong Hwan Kwak, and Kyu Hyuck Chung. 2019. "β-Peltoboykinolic Acid from Astilbe rubra Attenuates TGF-β1-Induced Epithelial-to-Mesenchymal Transitions in Lung Alveolar Epithelial Cells" Molecules 24, no. 14: 2573. https://doi.org/10.3390/molecules24142573
APA StyleBang, I. J., Kim, H. R., Jeon, Y., Jeong, M. H., Park, Y. J., Kwak, J. H., & Chung, K. H. (2019). β-Peltoboykinolic Acid from Astilbe rubra Attenuates TGF-β1-Induced Epithelial-to-Mesenchymal Transitions in Lung Alveolar Epithelial Cells. Molecules, 24(14), 2573. https://doi.org/10.3390/molecules24142573