
The Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′- dimethylchalcone from Cleistocalyx operculatus Buds on Human Pancreatic Cancer Cell Lines


Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
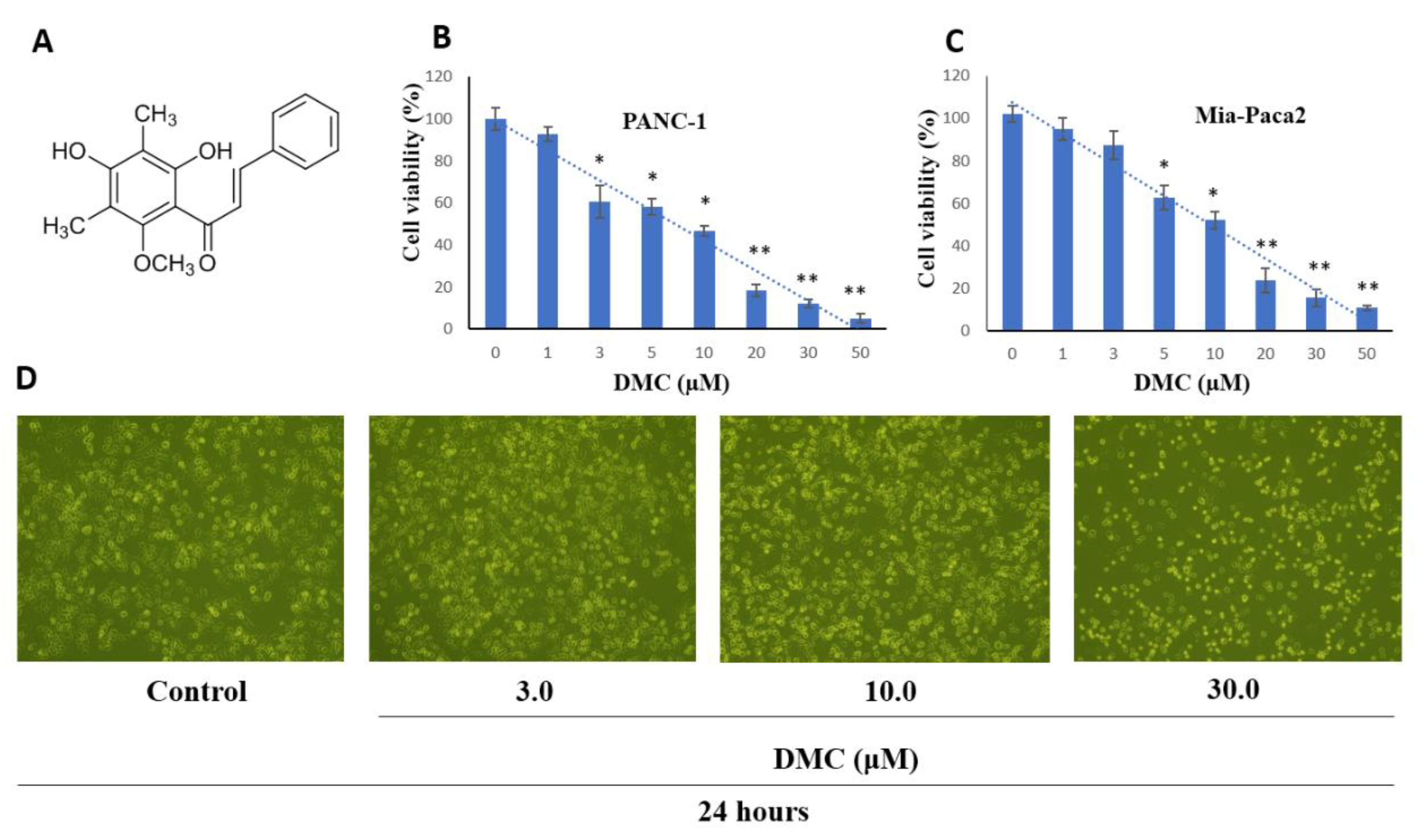
2.1. Cell Proliferation Activity
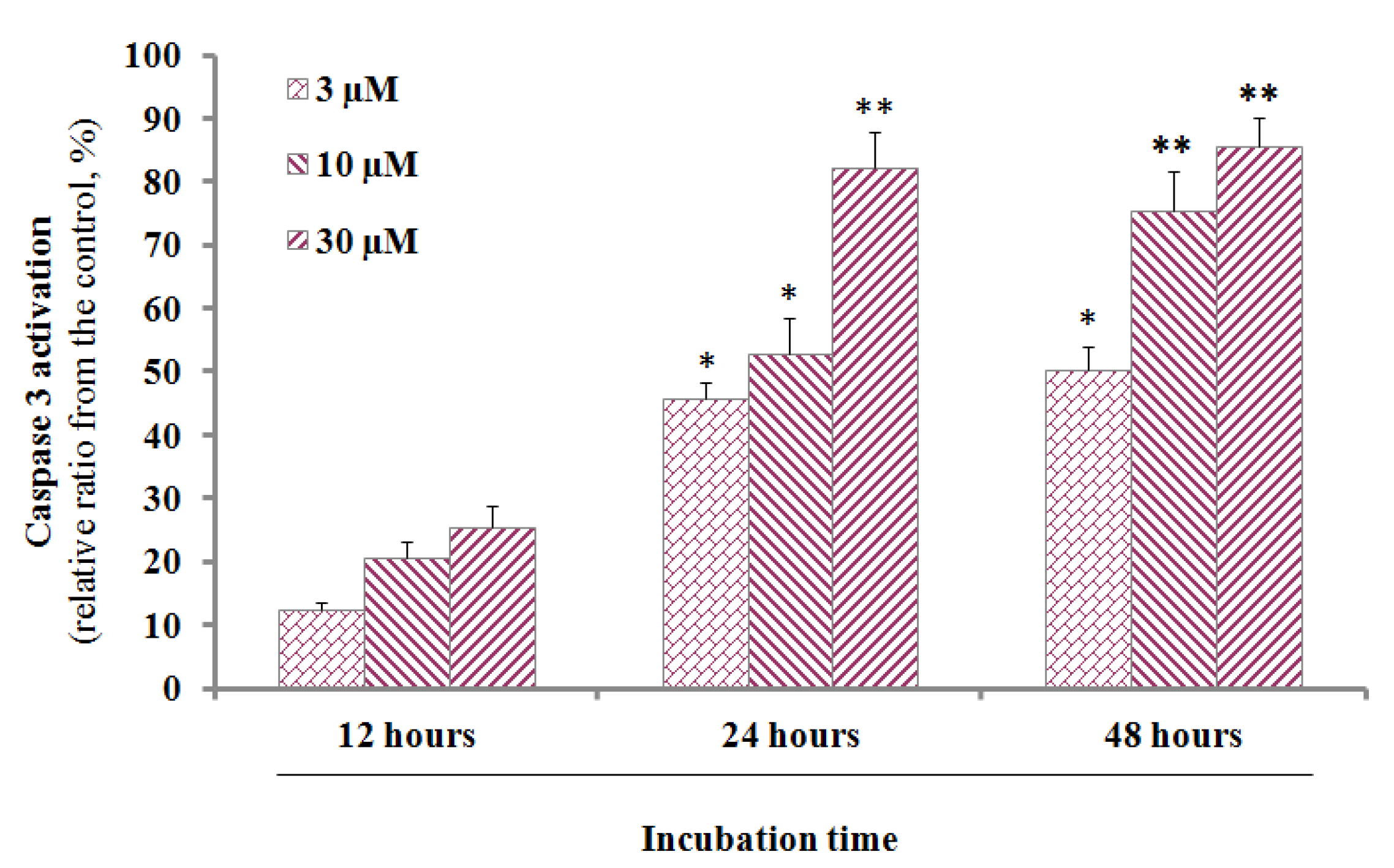
2.2. Caspase-3 activity
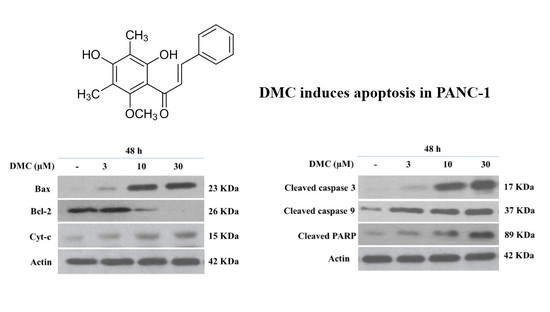
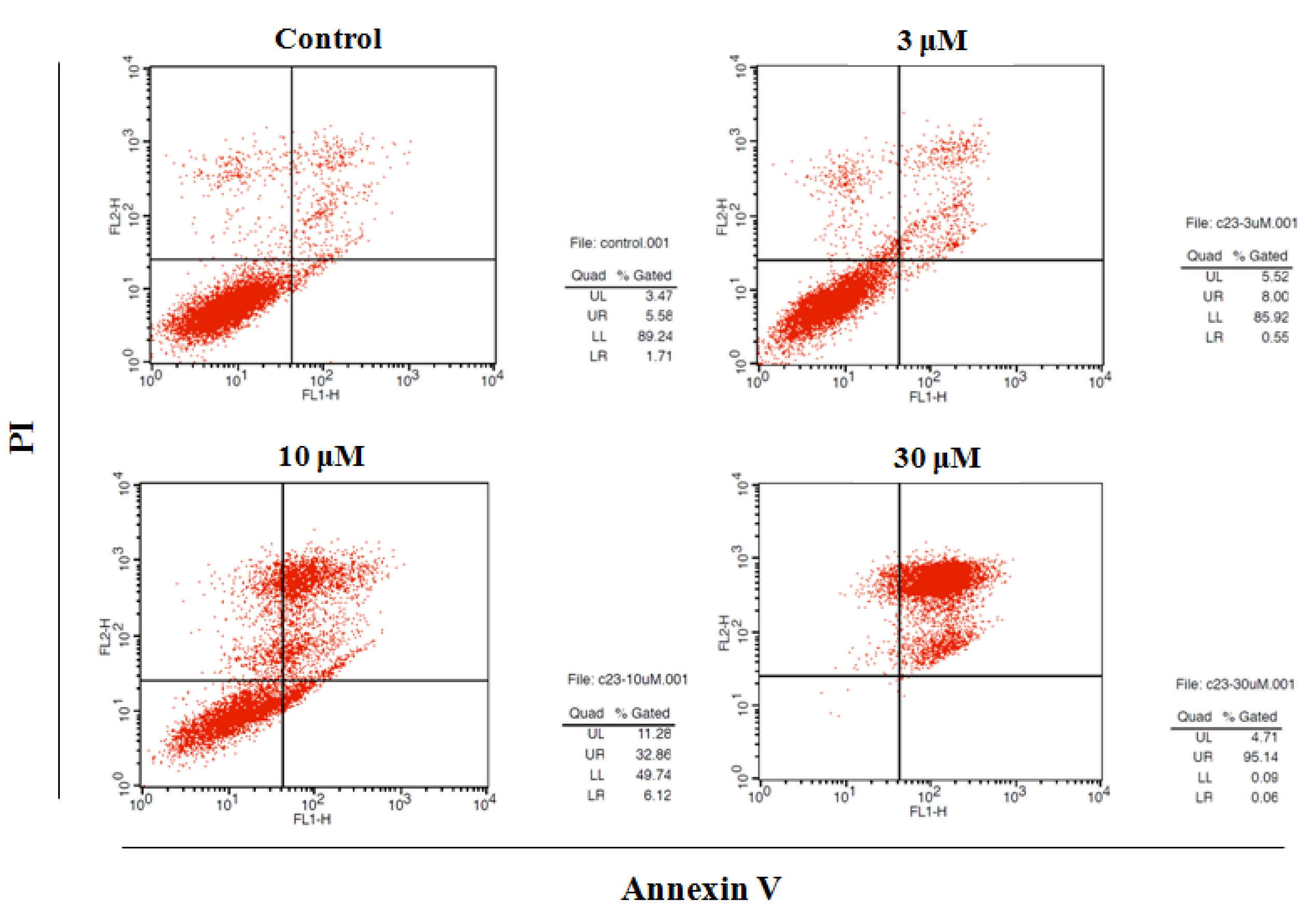
2.3. Induction of Apoptosis by DMC
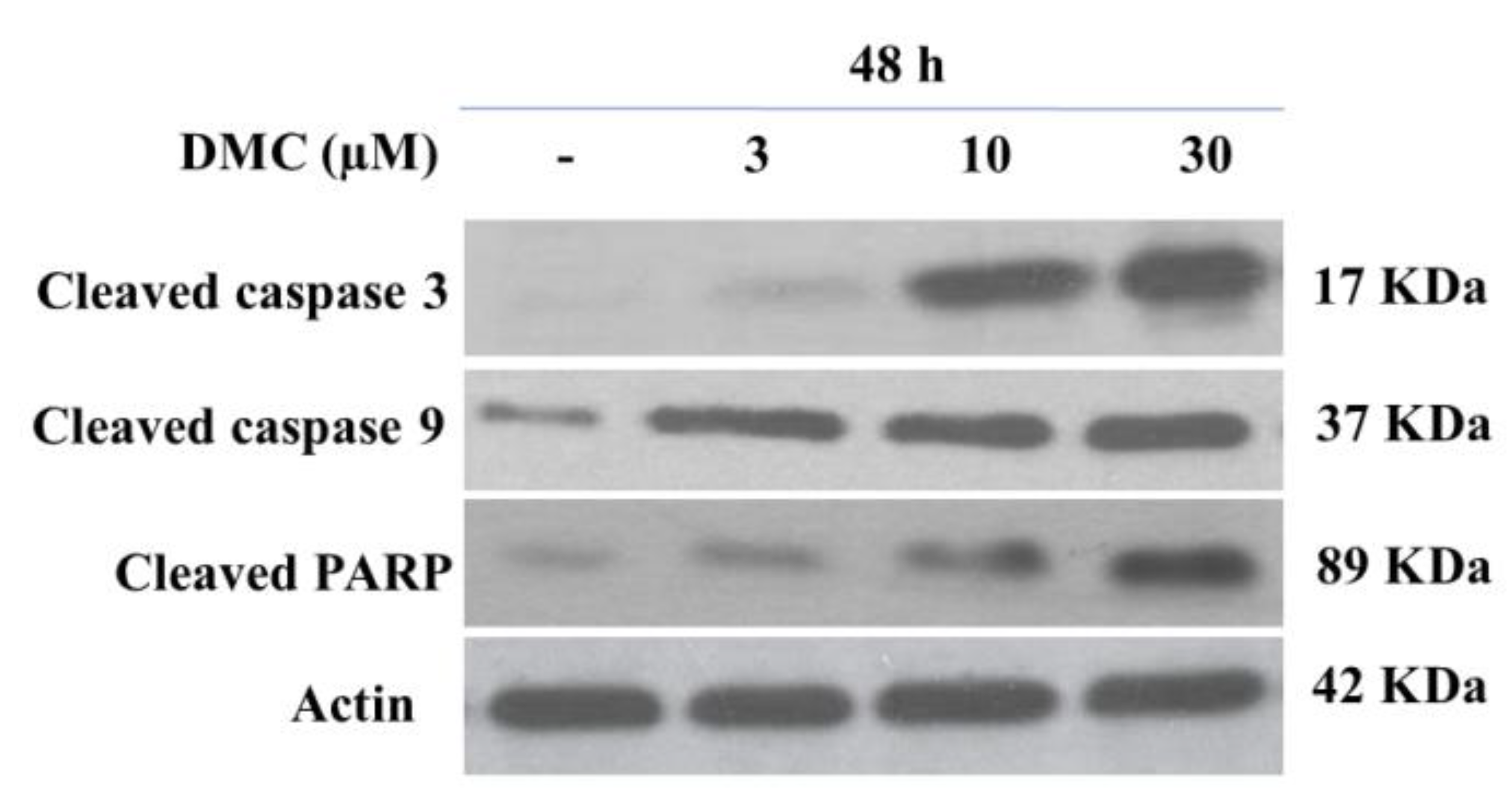
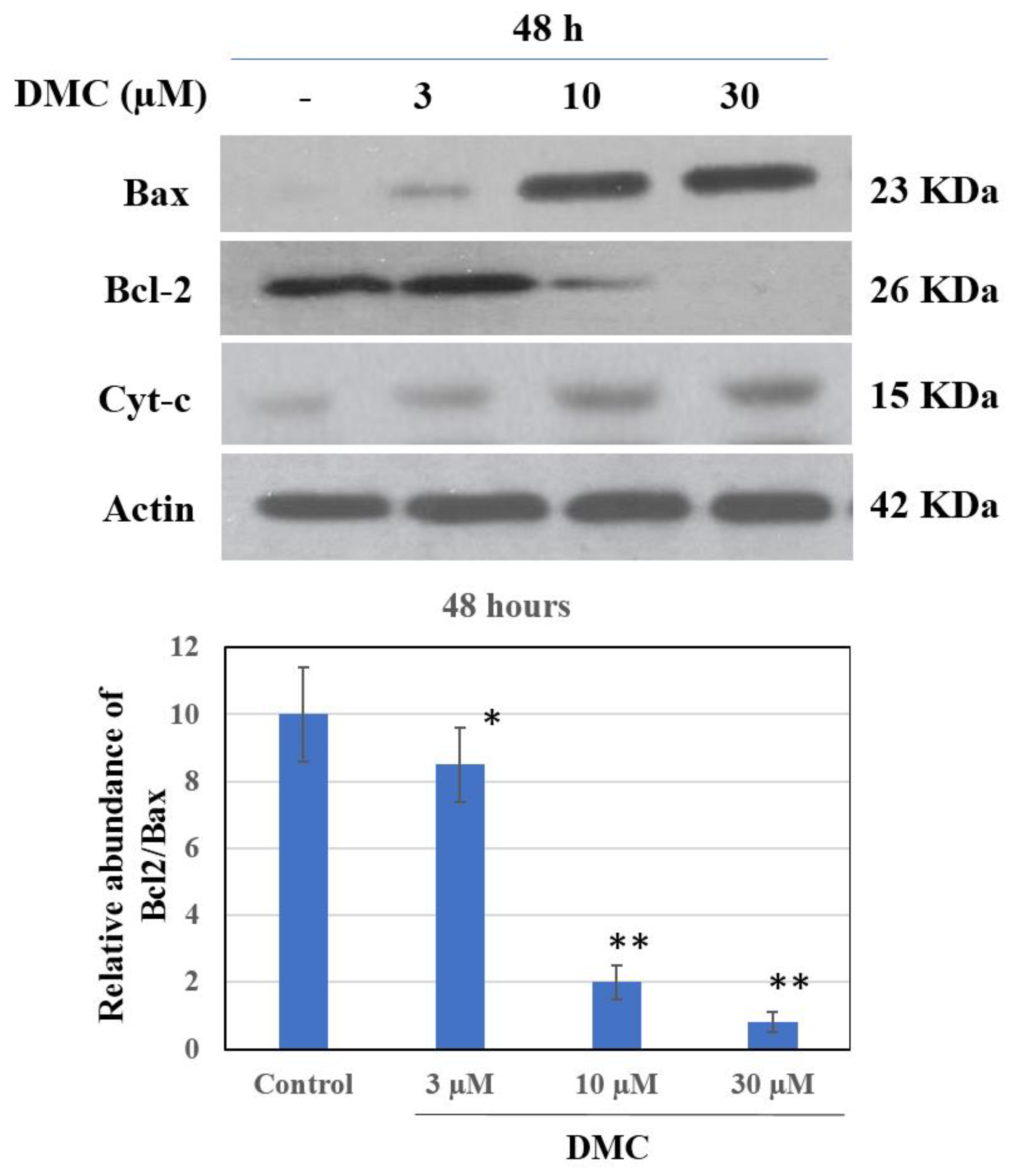
2.4. Effect of DMC on the Expression of Apoptosis-Related Protein
3. Discussion
4. Material and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Isolation of DCM
4.4. Cell Lines and Culture
4.5. Cell Proliferation Activity Assay
4.6. Caspase-3 Actyivation Assay
4.7. Detection of Apoptosis by Double Stanning
4.8. Preparation of Total Cell Extract and Immuno Blot Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Bishop, K.; Altekruse, S.F.; Kosary, C.L.; Yu, M.; Ruhl, J.; Tatalovich, Z.; et al. SEER Cancer Statistics Review, 1975–2013; National Cancer Institute: Bethesda, MD, USA, 2016. [Google Scholar]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. GLOBOCAN 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11; International Agency for Research on Cancer: Lyon, France, 2013. [Google Scholar]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Cascinu, S.; Kleeff, J.; Labianca, R.; Löhr, J.M.; Neoptolemos, J.; Real, F.X.; Van Laethem, J.L.; Heinemann, V. Addressing the challenges of pancreatic cancer: Future directions for improving outcomes. Pancreatology 2013, 15, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.; Herman, J.; Schulick, R.; Hruban, R.H.; Goggins, M. Pancreatic cancer. Lancet 2011, 378, 607–620. [Google Scholar] [CrossRef]
- Ilic, M.; Ilic, I. Epidemiology of pancreatic cancer. World J. Gastroenterol. 2016, 22, 9694–9705. [Google Scholar] [CrossRef] [PubMed]
- Chhoda, A.; Lu, L.; Clerkin, B.M.; Risch, H.; Farrell, J.J. Current approaches to pancreatic cancer screening. Am. J. Pathol. 2019, 189, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhao, G.; Zhao, Y. Combination Immunotherapy Approaches for Pancreatic Cancer Treatment. Can. J. Gastroenterol. Hepatol. 2018, 2018, 6240467. [Google Scholar] [CrossRef] [PubMed]
- Loi, D.T. Vietnamese Medicinal Plants and Ingredients; Medical Publishing House: Hanoi, Vietnam, 2001; pp. 423–424. [Google Scholar]
- Ye, C.L.; Lu, Y.H.; Wei, D.Z. Flavonoids from Cleistocalyx operculatus. Phytochemistry 2004, 65, 445–447. [Google Scholar] [CrossRef]
- Woo, A.Y.; Waye, M.M.; Kwan, H.S.; Chan, M.C.; Chau, C.F.; Cheng, C.H. Inhibition of ATPases by Cleistocalyx operculatus. A possible mechanism for the cardiotonic actions of the herb. Vasc. Pharmacol. 2002, 38, 163–168. [Google Scholar] [CrossRef]
- Mai, T.T.; Chuyen, N.V. Anti-hyperglycemic activity of an aqueous extract from flower buds of Cleistocalyx operculatus (Roxb.) Merr and Perry. Biosci. Biotechnol. Biochem. 2007, 71, 69–76. [Google Scholar] [CrossRef]
- Mai, T.T.; Thu, N.N.; Tien, P.G.; Van Chuyen, N. Alpha-glucosidase inhibitory and antioxidant activities of Vietnamese edible plants and their relationships with polyphenol contents. J. Nutr. Sci. Vitaminol. 2007, 53, 267–276. [Google Scholar] [CrossRef]
- Wang, C.; Wu, P.; Tian, S.; Xue, J.; Xu, L.; Li, H.; Wei, X. Bioactive pentacyclic triterpenoids from the leaves of Cleistocalyx operculatus. J. Nat. Prod. 2016, 79, 2912–2923. [Google Scholar] [CrossRef] [PubMed]
- Su, J.C.; Wang, S.; Cheng, W.; Huang, X.J.; Li, M.M.; Jiang, R.W.; Li, Y.L.; Wang, L.; Ye, W.C.; Wang, Y. Phloroglucinol derivatives with unusual skeletons from Cleistocalyx operculatus and their in vitro antiviral activity. J. Org. Chem. 2018, 83, 8522–8532. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.K.; Dao, T.T.; Nguyen, N.H.; Kim, J.; Kim, E.; Cho, T.O.; Oh, W.K. Antiviral phenolics from the leaves of Cleistocalyx operculatus. Fitoterapia 2016, 110, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Cuong, T.D.; Lee, J.S.; Shin, B.S.; Woo, M.H.; Hung, T.M. Cholinesterase inhibitors from Cleistocalyx operculatus buds. Arch. Pharm. Res. 2010, 33, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Dung, N.T.; Bajpai, V.K.; Yoon, J.I.; Kang, S.C. Anti-inflammatory effects of essential oil isolated from the buds of Cleistocalyx operculatus (Roxb.) Merr and Perry. Food Chem. Toxicol. 2009, 47, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Niu, J.L.; Zhao, L.M.; Lu, Y.H. Reversal effect of 2′,4′-dihydroxy-6′-methoxy-3′,5′-dimethylchalcone on multi-drug resistance in resistant human hepatocellular carcinoma cell line BEL-7402/5-FU. Phytomedicine 2011, 18, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Niu, J.L.; Lu, Y.H. Multidrug resistance reversal effect of DMC derived from buds of Cleistocalyx operculatus in human hepatocellular tumor xenograft model. J. Sci. Food Agric. 2012, 92, 135–140. [Google Scholar] [CrossRef]
- Yu, W.G.; Qian, J.; Lu, Y.H. Hepatoprotective effects of 2′, 4′-dihydroxy-6′-methoxy-3′, 5′-dimethylchalcone on CCl4-induced acute liver injury in mice. J. Agric. Food Chem. 2011, 59, 12821–12829. [Google Scholar] [CrossRef]
- Su, M.Y.; Huang, H.Y.; Li, L.; Lu, Y.H. Protective effects of 2′, 4′-dihydroxy-6′-methoxy-3′, 5′-dimethylchalcone to PC12 cells against cytotoxicity induced by hydrogen peroxide. J. Agric. Food Chem. 2011, 59, 521–527. [Google Scholar] [CrossRef]
- Ye, C.L.; Lai, Y.F. 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone, from buds of Cleistocalyxoperculatus, induces apoptosis in human hepatoma SMMC-7721 cells through a reactive oxygen species-dependent mechanism. Cytotechnology 2016, 68, 331–341. [Google Scholar] [CrossRef]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Kokuryo, T.; Yokoyama, Y.; Suzuki, H.; Itatsu, K.; Nakagawa, A.; Mizutani, T.; Miyake, T.; Uno, M.; Yamauchi, K.; et al. Antitumor effects of α-bisabolol against pancreatic cancer. Cancer Sci. 2011, 102, 2199–2205. [Google Scholar] [CrossRef] [PubMed]
- Paluszkiewicz, P.; Smolińska, K.; Dębińska, I.; Turski, W.A. Main dietary compounds and pancreatic cancer risk. The quantitative analysis of case-control and cohort studies. Cancer Epidemiol. 2012, 36, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.J.; Robinson, D.P.; Stolzenberg-Solomon, R.Z.; Bamlet, W.R.; de Andrade, M.; Oberg, A.L.; Hammer, T.J.; Rabe, K.G.; Anderson, K.E.; Olson, J.E.; et al. Fruit and vegetable consumption is inversely associated with having pancreatic cancer. Cancer Causes Control 2011, 22, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Lugo, A.; Lagiou, P.; Zucchetto, A.; Polesel, J.; Serraino, D.; Negri, E.; Trichopoulos, D.; La Vecchia, C. Proanthocyanidins and other flavonoids in relation to pancreatic cancer: A case-control study in Italy. Ann. Oncol. 2012, 23, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.; Liu, B.; Jones, P.; Houghton, P.J.; Mota-Filipe, H.; Paulo, A. Cytoprotective effect of Coreopsis tinctoria extracts and flavonoids on tBHP and cytokine-induced cell injury in pancreatic MIN6 cells. J. Ethnopharmacol. 2012, 139, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.Y.; Zhang, L.; Moro, A.; Chen, M.C.; Harris, D.M.; Eibl, G.; Go, V.L. Detection of baicalin metabolites baicalein and oroxylin-a in mouse pancreas and pancreatic xenografts. Pancreas 2012, 41, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, H.; Salmani, J.M.; Fu, R.; Chen, B. Advances of wogonin, an extract from Scutellaria baicalensis for the treatment of multiple tumors. Onco Targets Ther. 2016, 9, 2935–2943. [Google Scholar] [PubMed]
- Ye, C.L.; Liu, J.W.; Wei, D.Z.; Lu, Y.H.; Qian, F. In vitro anti-tumor activity of 2’, 4’-dihydroxy-6’-methoxy-3’, 5’-dimethylchalcone against six established human cancer cell lines. Pharmacol. Res. 2004, 50, 505–510. [Google Scholar] [CrossRef]
- Ye, C.L.; Qian, F.; Wei, D.Z.; Lu, Y.H.; Liu, J.W. Induction of apoptosis in K562 human leukemia cells by 2′, 4′-dihydroxy-6′-methoxy-3′, 5′-dimethylchalcone. Leuk. Res. 2005, 29, 887–892. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



| LL (Annexin V−/PI–) | LR (Annexin V+/PI–) | UL (Annexin V–/PI+) | UR (Annexin V+/PI+) | |
| Control | 92.4 ± 3.7 | 2.8 ± 1.0 | 3.50 ± 0.8 | 6.1 ± 0.9 |
| DMC (3 μM) | 85.4 ± 2.3 | 0.5 ± 0.2 | 5.5 ± 1.0 | 8.2 ± 1.4 |
| DMC (10 μM) | 50.7 ± 4.4 | 6.2 ± 1.5 | 12.0 ± 2.5 | 32.8 ± 2.8 |
| DMC (30 μM) | 1.0 ± 0.3 | 0.1 ± 0.1 | 5.2 ± 0.5 | 95.1 ± 1.6 |


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuan, H.N.; Minh, B.H.; Tran, P.T.; Lee, J.H.; Oanh, H.V.; Ngo, Q.M.T.; Nguyen, Y.N.; Lien, P.T.K.; Tran, M.H. The Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′- dimethylchalcone from Cleistocalyx operculatus Buds on Human Pancreatic Cancer Cell Lines. Molecules 2019, 24, 2538. https://doi.org/10.3390/molecules24142538
Tuan HN, Minh BH, Tran PT, Lee JH, Oanh HV, Ngo QMT, Nguyen YN, Lien PTK, Tran MH. The Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′- dimethylchalcone from Cleistocalyx operculatus Buds on Human Pancreatic Cancer Cell Lines. Molecules. 2019; 24(14):2538. https://doi.org/10.3390/molecules24142538
Chicago/Turabian StyleTuan, Huynh Nhu, Bui Hoang Minh, Phuong Thao Tran, Jeong Hyung Lee, Ha Van Oanh, Quynh Mai Thi Ngo, Yen Nhi Nguyen, Pham Thi Kim Lien, and Manh Hung Tran. 2019. "The Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′- dimethylchalcone from Cleistocalyx operculatus Buds on Human Pancreatic Cancer Cell Lines" Molecules 24, no. 14: 2538. https://doi.org/10.3390/molecules24142538
APA StyleTuan, H. N., Minh, B. H., Tran, P. T., Lee, J. H., Oanh, H. V., Ngo, Q. M. T., Nguyen, Y. N., Lien, P. T. K., & Tran, M. H. (2019). The Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′- dimethylchalcone from Cleistocalyx operculatus Buds on Human Pancreatic Cancer Cell Lines. Molecules, 24(14), 2538. https://doi.org/10.3390/molecules24142538