
Effect of Clove and Thyme Essential Oils on Candida Biofilm Formation and the Oil Distribution in Yeast Cells


Abstract
1. Introduction
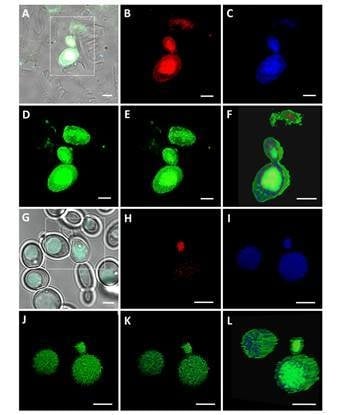
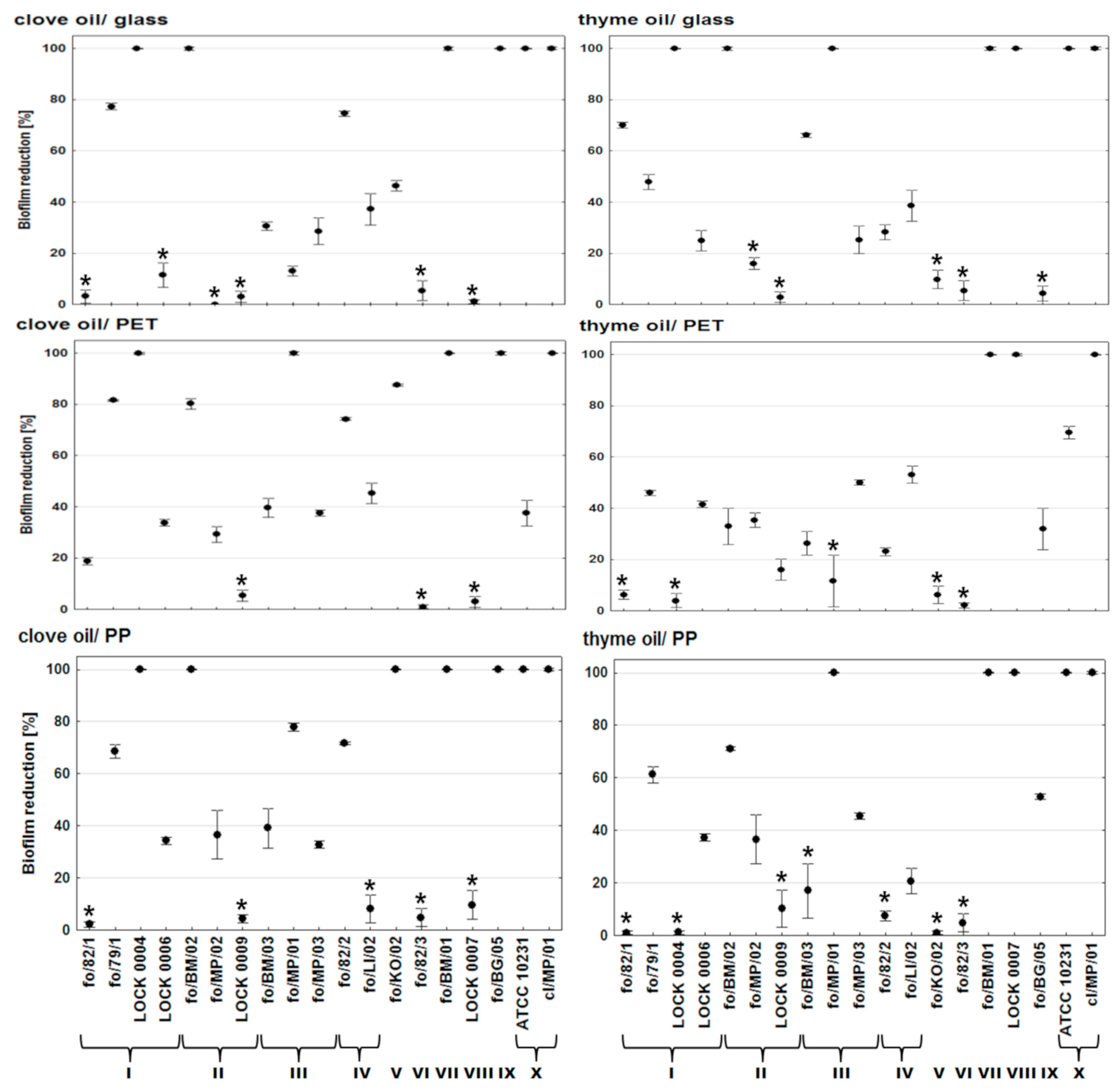
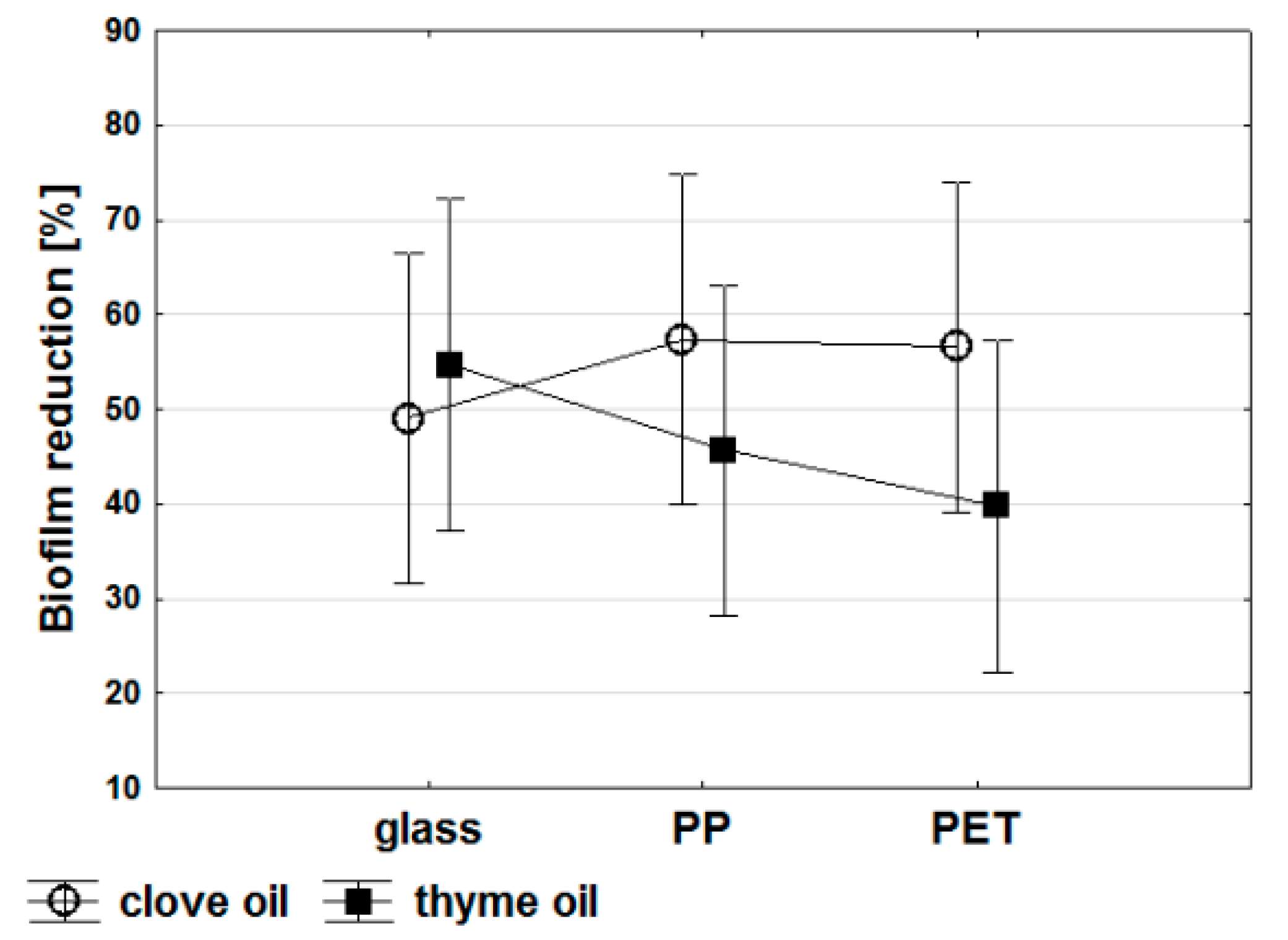
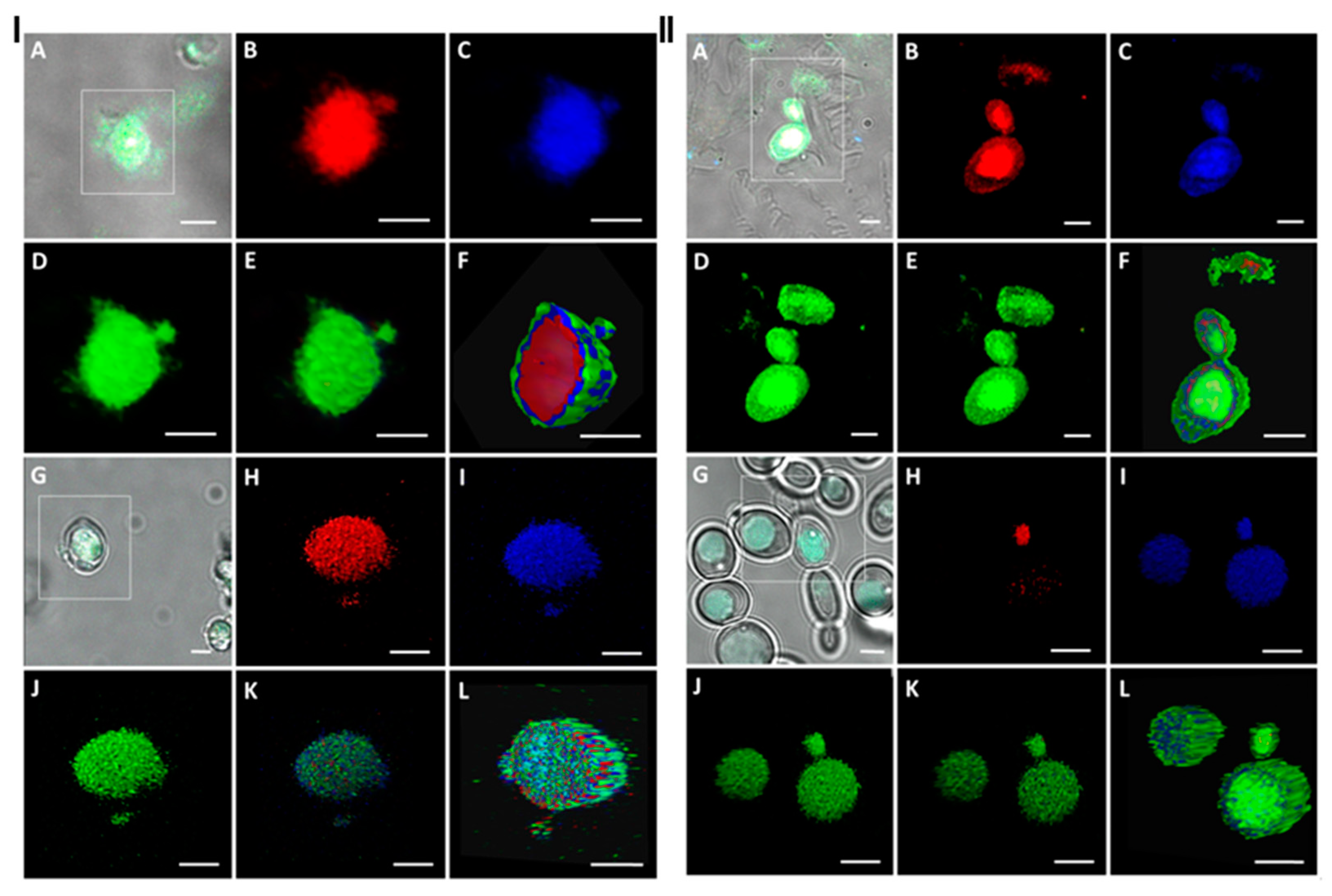
2. Results and Discussion
3. Materials and Methods
3.1. Essential Oils
3.2. Yeasts
3.3. Biofilm Formation
3.4. Observations using Confocal Laser Scanning Microscope (CLSM)
3.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tascini, C.; Sozio, E.; Corte, L.; Sbrana, F.; Scarparo, C.; Ripoli, A.; Bertolino, G.; Merelli, M.; Tagliaferri, E.; Corcione, A.; et al. The role of biofilm forming on mortality in patients with candidemia: A study derived from real world data. Infect. Dis. 2018, 50, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Stratford, M. Food and beverage spoilage yeasts. In Yeasts in Food and Beverages; Querol, A., Fleet, G.H., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2006; pp. 335–379. [Google Scholar] [CrossRef]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormik, T.; Ghanoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [PubMed]
- Simoes, M.; Simoes, L.C.; Vieira, M.J. A review of current and emergent biofilm control strategies. LWT Food Sci. Technol. 2010, 43, 573–583. [Google Scholar] [CrossRef]
- Baillie, G.S.; Douglas, L.J. Matrix polymers of Candida biofilms and their possible role in biofilm resistance to antifungal agents. J. Antimicrobial. Chemother. 2000, 46, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Bachman, S.; Patterson, T.F.; Wickes, B.L.; Lopez-Ribot, J.L. Investigation of multidrug efflux pumps in relations to fluconazole resistance in Candida albicans biofilms. J. Antimicrobial. Chemother. 2002, 49, 973–980. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of fluconazole resistance in Candida albicans biofilm: Phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef]
- Al-Fattani, M.A.; Douglas, L.J. Penetration of Candida biofilms by antifungal agents. Antimicrob. Agents Chemother. 2004, 48, 3291–3297. [Google Scholar] [CrossRef]
- LaFleur, M.; Kumamoto, C.A.; Lewis, K. Candida albicans biofilms produce antifungal-tolerant persister cells. Antimicrobial. Agents Chemother. 2006, 50, 3839–3846. [Google Scholar] [CrossRef]
- Rajkowska, K.; Kunicka-Styczyńska, A.; Maroszyńska, M. Selected essential oils as antifungal agents against antibiotic-resistant Candida spp.: In vitro study on clinical and food-borne isolates. Microb. Drug Resist. 2017, 23, 18–24. [Google Scholar] [CrossRef]
- Rajkowska, K.; Kunicka-Styczyńska, A. Molecular typing and virulence index of food-borne Candida spp. Isolates—A group of potential emerging pathogens. Int. J. Food Microbiol. 2018, 279, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Targalska, M.; Kunicka-Styczyńska, A. Candida biofilms: Environmental and clinical aspects. In The Yeast Role in Medical Applications; Abdulkhair, W.M.H., Ed.; IntechOpen: London, UK, 2018; pp. 344–350. [Google Scholar]
- Ramage, G.; Martínez, J.P.; López-Ribot, J.L. Candida biofilms on implanted biomaterials: A clinically significant problem. FEMS Yeast Res. 2006, 6, 979–986. [Google Scholar] [CrossRef]
- Khan, W.; Muntimadugu, E.; Jaffe, M.; Domb, A.J. Implantable medical devices. In Focal Controlled Drug Delivery. Advances in Delivery Science and Technology; Domb, A., Khan, W., Eds.; Springer: Boston, MA, USA, 2014; pp. 33–59. [Google Scholar]
- Arkles, B. Hydrophobicity, hydrophilicity and silanes. Paint. Coating. Ind. 2006, 22, 114–123. [Google Scholar]
- Hyde, F.W.; Alberg, M.; Smith, K. Comparison of fluorinated polymers against stainless steel, glass and polypropylene in microbial biofilm adherence and removal. J. Ind. Microbiol. Biotechnol. 1997, 19, 142–149. [Google Scholar] [CrossRef]
- Wolfmeier, H.; Pletzer, D.; Mansour, S.C.; Hancock, R.E.W. New perspectives in biofilm eradication. ACS Infect. Dis. 2018, 4, 93–106. [Google Scholar] [CrossRef]
- Khan, M.S.; Ahmad, I. Biofilm inhibition by Cymbopogon citratus and Syzygium aromaticum essential oils in the strains of Candida albicans. J. Ethnopharmacol. 2012, 140, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Lal, P.; Pruthi, V. Prevention of Candida albicans biofilm by plant oils. Mycopathologia 2008, 165, 13–19. [Google Scholar] [CrossRef]
- Sookto, T.; Srithavaj, T.; Thaweboon, S.; Thaweboon, B.; Shrestha, B. In vitro effects of Salvia officinalis L. essential oil on Candida albicans. Asian Pac. J. Trop. Biomed. 2013, 3, 376–380. [Google Scholar] [CrossRef]
- Khan, M.S.; Ahmad, I.; Cameotra, S.S.; Botha, F. Sub-MICs of Carum copticum and Thymus vulgaris influence virulence factors and biofilm formation in Candida spp. BMC Complement. Altern. Med. 2014, 14, 337. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Khan, A.; Manzoor, N. Reversal of efflux mediated antifungal resistance underlies synergistic activity of two monoterpenes with fluconazole. Eur. J. Pharm. Sci. 2013, 48, 80–86. [Google Scholar] [CrossRef]
- Stringaro, A.; Vavala, E.; Colone, M.; Pepi, F.; Mignogna, G.; Garzoli, S.; Cecchetti, S.; Ragno, R.; Angiolella, L. Effects of Mentha suaveolens essential oil alone or in combination with other drugs in Candida albicans. Evid. Based Complement. Altern. Med. 2014, 2014, 12590. [Google Scholar] [CrossRef]
- Pemmaraju, S.C.; Pruthi, P.A.; Prasad, R.; Pruthi, V. Candida albicans biofilm inhibition by synergistic action of terpenes and fluconazole. Indian J. Exp. Biol. 2013, 51, 1032–1037. [Google Scholar]
- Kerekes, E.B.; Deak, E.; Tako, M.; Tserennadmid, R.; Petkovits, T.; Vagvolgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related microorganisms. J. Appl. Microbiol. 2013, 115, 933–942. [Google Scholar] [CrossRef]
- Tumbarello, M.; Posteraro, B.; Trecarichi, E.M.; Fiori, B.; Rossi, M.; Porta, R. Biofilm production by Candida species and inadequate antifungal therapy as predictors of mortality for patients with candidemia. J. Clin. Microbiol. 2007, 45, 1843–1850. [Google Scholar] [CrossRef]
- Dalle, F.; Wächtler, B.; L’Ollivier, C.; Holland, G.; Bannert, N.; Wilson, D.; Labruère, C.; Bonnin, A.; Hube, B. Cellular interactions of Candida albicans with human oral epithelial cells and enterocytes. Cell. Microbiol. 2010, 12, 248–271. [Google Scholar] [CrossRef]
- Segal, E. Experimental infection. In Pathogenic Yeasts and Yeast Infections; Segal, E., Baum, G.L., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1994; pp. 71–79. [Google Scholar]
- Chandra, J.; Patel, J.D.; Li, J.; Zhou, G.; Mukherjee, P.K.; McCormick, T.S.; Anderson, J.M.; Ghannoum, M.A. Modification of surface properties of biomaterials influences the ability of Candida albicans to form biofilms. Appl. Environ. Microbiol. 2005, 71, 8795–8801. [Google Scholar] [CrossRef]
- Llana-Ruiz-Cabello, M.; Pichardo, S.; Bermudez, J.M.; Banos, A.; Nunez, C.; Guillamon, E.; Aucejo, S.; Camean, A.M. Development of PLA films containing oregano essential oil (Origanum vulgare L. virens) intended for use in food packaging. Food Addit. Contam. Part. A 2016, 33, 1374–1386. [Google Scholar] [CrossRef]
- Cui, H.; Bai, M.; Lin, L. Plasma-treated poly(ethylene oxide) nanofibers containing tea tree oil/beta-cyclodextrin inclusion complex for antibacterial packaging. Carbohydr. Polym. 2018, 179, 360–369. [Google Scholar] [CrossRef]
- Roshchina, V.V.; Kuchin, A.V.; Yashin, V.A. Application of autofluorescence for analysis of medicinal plants. Int. J. Spectrosc. 2017, 2017, 1–8. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Zhu, X.; Cao, P.; Wei, S.; Lu, Y. Antibacterial and antibiofilm activities of eugenol from essential oil of Syzygium aromaticum (L.) Merr. & L. M. Perry (clove) leaf against periodontal pathogen Porphyromonas gingivalis. Microb. Pathog. 2017, 113, 396–402. [Google Scholar] [CrossRef]
- Boschi, F.; Fontanella, M.; Calderan, L.; Sbarbati, A. Luminescence and fluorescence of essential oils. Fluorescence imaging in vivo of wild chamomile oil. Eur. J. Histochem. 2011, 55, 97–100. [Google Scholar] [CrossRef]
- Rajkowska, K.; Nowak, A.; Kunicka-Styczyńska, A.; Siadura, A. Biological effects of various chemically characterized essential oils: Investigation of the mode of action against Candida albicans and HeLa cells. RSC Adv. 2016, 6, 97199–97207. [Google Scholar] [CrossRef]
- Ramsay, L.M.; Gadd, G.M. Mutants of Saccharomyces cerevisiae defective in vacuolar function confirm a role for the vacuole in toxic metal ion detoxification. FEMS Microbiol. Lett. 1997, 152, 293–298. [Google Scholar] [CrossRef]
- Cornelius, G.; Nakashima, H. Vacuoles play a decisive role in calcium homeostasis in Neurospora crassa. J. Gen. Microbiol. 1987, 133, 2341–2347. [Google Scholar] [CrossRef][Green Version]
- Armstrong, J. Yeast vacuoles: More than a model lysosome. Trends Cell. Biol. 2010, 20, 580–585. [Google Scholar] [CrossRef]
- Palmer, G.E. Vacuolar trafficking and Candida albicans pathogenesis. Commun. Integr. Biol. 2011, 4, 240–242. [Google Scholar] [CrossRef][Green Version]
Sample Availability: Not available. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RI 1 | Clove Oil | Thyme Oil |
|---|---|---|---|
| Content [%] | |||
| α-Thujene | 926 | − | 0.9 |
| α-Pinene | 934 | − | 0.9 |
| Campehen | 940 | − | 0.4 |
| β-Myrcene | 983 | − | 1.8 |
| Car-3-ene | 1008 | − | 2 |
| p-Cymene | 1016 | − | 18.4 |
| β-Phellandrene | 1019 | − | 0.4 |
| Limonene | 1025 | − | 0.9 |
| γ-Terpinene | 1055 | − | 8.8 |
| Linalool | 1086 | − | 3.2 |
| Borneol | 1155 | − | 0.7 |
| Thymol | 1281 | − | 48.6 |
| Carvacrol | 1285 | − | 5.5 |
| Eugenol | 1342 | 85.2 | − |
| (E)-β-Caryophyllene | 1421 | 9.9 | 2.3 |
| α-Humulene | 1453 | 1.9 | 0.1 |
| δ-Cadinene | 1513 | 0.4 | 0.2 |
| (E)-β-Caryophyllene oxide | 1568 | 0.4 | 0.4 |
| Species | Isolate | Origin |
|---|---|---|
| C. lusitaniae | fo/82/1, fo/79/1 | fruit yoghurt |
| LOCK 0004, LOCK 0006 | feed | |
| C. krusei | fo/BM/02, fo/MP/02 | pickled cucumber |
| LOCK 0009 | bakery | |
| C. boidinii | fo/BM/03, fo/MP/01, fo/MP/03 | pickled cucumber |
| C. famata | fo/82/2 | fruit yoghurt |
| fo/LI/02 | herring salad | |
| C. colliculosa | fo/KO/02 | fruit yoghurt |
| C. parapsilosis | fo/82/3 | fruit yoghurt |
| C. tropicalis | fo/BM/01 | pickled cucumber |
| C. pelliculosa | LOCK 0007 | feed |
| C. rugosa | fo/BG/05 | sauerkraut |
| Strain | Clove Oil | Thyme Oil |
|---|---|---|
| Concentration [% v/v] | ||
| C. lusitaniae fo/82/1 | 0.12 | 0.06 |
| C. lusitaniae fo/79/1 | 0.25 | 0.12 |
| C. lusitaniae LOCK 0004 | 0.12 | 0.12 |
| C. lusitaniae LOCK 0006 | 0.06 | 0.12 |
| C. krusei fo/BM/02 | 0.12 | 0.12 |
| C. krusei fo/MP/02 | 0.06 | 0.03 |
| C. krusei LOCK 0009 | 0.12 | 0.12 |
| C. boidinii fo/BM/03 | 0.12 | 0.25 |
| C. boidinii fo/MP/01 | 0.12 | 0.12 |
| C. boidinii fo/MP/03 | 0.06 | 0.03 |
| C. famata fo/82/2 | 0.12 | 0.03 |
| C. famata fo/LI/02 | 0.03 | 0.03 |
| C. colliculosa fo/KO/02 | 0.12 | 0.12 |
| C. parapsilosis fo/82/3 | 0.06 | 0.06 |
| C. tropicalis fo/BM/01 | 8 | 8 |
| C. pelliculosa LOCK 0007 | 0.06 | 0.06 |
| C. rugosa fo/BG/05 | 0.12 | 0.12 |
| C. albicans ATCC 10231 | 0.25 | 0.25 |
| C. albicans cl/MP/01 | 0.25 | 0.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajkowska, K.; Nowicka-Krawczyk, P.; Kunicka-Styczyńska, A. Effect of Clove and Thyme Essential Oils on Candida Biofilm Formation and the Oil Distribution in Yeast Cells. Molecules 2019, 24, 1954. https://doi.org/10.3390/molecules24101954
Rajkowska K, Nowicka-Krawczyk P, Kunicka-Styczyńska A. Effect of Clove and Thyme Essential Oils on Candida Biofilm Formation and the Oil Distribution in Yeast Cells. Molecules. 2019; 24(10):1954. https://doi.org/10.3390/molecules24101954
Chicago/Turabian StyleRajkowska, Katarzyna, Paulina Nowicka-Krawczyk, and Alina Kunicka-Styczyńska. 2019. "Effect of Clove and Thyme Essential Oils on Candida Biofilm Formation and the Oil Distribution in Yeast Cells" Molecules 24, no. 10: 1954. https://doi.org/10.3390/molecules24101954
APA StyleRajkowska, K., Nowicka-Krawczyk, P., & Kunicka-Styczyńska, A. (2019). Effect of Clove and Thyme Essential Oils on Candida Biofilm Formation and the Oil Distribution in Yeast Cells. Molecules, 24(10), 1954. https://doi.org/10.3390/molecules24101954