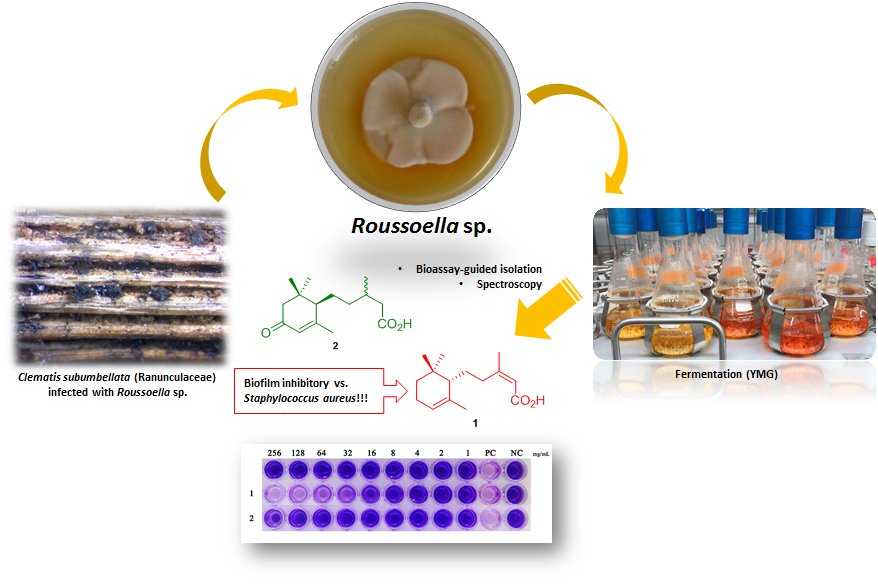
Biofilm Inhibitory Abscisic Acid Derivatives from the Plant-Associated Dothideomycete Fungus, Roussoella sp.

 ,
, 

Abstract
:
1. Introduction
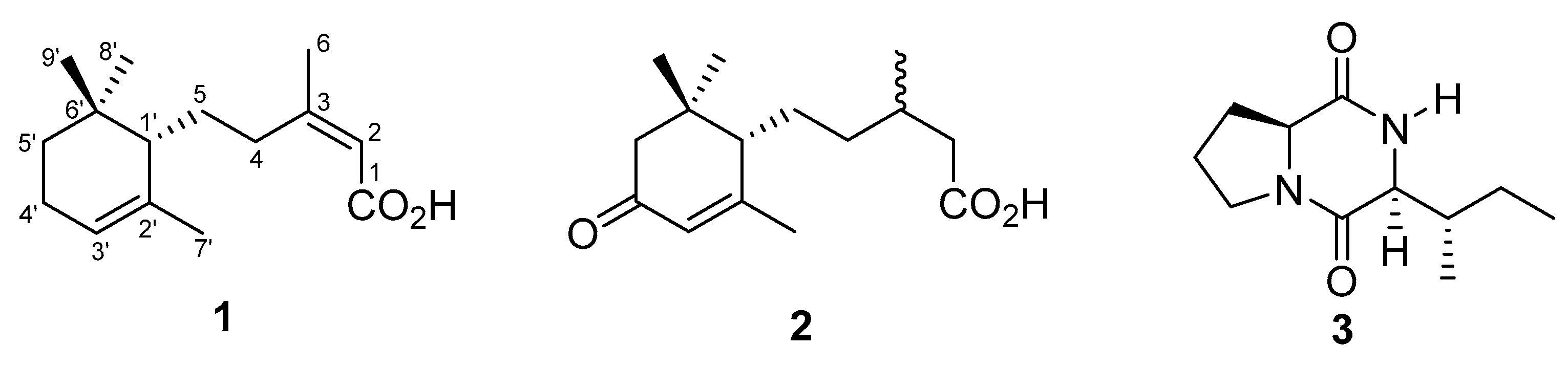
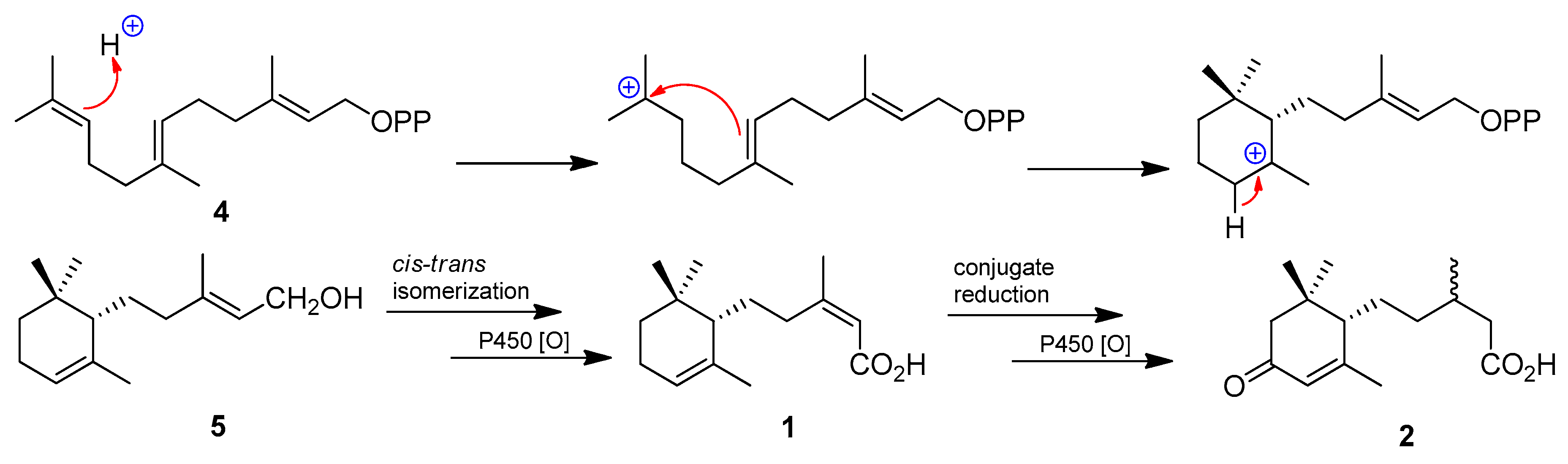
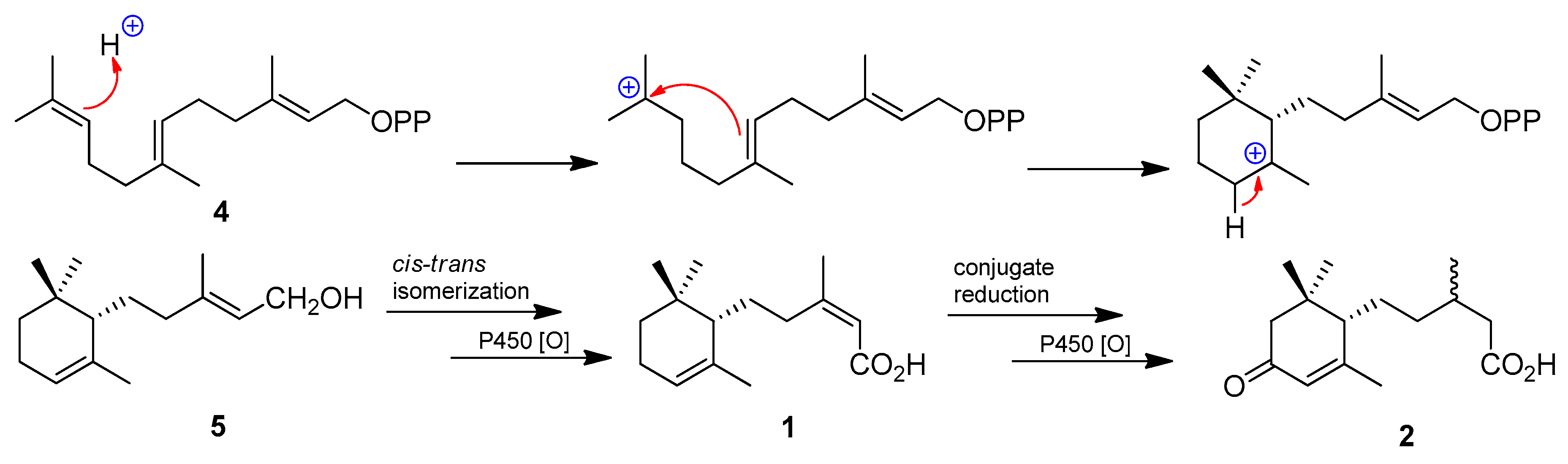
2. Results and Conclusions
3. Materials and Methods
3.1. General Information
3.2. Fungal Material
3.3. Fermentation and Extraction
3.4. Isolation of 1–3
3.5. Biological Activities
Antimicrobial Activity and Cytotoxicity Assays
3.6. Inhibition of Biofilm Formation
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Beimforde, C.; Feldberg, K.; Nylinder, S.; Rikkinen, J.; Tuovila, H.; Dörfelt, H.; Gube, M.; Jackson, D.J.; Reitner, J.; Seyfullah, L.J.; et al. Estimating the phanerozoic history of the Ascomycota lineages: Combining fossil and molecular data. Mol. Phylogenet. Evol. 2014, 78, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008; pp. 55–56. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, J.K.; Maharachchikumbura, S.S.; Ekanayaka, A.H.; Tian, Q.; Phookamsak, R. Outline of Ascomycota: 2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Chagas, F.; Caraballo-Rodriguez, A.; Pupo, M. Endophytic fungi as a source of novel metabolites. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Zeilinger, S., Martín, J.F., García-Estrada, C., Eds.; Fungal Biology; Springer: New York, NY, USA, 2015; Volume 2, pp. 123–176. [Google Scholar]
- Gutierrez, R.M.P.; Gonzalez, A.M.N.; Ramirez, A.M. Compounds derived from endophytes: A review of phytochemistry and pharmacology. Curr. Med. Chem. 2012, 19, 2992–3030. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.J.M.; Chooi, Y.H.; Breen, S.; Solomon, P.S. The past, present and future of secondary metabolite research in the Dothideomycetes. Mol. Plant Pathol. 2015, 16, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.K.; Phookamsak, R.; Dai, D.Q.; Tanaka, K.; Jones, E.G.; Xu, J.C.; Chukeatirote, E.; Hyde, K.D. Roussoellaceae, a new pleosporalean family to accommodate the genera Neoroussoella gen. nov., Roussoella and Roussoellopsis. Phytotaxa 2014, 181, 1–33. [Google Scholar] [CrossRef]
- Takekawa, H.; Tanaka, K.; Fukushi, E.; Matsuo, K.; Nehira, T.; Hashimoto, M. Roussoellols A and B, tetracyclic fusicoccanes from Roussoella hysterioides. J. Nat. Prod. 2013, 76, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Honmura, Y.; Takekawa, H.; Tanaka, K.; Maeda, H.; Nehira, T.; Hehre, W.; Hashimoto, M. Computation-assisted structural elucidation of epoxyroussoeone and epoxyroussoedione isolated from Roussoella japanensis KT1651. J. Nat. Prod. 2015, 78, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, E.L.F.; Williams, D.E.; Ióca, L.P.; Morais-Urano, R.P.; Santos, M.F.C.; Patrick, B.O.; Elias, L.M.; Lira, S.P.; Ferreira, A.G.; Passarini, M.R.Z.; et al. Structure and biogenesis of roussoellatide, a dichlorinated polyketide from the marine-derived fungus Roussoella sp. DLM33. Org. Lett. 2015, 17, 5152–5155. [Google Scholar] [CrossRef] [PubMed]
- Millborrow, B.V. The origin of the methyl groups of abscisic acid. Phytochemistry 1975, 14, 2403–2405. [Google Scholar] [CrossRef]
- Soorukram, D.; Knochel, P. Enantioselective synthesis of α-ionone derivatives using an anti SN2 substitution of functionalized zinc organometallics. Org. Lett. 2004, 6, 2409–2411. [Google Scholar] [CrossRef] [PubMed]
- Oritani, T.; Yamashita, K. Studies on abscisic acid. Agric. Biol. Chem. 1970, 34, 830–837. [Google Scholar] [CrossRef]
- Liu, S.; Dai, H.; Heering, C.; Jania, C.; Lin, W.; Liu, Z.; Proksch, P. Inducing new secondary metabolites through co-cultivation of the fungus Pestalotiopsis sp. with the bacterium Bacillus subtilis. Tetrahedron Lett. 2017, 58, 257–261. [Google Scholar] [CrossRef]
- Pedras, M.; Soledade, C.; Yu, Y.; Liu, J.; Tandron-Moya, Y.A. Metabolites produced by the phytopathogenic fungus Rhizoctonia solani: Isolation, chemical structure determination, syntheses and bioactivity. Z. Naturforsch. C J. Biosci. 2005, 60, 717–722. [Google Scholar] [CrossRef]
- Kettner, J.; Dörffling, K. Biosynthesis and metabolism of abscisic acid in tomato leaves infected with Botrytis cinerea. Planta 1995, 196, 627–634. [Google Scholar] [CrossRef]
- Krochko, J.E.; Abrams, G.D.; Loewen, M.K.; Abrams, S.R.; Cutler, A.J. (+)-Abscisic acid 8′-hydroxylase is a cytochrome P450 monooxygenase. Plant Physiol. 1998, 118, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Siewers, V.; Smedsgaard, J.; Tudzynski, P. The P450 monooxygenase BcABA1 is essential for abscisic acid biosynthesis in Botrytis cinerea. Appl. Environ. Microbiol. 2004, 70, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Chepkirui, C.; Yuyama, K.; Wanga, L.; Decock, C.; Matasyoh, J.; Abraham, W.R.; Stadler, M. Microporenic acids A-G, biofilm inhibitors and antimicrobial agents from the basidiomycete Microporus sp. J. Nat. Prod. 2018, 81, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, K.; Chepkirui, C.; Wendt, L.; Fortkamp, D.; Stadler, M.; Abraham, W.R. Bioactive compounds produced by Hypoxylon fragiforme against Staphylococcus aureus biofilms. Microorganisms 2017, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Scognamiglio, M.; Donnarumma, G.; Fiorentino, A.; D′Abrosca, B. Recent advances in natural product-based anti-biofilm approaches to control infections. Mini Rev. Med. Chem. 2014, 14, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Narmani, A.; Pichai, S.; Palani, P.; Arzanlou, M.; Surup, F.; Stadler, M. Daldinia sacchari (Hypoxylaceae) from India produces the new cytochalasins Saccalasins A and B and belongs to the D. eschscholtzii species complex. Mycol. Prog. 2018, 1–11. [Google Scholar] [CrossRef]
- Surup, F.; Kuhnert, E.; Böhm, A.; Pendzialek, T.; Solga, D.; Wiebach, V.; Engler, H.; Berkessel, A.; Stadler, M.; Kalesse, M. The rickiols, 20-, 22-, and 24-membered macrolides from the ascomycete Hypoxylon rickii. Chem. Eur. J. 2018, 24, 2200–2213. [Google Scholar] [CrossRef] [PubMed]
- Sandargo, B.; Thongbai, B.; Stadler, M.; Surup, F. Cysteine-derived pleurotin congeners from the nematode-trapping basidiomycete Hohenbuehelia grisea. J. Nat. Prod. 2018, 81, 286–291. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds 1–3 are available from the authors. |
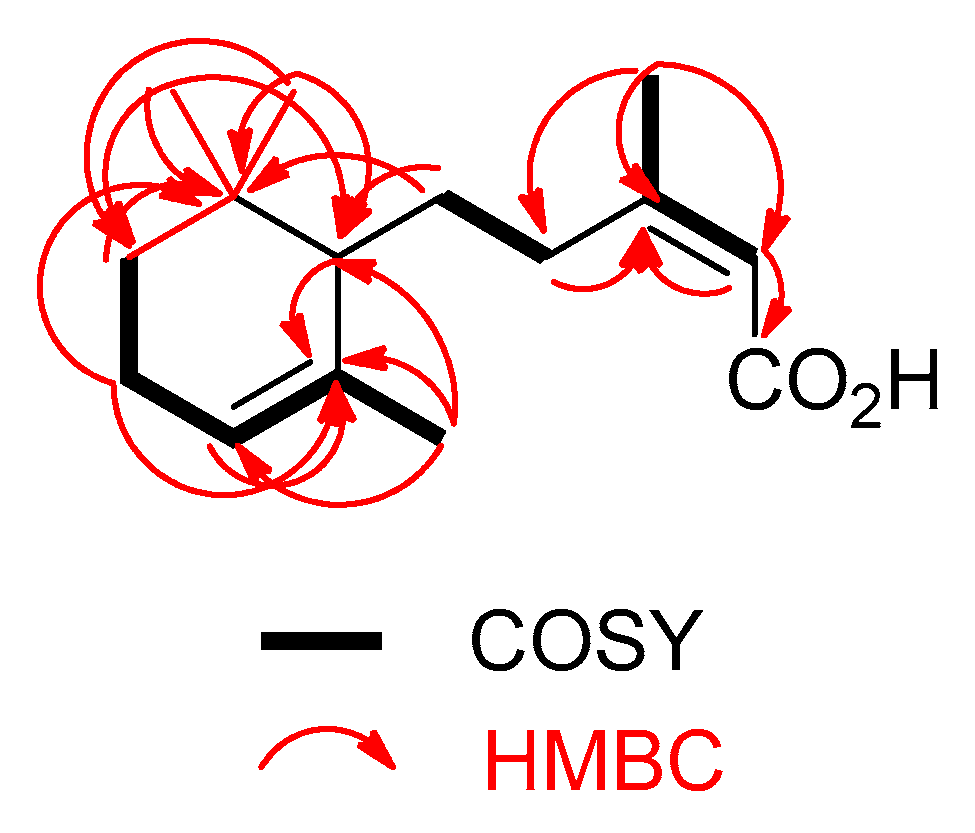

{kind=link}
{kind=link}
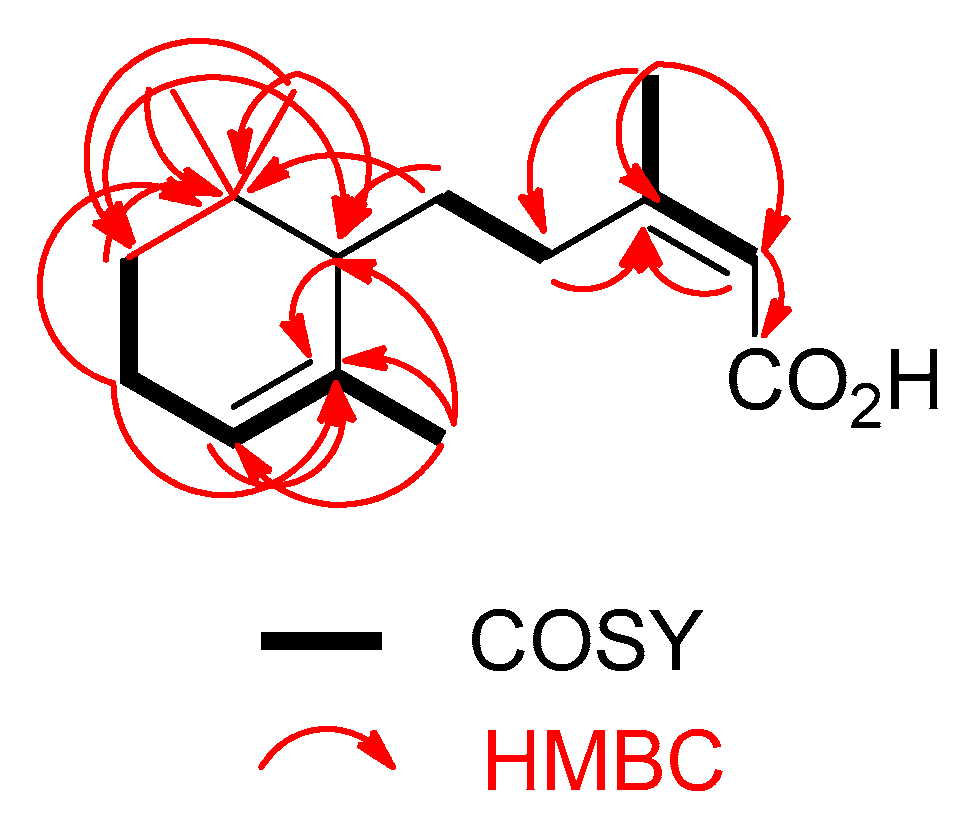{kind=link}
{kind=link}
| Position | δH 1 | δC 1 | HMBC 2 |
|---|---|---|---|
| 1 | - | 169.8 | - |
| 2 | 5.62, s | 117.2 | 1, 3, 4, 6 |
| 3 | - | 162.2 | - |
| 4a | 2.82 (ddd, 11.5, 10.5, 4.9) | 35.1 | 1, 2, 3, 6, 1′ |
| 4b | 2.48 (ddd, 11.5, 10.5, 4.9) | ||
| 5a | 1.61 3 | 30.7 | 3, 1′, 2′, 6′ |
| 5b | 1.48 3 | ||
| 6 | 1.90, s | 25.5 | 2, 3, 4, 6 |
| 1′ | 1.50 3 | 50.9 | 4, 2′, 5′, 6′, 7′, 8′, 9′ |
| 2′ | - | 137.7 | - |
| 3′ | 5.31, brs | 121.3 | 1′, 2′, 4′, 7′ |
| 4′ | 1.97 3 | 24.1 | 2′, 3′, 5′ |
| 5a′ | 1.48 3 | 32.8 | 1′, 2′, 5′, 8′, 9′ |
| 5b′ | 1.14 (dd, 7.1, 6.0) | ||
| 6′ | - | 33.7 | - |
| 7′ | 1.72, (d, 1.5) | 23.8 | 1′, 2′, 3′ |
| 8′ | 1.00, s | 28.2 | 1′, 5′, 6′, 9′ |
| 9′ | 0.89, s | 28.1 | 1′, 5′, 6′ 8′ |
| Strain | Compound 1 | Compound 2 | Positive Control (µg/mL) | |
|---|---|---|---|---|
| Bacteria | MIC (µg/mL) | |||
| Bacillus subtilis DSM 10 | 66.7 | - | 4.1 | Oxytetracyclin |
| Chromobacterium violaceum DSM 30191 | - | - | 0.4 | Oxytetracyclin |
| Escherichia coli DSM 1116 | - | - | 3.3 | Oxytetracyclin |
| Micrococcus luteus DSM 1790 | 66.7 | - | 0.2 | Oxytetracyclin |
| Mycobacterium smegmatis ATCC 700084 | - | - | 3.3 | Kanamycin |
| Pseudomonas aeruginosa PA14 | - | - | 1.7 | Gentamycin |
| Staphylococcus aureus DSM 346 | 66.7 | - | 3.3 | Oxytetracyclin |
| Fungi | ||||
| Candida albicans DSM 1665 | - | - | 66.7 | Nystatin |
| Mucor hiemalis DSM 2656 | 66.7 | - | 33.3 | Nystatin |
| Pichia anomala DSM 6766 | - | - | 33.3 | Nystatin |
| Rhodoturula glutinis DSM 10134 | - | - | 33.3 | Nystatin |
| Schizosaccharomyces pombe DSM 70572 | - | - | 33.3 | Nystatin |
| Cell lines | Cytotoxicity IC50 (µg/mL) | |||
| KB 3.1 HeLa | 14 | - | 5 × 10−4 | epothilone B |
| L929 | 27 | - | 1.1 × 10−3 | epothilone B |
| Test Organisms | Compound 1 | Compound 2 | Tetracycline |
|---|---|---|---|
| Biofilm Inhibition % | Biofilm Inhibition % | Biofilm Inhibition % | |
| Staphylococcus aureus DSM 1104 | 98% (256 µg/mL) | 36% (256 µg/mL) | 88% (100 µg/mL) |
| 83% (128 µg/mL) | 23% (128 µg/mL) | ||
| 73% (64 µg/mL) | - | ||
| 61% (32 µg/mL) | - | ||
| 34% (16 µg/mL) | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phukhamsakda, C.; Macabeo, A.P.G.; Yuyama, K.T.; Hyde, K.D.; Stadler, M. Biofilm Inhibitory Abscisic Acid Derivatives from the Plant-Associated Dothideomycete Fungus, Roussoella sp. Molecules 2018, 23, 2190. https://doi.org/10.3390/molecules23092190
Phukhamsakda C, Macabeo APG, Yuyama KT, Hyde KD, Stadler M. Biofilm Inhibitory Abscisic Acid Derivatives from the Plant-Associated Dothideomycete Fungus, Roussoella sp. Molecules. 2018; 23(9):2190. https://doi.org/10.3390/molecules23092190
Chicago/Turabian StylePhukhamsakda, Chayanard, Allan Patrick G. Macabeo, Kamila Tomoko Yuyama, Kevin David Hyde, and Marc Stadler. 2018. "Biofilm Inhibitory Abscisic Acid Derivatives from the Plant-Associated Dothideomycete Fungus, Roussoella sp." Molecules 23, no. 9: 2190. https://doi.org/10.3390/molecules23092190
APA StylePhukhamsakda, C., Macabeo, A. P. G., Yuyama, K. T., Hyde, K. D., & Stadler, M. (2018). Biofilm Inhibitory Abscisic Acid Derivatives from the Plant-Associated Dothideomycete Fungus, Roussoella sp. Molecules, 23(9), 2190. https://doi.org/10.3390/molecules23092190