A Three Component Synthetic Vaccine Containing a β-Mannan T-Cell Peptide Epitope and a β-Glucan Dendritic Cell Ligand

 ,
,
Abstract
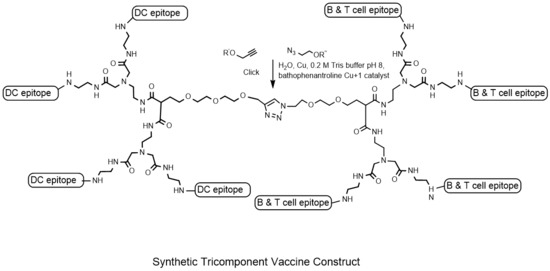
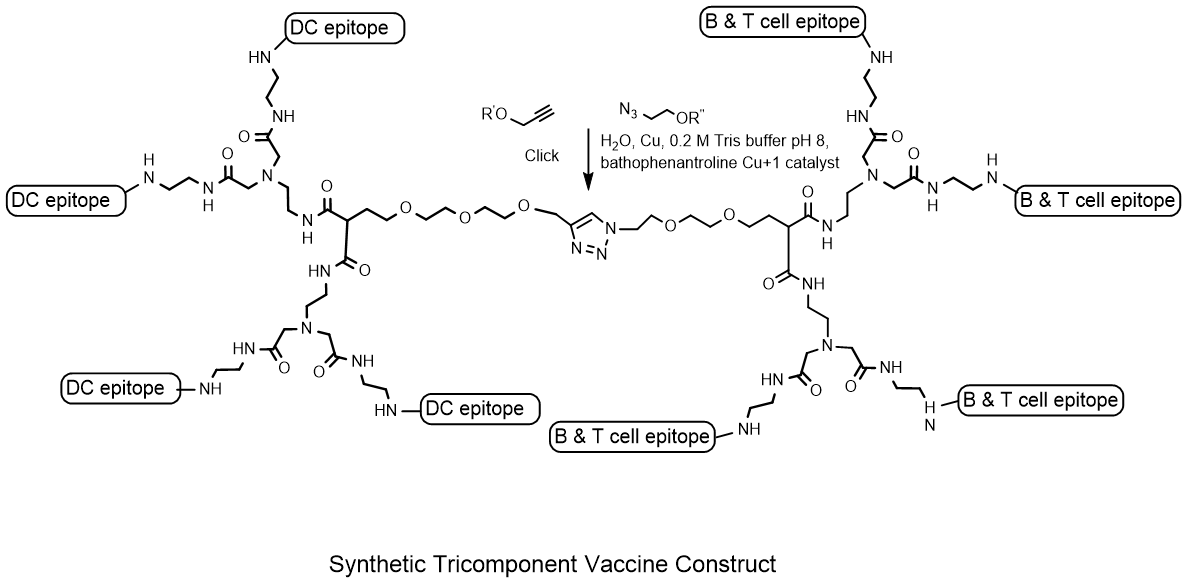{kind=link}
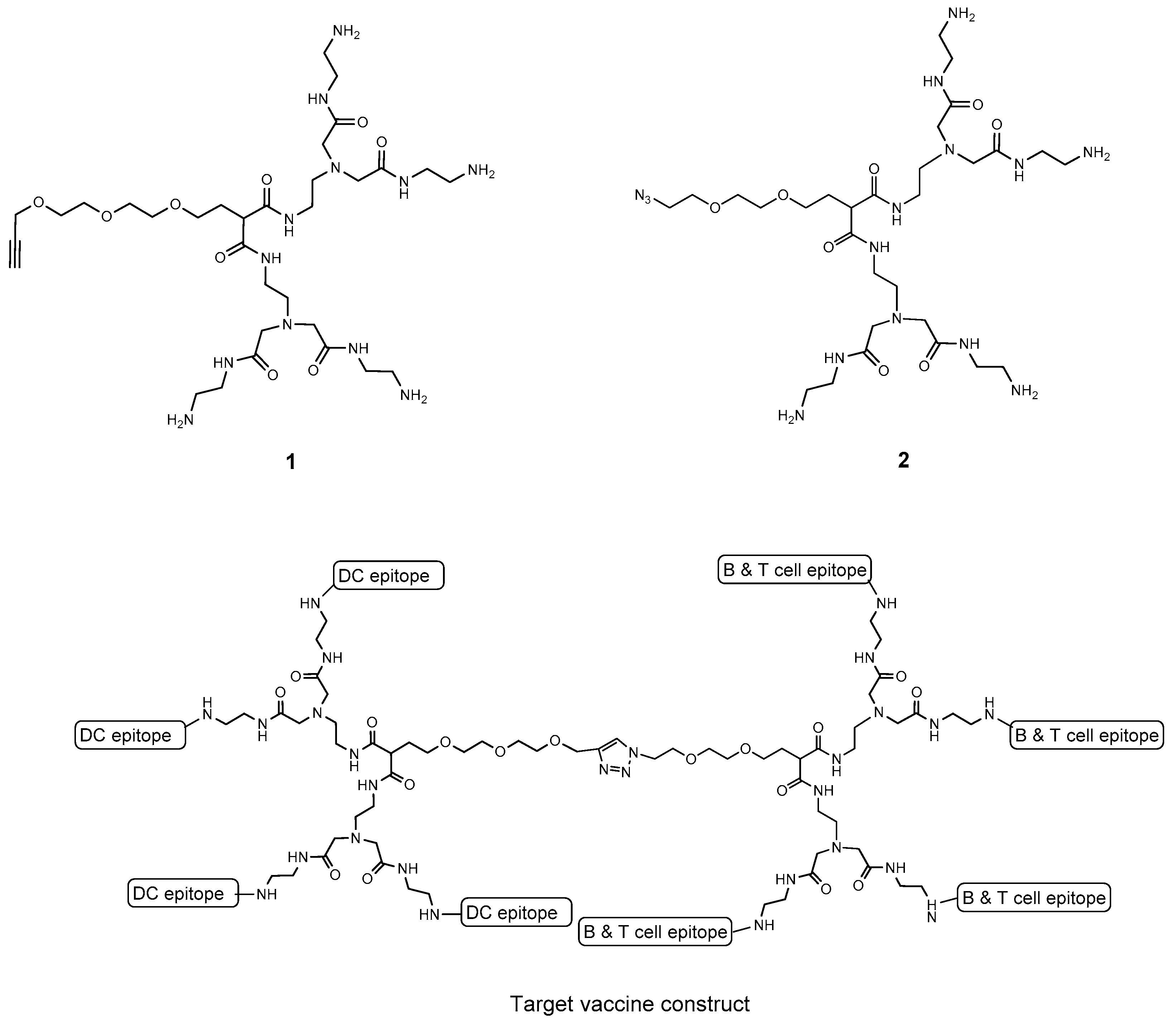{kind=link}
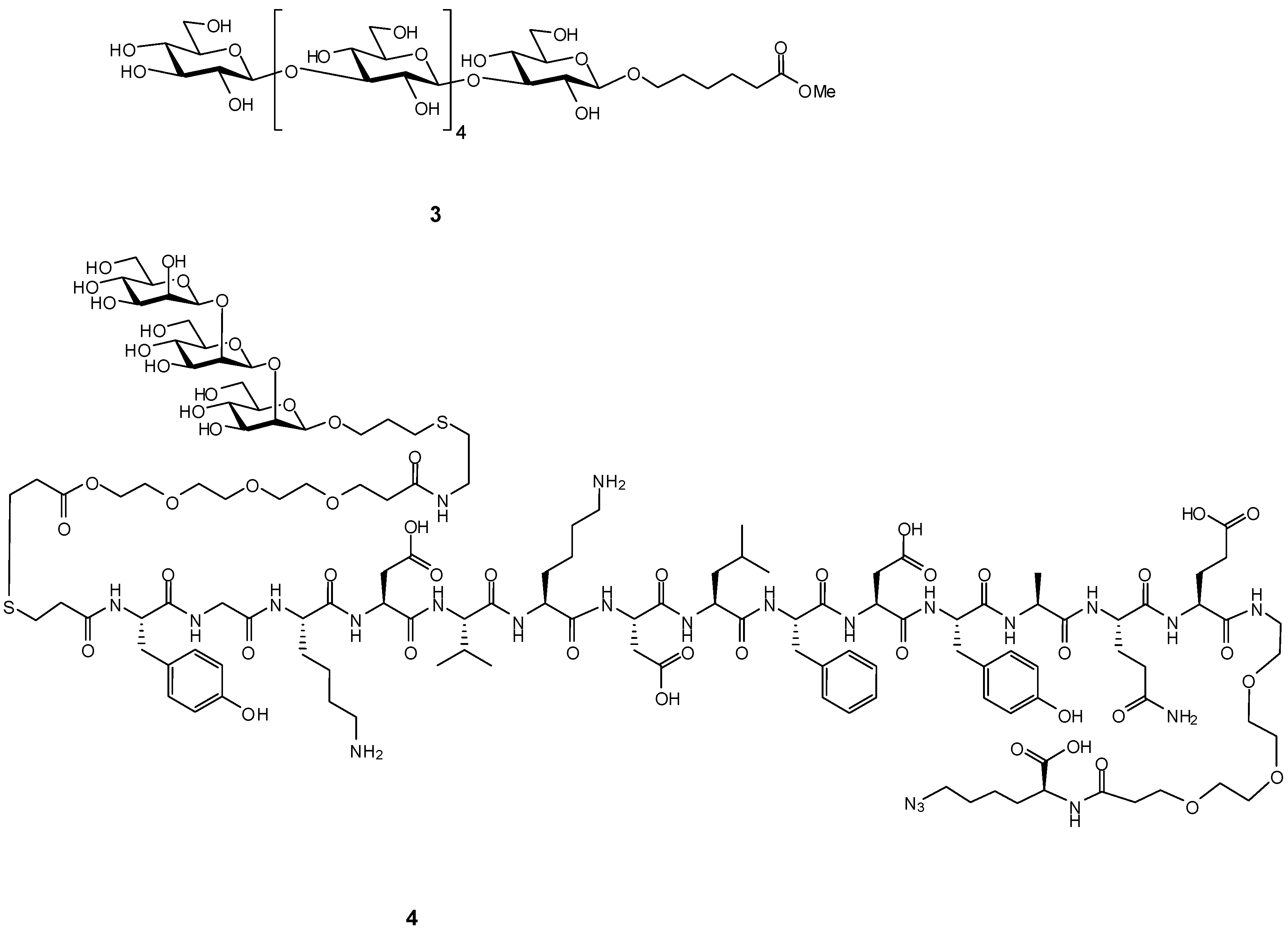{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
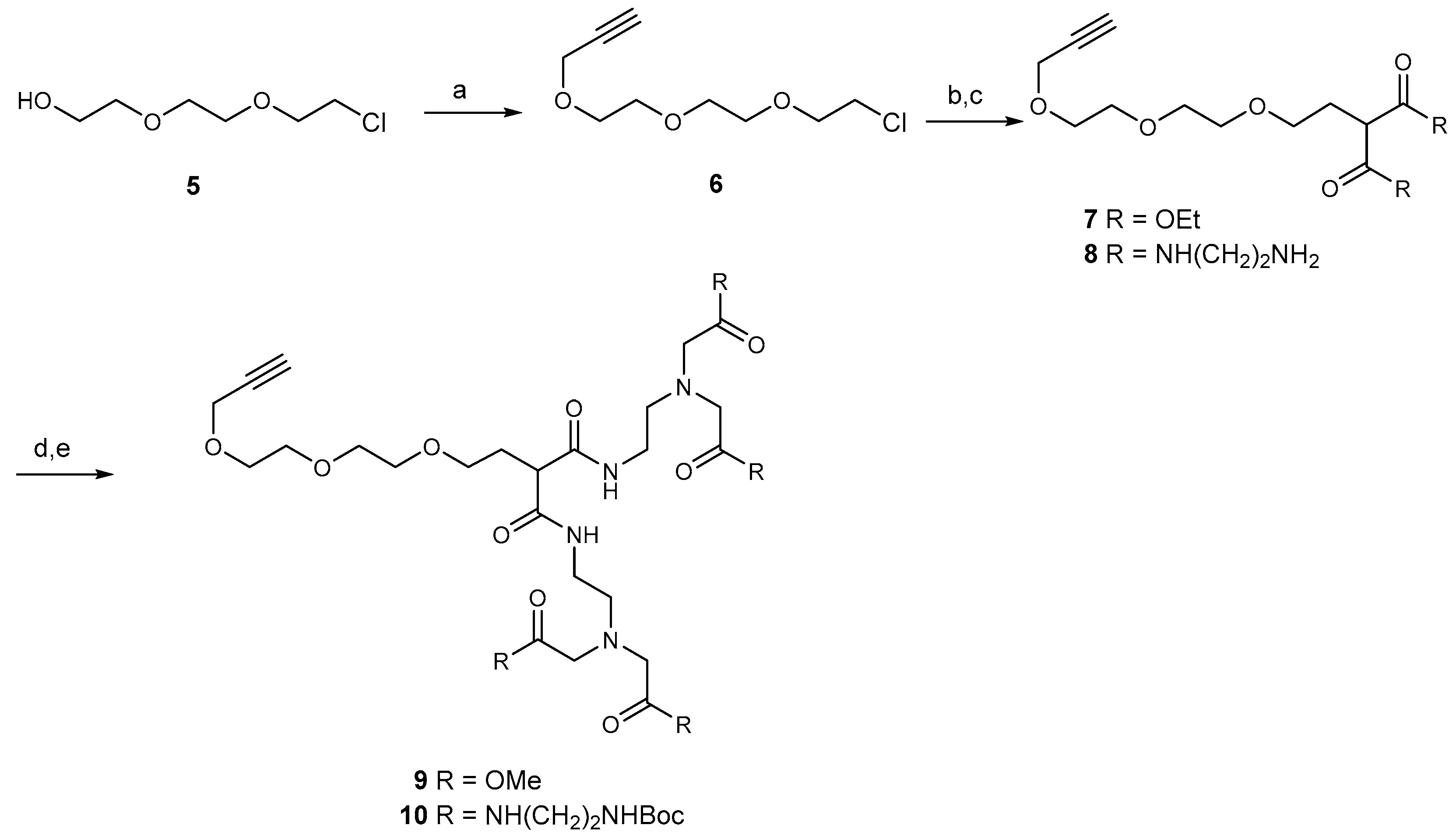
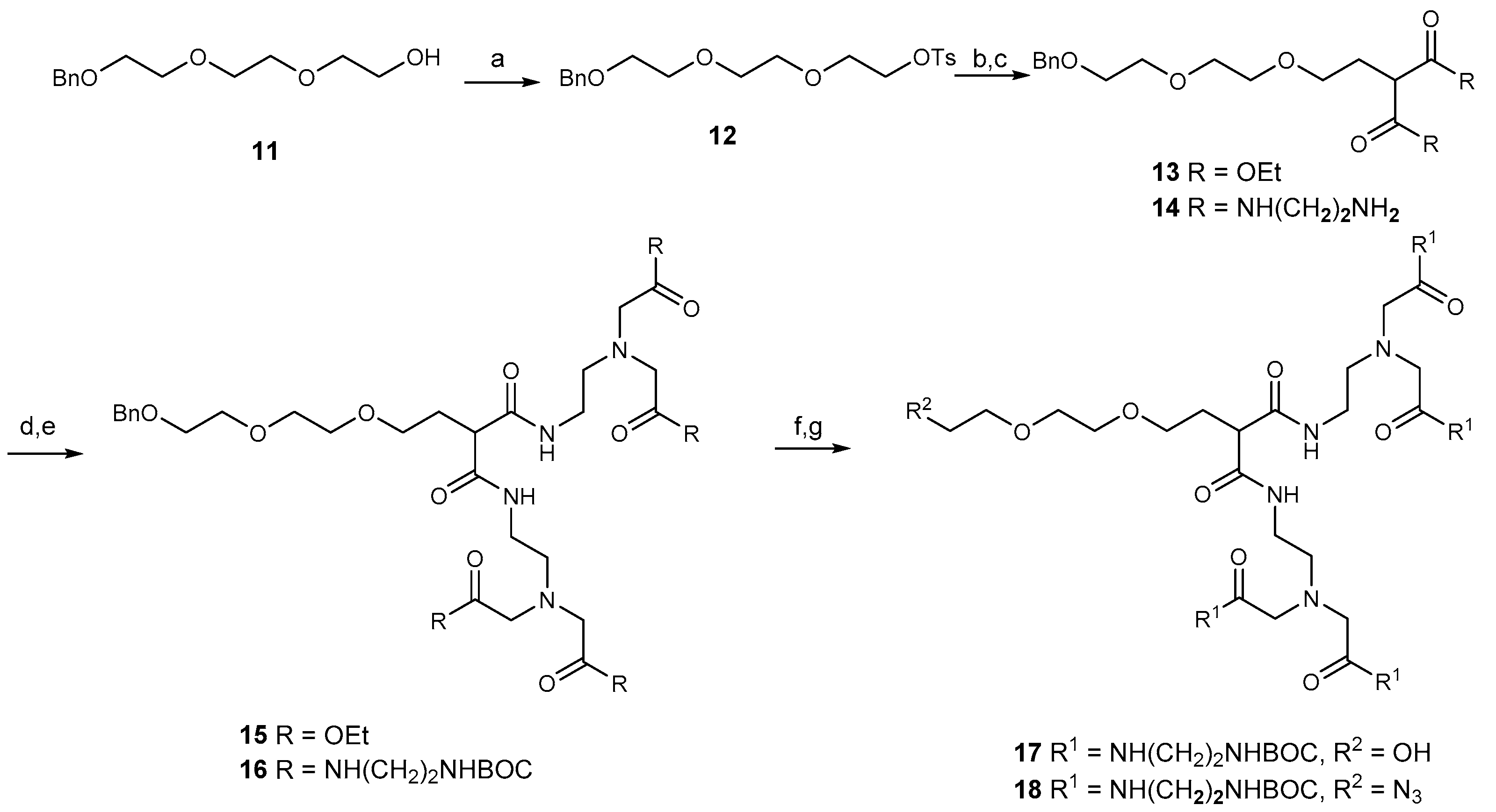
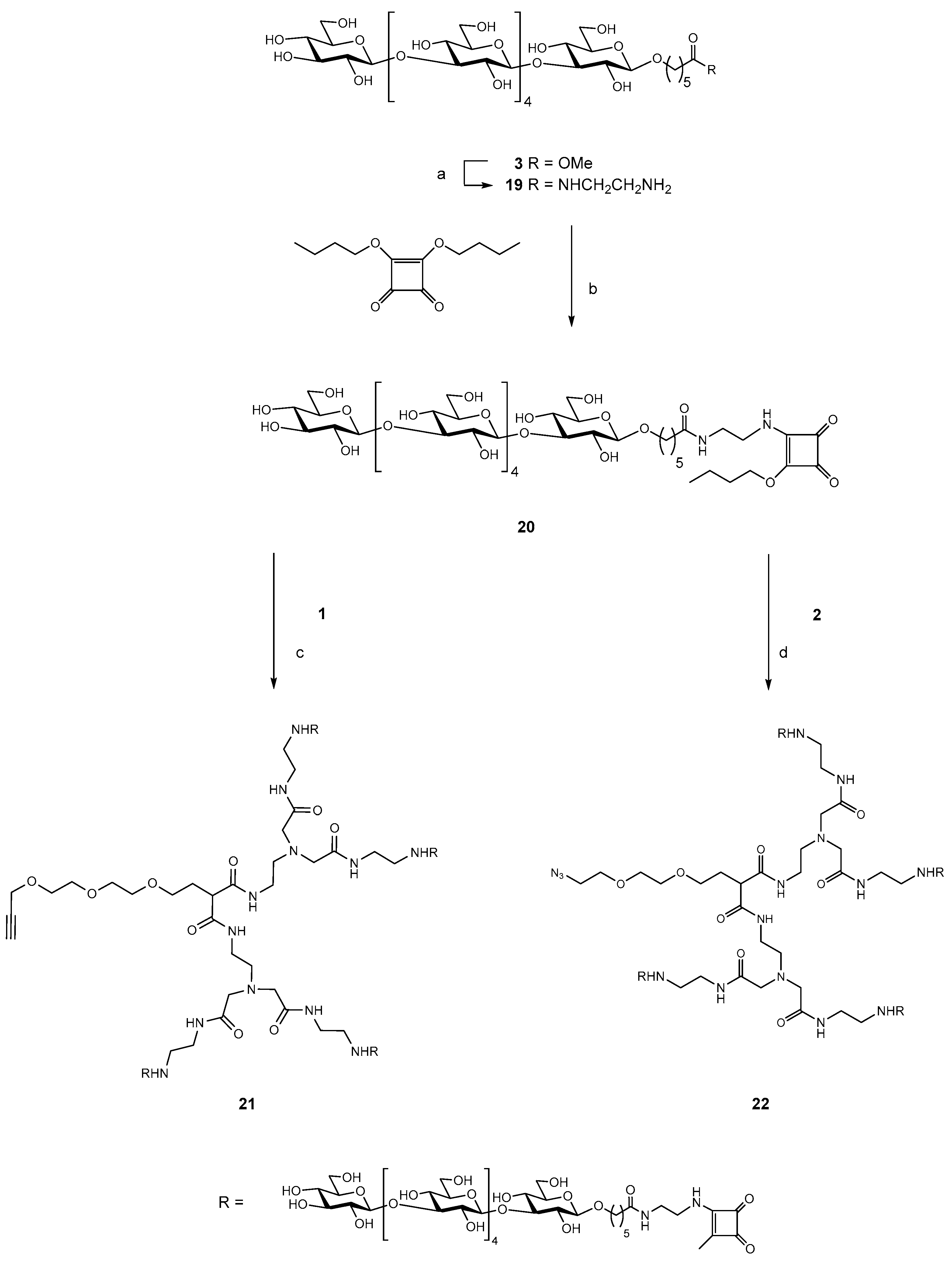
2.1. Synthesis of Dendrimers
2.2. Synthesis of the Tetravalent Dectin-1 Ligand
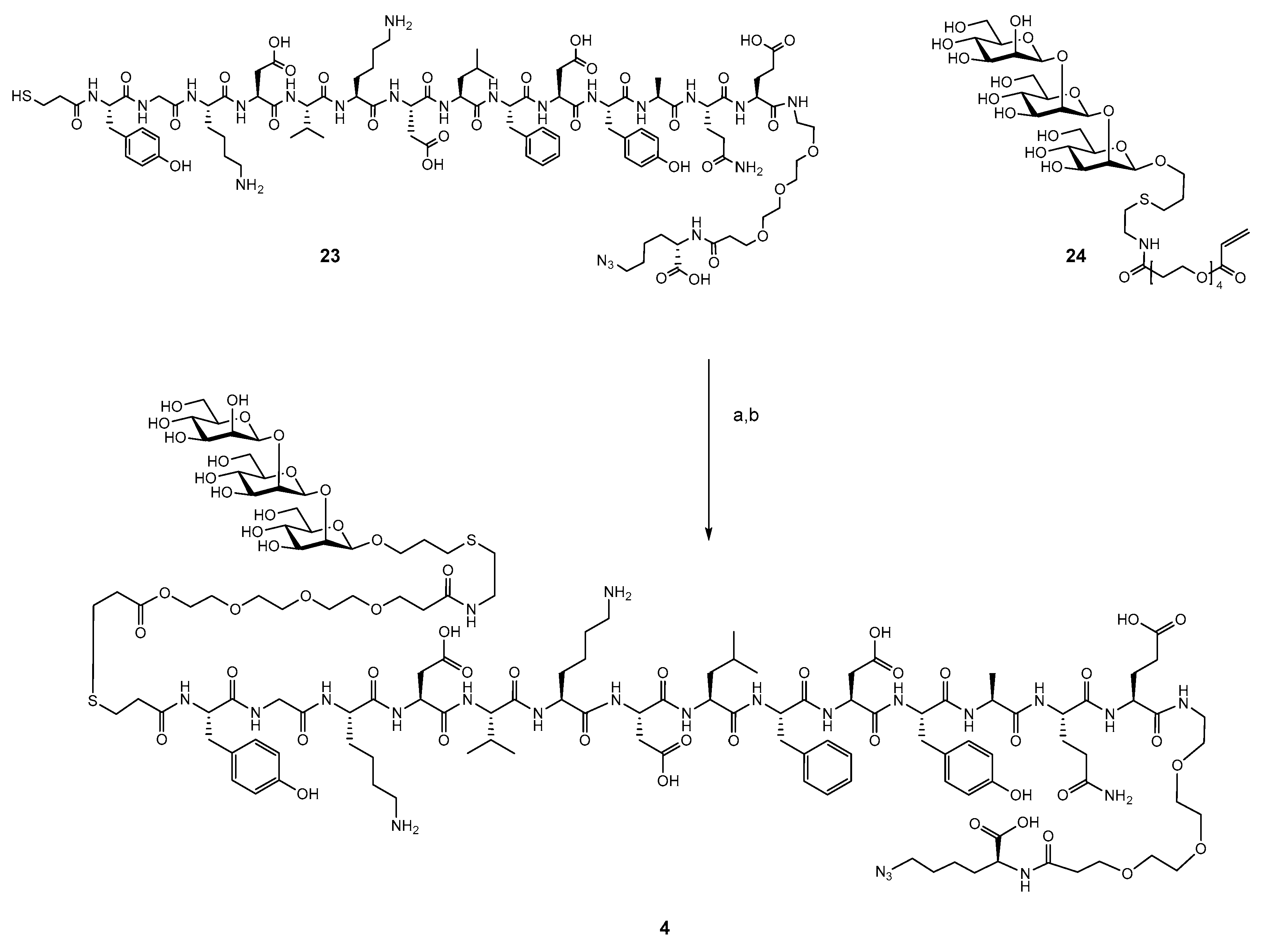
2.3. Synthesis of B- and T-Cell Epitope, β-Mannotriose Fba Glycopeptide
2.4. Attempts to Conjugate Glycopeptide 4 to Dendrimer
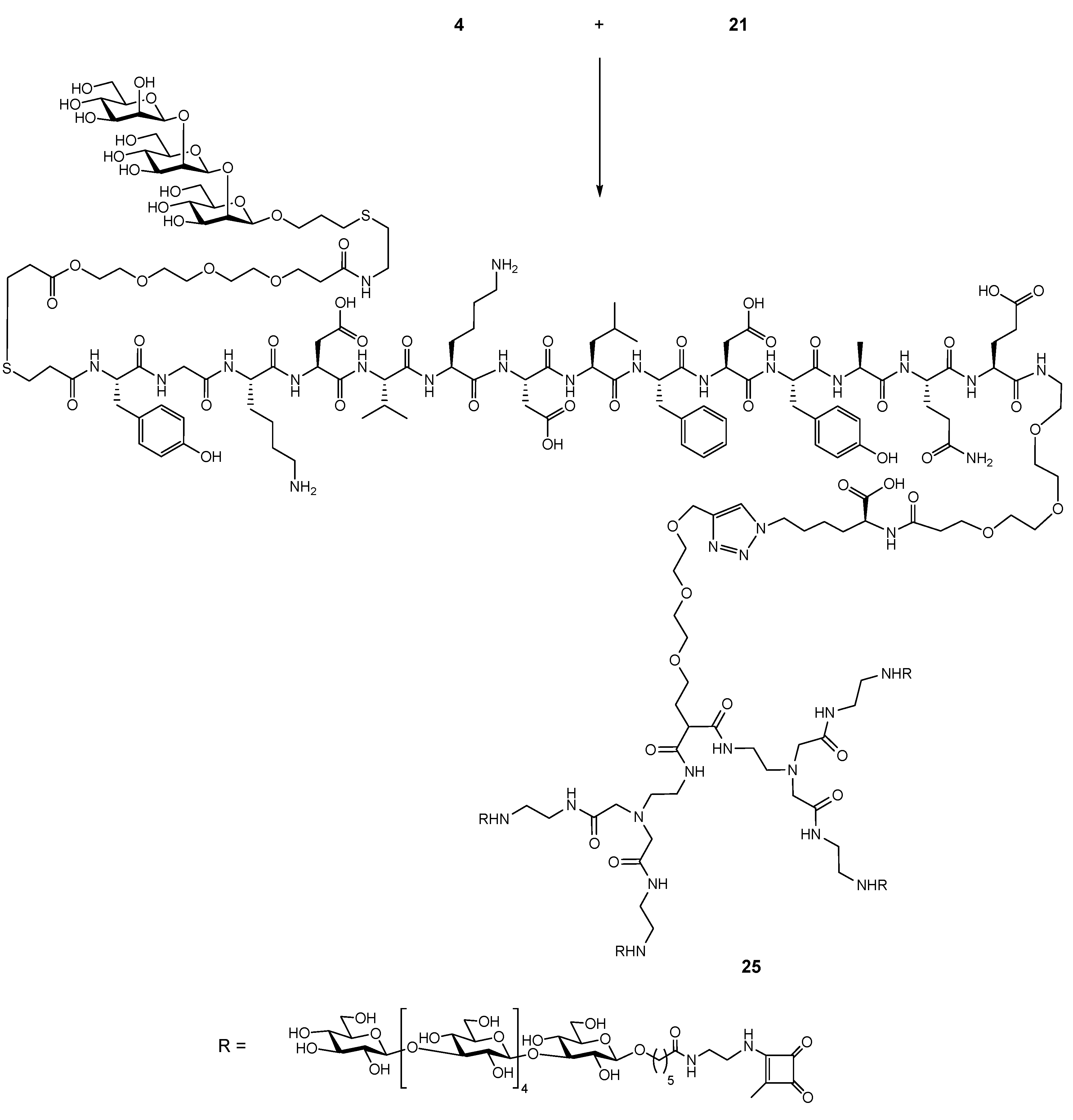
2.5. Conjugation of Glycopeptide 4 with the Tetravalent β-Glucan Dendrimer 21
2.6. Conjugation of β-Glucan Dendrimer 22 to Activated Ovalbumin
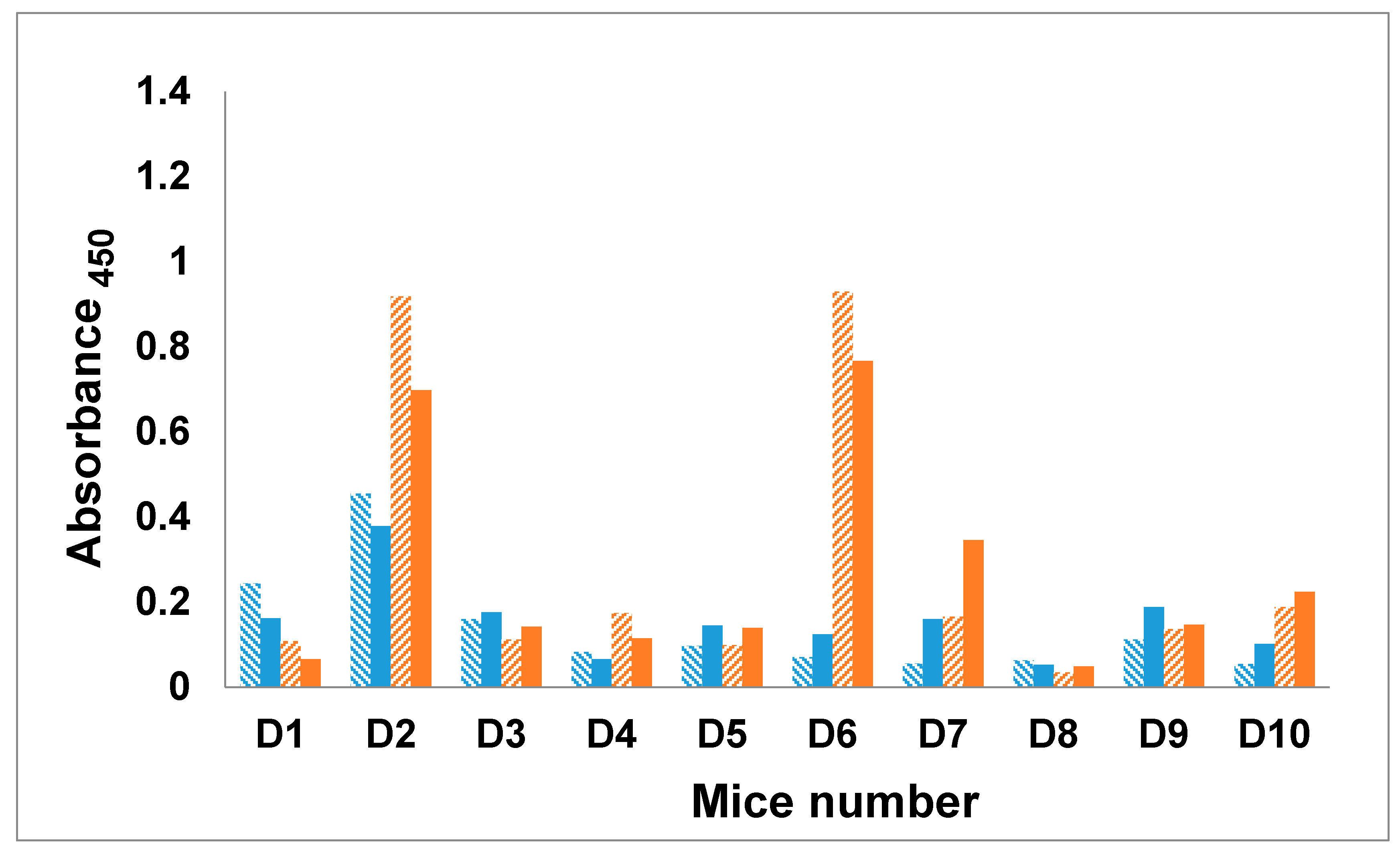
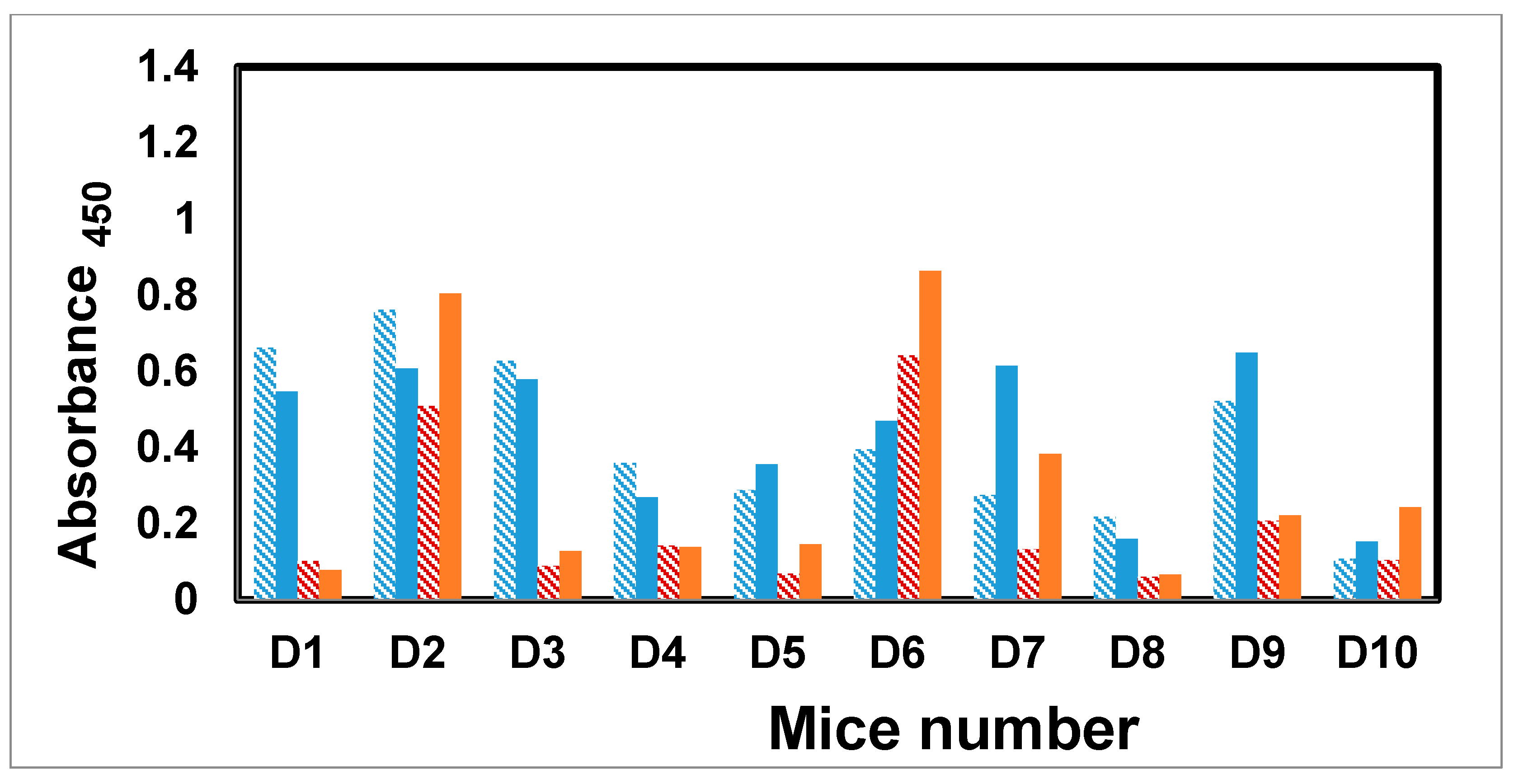
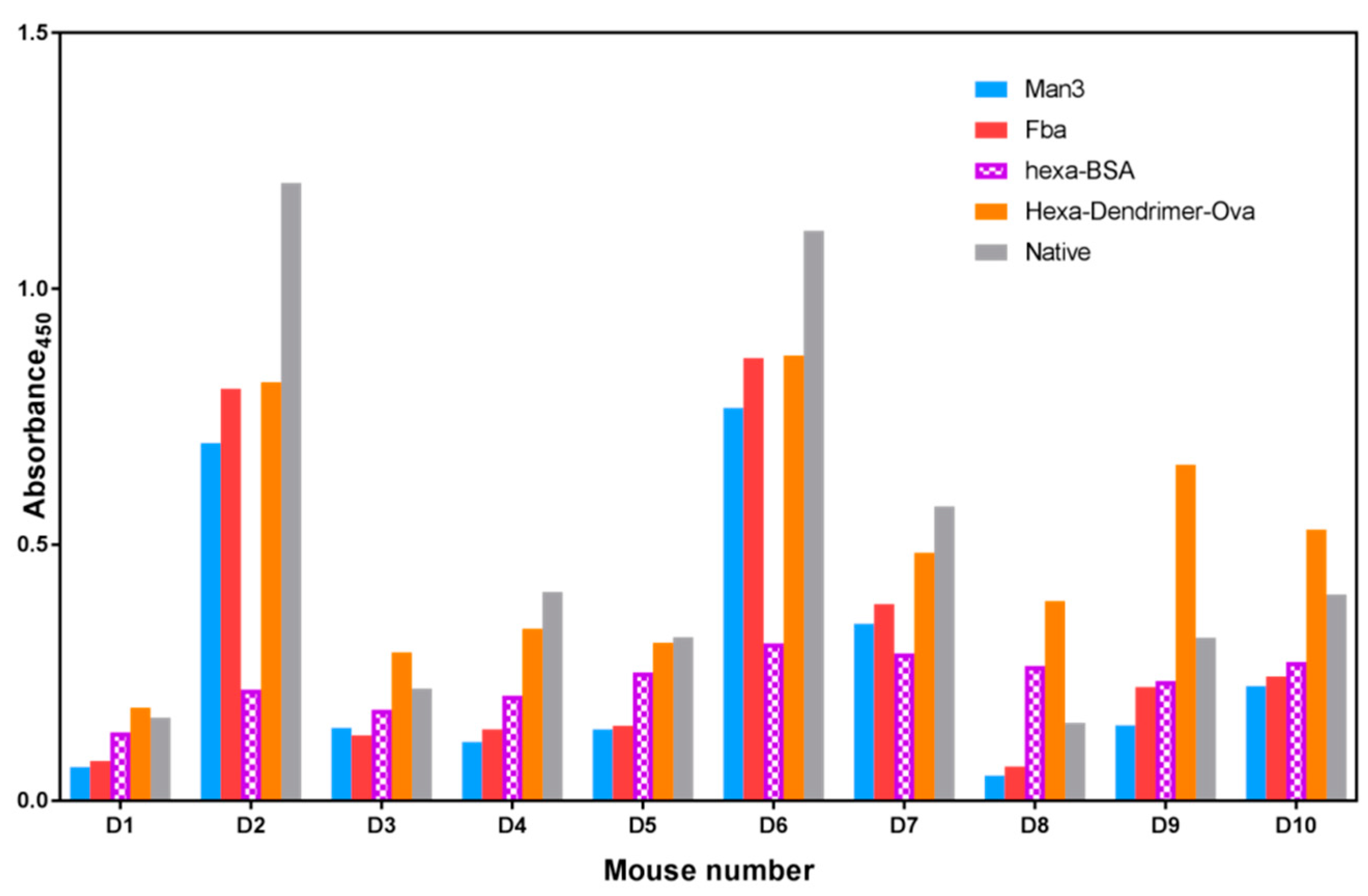
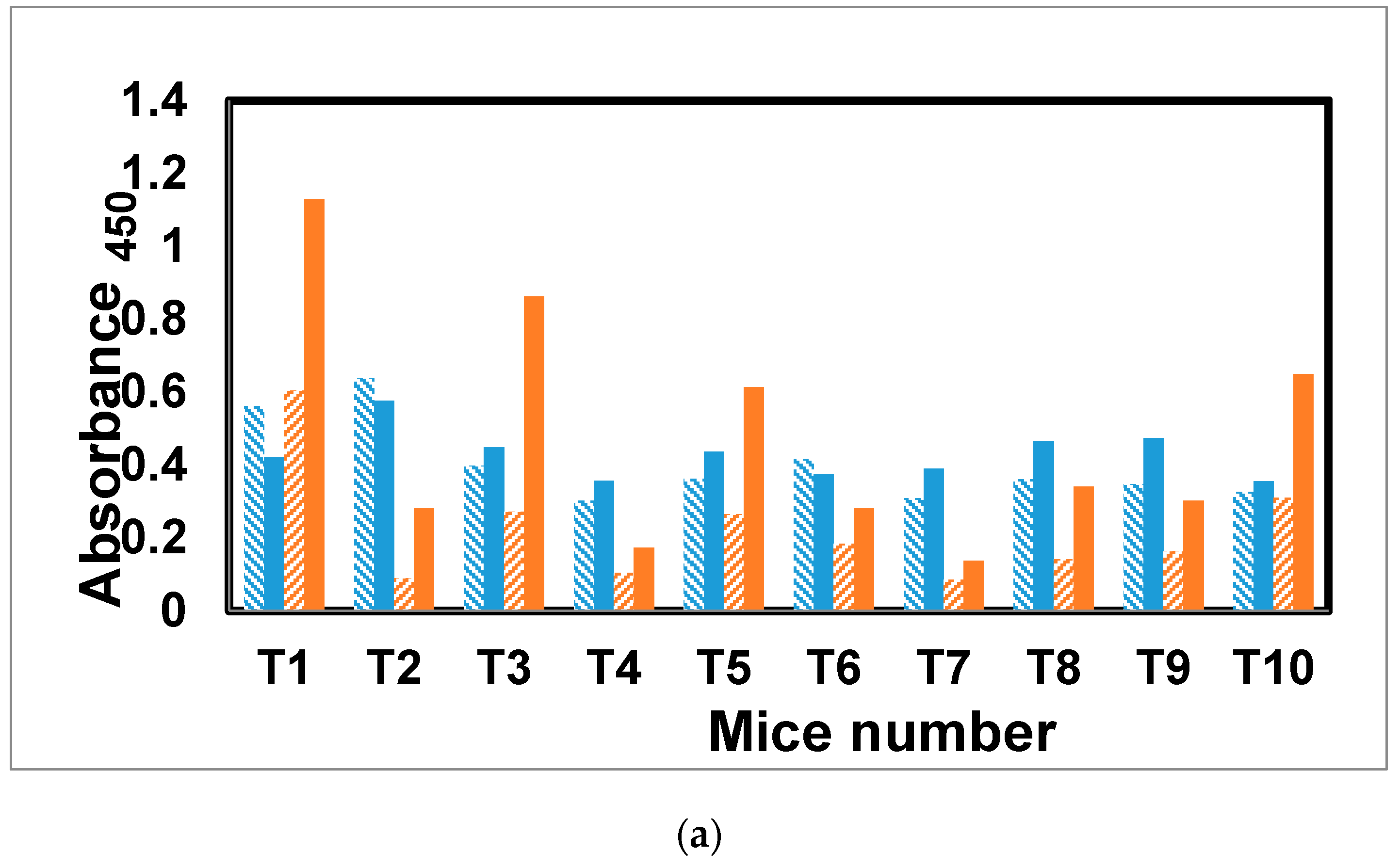
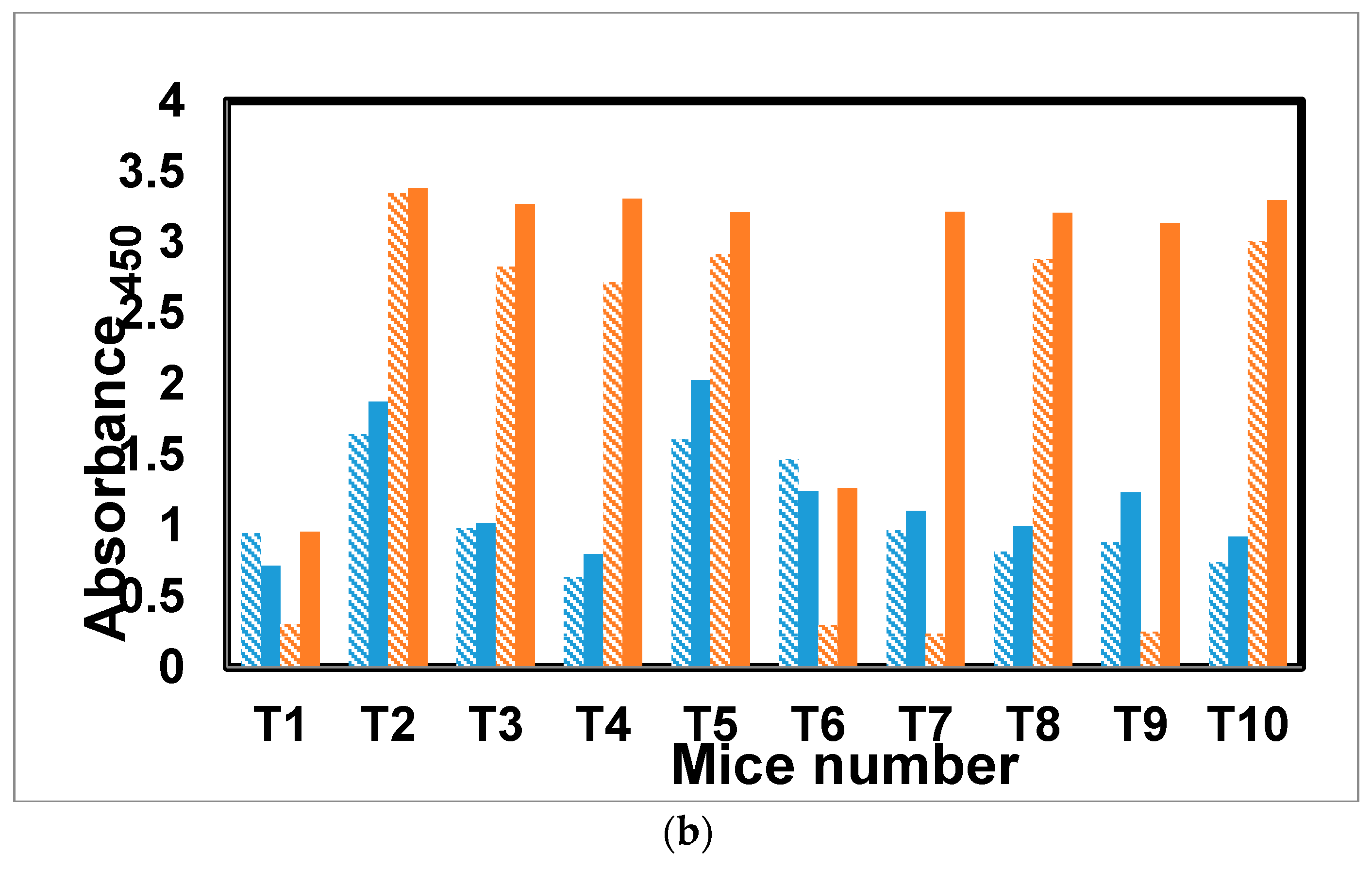
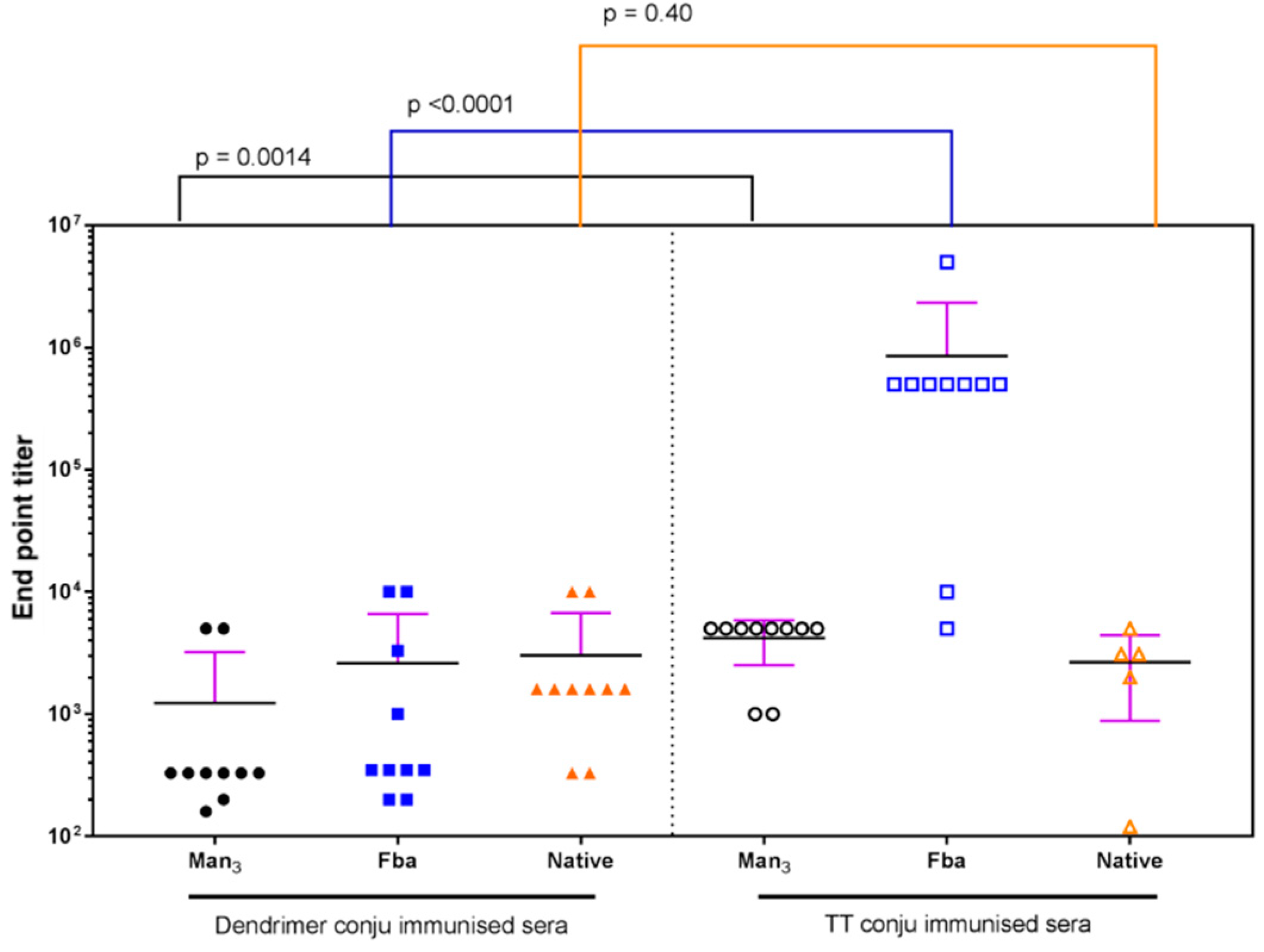
2.7. Immunization of Mice with the Trivalent Antigen 25 and Assay of Antibody Profiles
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Antigens
4.3. Vaccine Formulation
4.4. Immunization
4.5. Serum Processing
4.6. Immunoassays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maiden, M.C. The impact of protein-conjugate polysaccharide vaccines: An endgame for meningitis? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120147. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Perrett, K.P.; Beverley, P.C. Maintaining protection against invasive bacteria with protein-polysaccharide conjugate vaccines. Nat. Rev. Immunol. 2009, 9, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Blanchard-Rohner, G.; Pollard, A.J. Long-term protection after immunization with protein-polysaccharide conjugate vaccines in infancy. Expert Rev. Vaccines 2011, 5, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Bonten, M.J.M.; Huijts, S.M.; Bolkenbaas, M.; Webber, C.; Patterson, S.; Gault, S.; van Werkhoven, C.H.; van Deursen, A.M.M.; Sanders, E.A.M.; Verheij, T.J.M.; et al. Polysaccharide conjugate vaccine against pneumococcal pneumonia in adults. N. Engl. J. Med. 2015, 372, 1114–1125. [Google Scholar] [CrossRef] [PubMed]
- Shiramoto, M.; Hanada, R.; Juergens, C.; Shoji, Y.; Yoshida, M.; Ballan, B.; Cooper, D.; Gruber, W.C.; Scott, D.A.; Schmoele-Thoma, B. Immunogenicity and safety of the 13-valent pneumococcal conjugate vaccine compared to the 23-valent pneumococcal polysaccharide vaccine in elderly Japanese adults. Hum. Vaccines Immunother. 2015, 11, 2198–2206. [Google Scholar] [CrossRef] [PubMed]
- Avci, F.Y.; Li, X.; Tsuji, M.; Kasper, D.L. A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design. Nat. Med. 2011, 17, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Avci, F.Y. Novel strategies for development of next-generation glycoconjugate vaccines. Curr. Top. Med. Chem. 2013, 13, 2535–2540. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M. Dendritic cells: Versatile controllers of the immune system. Nat. Med. 2007, 13, vii–xi. [Google Scholar] [CrossRef] [PubMed]
- Tacken, P.J.; Figdor, C.G. Targeted antigen delivery and activation of dendritic cells in vivo: Steps towards cost effective vaccines. Semin. Immunol. 2011, 23, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.W.; Thompson, C.; Reid, D.M.; Wong, S.Y.C.; Tough, D.F. Preferential Induction of CD4+ T Cell Responses through in vivo Targeting of Antigen to Dendritic Cell-Associated C-Type Lectin-1. J. Immunol. 2006, 177, 2276–2284. [Google Scholar] [CrossRef] [PubMed]
- Miyata, T.; Harakuni, T.; Tsuboi, T.; Sattabongkot, J.; Ikehara, A.; Tachibana, M.; Torii, M.; Matsuzaki, G.; Arakawa, T. Tricomponent Immunopotentiating System as a Novel Molecular Design Strategy for Malaria Vaccine Development. Infect. Immun. 2011, 79, 42604275. [Google Scholar] [CrossRef] [PubMed]
- Unger, W.W.J.; van Kooyk, Y. ‘Dressed for success’ C-type lectin receptors for the delivery of glyco-vaccines to dendritic cells. Curr. Opin. Immunol. 2011, 23, 131–137. [Google Scholar] [CrossRef] [PubMed]
- De Koker, S.; Lambrecht, B.N.; Willart, M.A.; van Kooyk, Y.; Grooten, J.; Vervaet, C.; Remona, J.P.; De Geest, B.G. Designing polymeric particles for antigen delivery. Chem. Soc. Rev. 2011, 40, 320–339. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, S.J.; García-Vallejo, J.J.; van Kooyk, Y. Dendritic cells and C-type lectin receptors: Coupling innate to adaptive immune responses. Immunol. Cell Biol. 2008, 86, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Van Kooyk, Y.; Rabinovich, G.A. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat. Immunol. 2008, 9, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Palma, A.S.; Feizi, T.; Zhang, Y.; Stoll, M.S.; Lawson, A.M.; Díaz-Rodríguez, E.; Campanero-Rhodes, M.A.; Costa, J.; Gordon, S.; Brown, G.D.; et al. Ligands for the β-Glucan Receptor, Dectin-1, Assigned Using “Designer” Microarrays of Oligosaccharide Probes (Neoglycolipids) Generated from Glucan Polysaccharides. J. Biol. Chem. 2006, 281, 5771–5779. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.L.; Rice, P.J.; Graves, B.; Ensley, H.E.; Yu, H.; Brown, G.D.; Gordon, S.; Monteiro, M.A.; Papp-Szabo, E.; Lowman, D.W.; et al. Differential high-affinity interaction of dectin-1 with natural or synthetic glucans is dependent upon primary structure and is influenced by polymer chain length and side-chain branching. J. Pharmacol. Exp. Ther. 2008, 325, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Donadei, A.; Gallorini, S.; Berti, F.; O’Hagan, D.T.; Adamo, R.; Baudner, B.C. Rational Design of Adjuvant for Skin Delivery: Conjugation of Synthetic β-Glucan Dectin-1 Agonist to Protein Antigen. Mol. Pharm. 2015, 12, 1662–1672. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Dziadek, S.; Bundle, D.R.; Cutler, J. Synthetic glycopeptide vaccines combining β-mannan and peptide epitopes induce protection against candidiasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13526–13531. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Cartmell, J.; Bailey, J.J.; Dziadek, S.; Bundle, D.R.; Cutler, J.E. Self-adjuvanting Glycopeptide Conjugate Vaccine against Disseminated Candidiasis. PLoS ONE 2012, 7, e35106. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, T.; Fitieh, A.; St. Pierre, J.; Ostergaard, H.L.; Bundle, D.R.; Touret, N. Enhanced Immunogenicity of a tricomponent mannan tetanus toxoid conjugate vaccine targeted to DCs via Dectin-1 by incorporating β-glucan. J. Immunol. 2013, 190, 4116–4128. [Google Scholar] [CrossRef] [PubMed]
- Elsaidi, H.; Paszkiewicz, E.; Bundle, D.R. Synthesis of a 1,3 β-glucan hexasaccharide designed to target vaccines to the dendritic cell receptor, Dectin-1. Carbohydr. Res. 2015, 408, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Kawai, T.; Adachi, Y.; Hanashima, S.; Yamaguchi, Y.; Ohno, N.; Takahashi, T. Synthesis of β-(1,3) oligoglucans exhibiting a Dectin-1 binding affinity and their biological evaluation. Bioorg. Med. Chem. 2012, 20, 3898–3914. [Google Scholar] [CrossRef] [PubMed]
- Hanashima, S.; Ikeda, A.; Tanaka, H.; Adachi, Y.; Ohno, N.; Takahashi, T.; Yamaguchi, Y. NMR study of short β(1,3)-glucans provides insights into the structure and interaction with Dectin-1. Glycoconj. J. 2014, 31, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Bouzide, A.; Sauve, G. Highly selective silver(I) oxide mediated monoprotection of symmetrical diols. Tetrahedron Lett. 1997, 38, 5945–5948. [Google Scholar] [CrossRef]
- Hou, S.J.; Saksena, R.; Kovac, P. Preparation of glycoconjugates by dialkyl squarate chemistry revisited. Carbohydr. Res. 2009, 343, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Guiard, J.; Paszkiewicz, E.; Sadowska, J.; Bundle, D.R. Design and synthesis of a universal antigen to detect brucellosis. Angew. Chem. Int. Ed. 2013, 52, 7181–7185. [Google Scholar] [CrossRef] [PubMed]
- Dziadek, S.; Jacques, S.; Bundle, D.R. A novel linker methodology for the synthesis of tailored conjugate vaccines composed of complex carbohydrate antigens and specific TH-cell peptide epitopes. Chem. Eur. J. 2008, 14, 5908–5917. [Google Scholar] [CrossRef] [PubMed]
- Cartmell, J.; Paszkiewicz, E.; Dziadek, S.; Tam, P.-H.; Luu, T.; Sarkar, S.; Lipinski, T.; Bundle, D.R. Synthesis of antifungal vaccines by conjugation of β-1,2 trimannosides with T-cell peptides and covalent anchoring of neoglycopeptide to tetanus toxoid. Carbohydr. Res. 2015, 403, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.C.; Finn, M.G.; Sharpless, K.B. Click Chemistry: Diverse Chemical Function from a Few Good Reactions. Angew. Chem. Int. Ed. 2001, 40, 2004–2021. [Google Scholar] [CrossRef]
- Rostovtsev, V.V.; Green, L.G.; Fokin, V.V.; Sharpless, K.B. A Stepwise Huisgen Cycloaddition Process: Copper(I)-Catalyzed Regioselective “Ligation” of Azides and Terminal Alkynes. Angew. Chem. Int. Ed. 2002, 41, 2596–2599. [Google Scholar] [CrossRef]
- Lipinski, T.; Kitov, P.I.; Szpacenko, A.; Paszkiewicz, E.; Bundle, D.R. Synthesis and Immunogenicity of a Glycopolymer Conjugate. Bioconjug. Chem. 2011, 22, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Sen Gupta, S.; Kuzelka, J.; Singh, P.; Lewis, W.G.; Manchester, M.; Finn, M.G. Accelerated bioorthogonal conjugation: A practical method for the ligation of diverse functional molecules to a polyvalent virus scaffold. Bioconjug. Chem. 2005, 16, 1572–1579. [Google Scholar] [CrossRef] [PubMed]
- Kitov, P.I.; Kotsuchibashi, Y.; Paszkiewicz, E.; Wilhelm, D.; Narain, R.; Bundle, D.R. Poly(n-vinyl-2-pyrrolidone-co-vinyl alcohol) as a versatile amphiphilic polymeric scaffold for multivalent probes. Org. Lett. 2013, 15, 5190–5193. [Google Scholar] [CrossRef] [PubMed]
- Bundle, D.R.; Tam, P.-H.; Tran, H.-A.; Paszkiewicz, E.; Cartmell, J.; Sadowska, J.M.; Sarkar, S.; Joe, M.; Kitov, P.I. Oligosaccharides and peptide displayed on an amphiphilic polymer enable solid phase assay of hapten specific antibodies. Bioconjug. Chem. 2014, 25, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Bishop, C.T.; Blank, F.; Gardner, P.E. The cell wall polysaccharides of Candida albicans: Glucan, mannan, and chitin. Can. J. Chem. 1960, 38, 869–881. [Google Scholar] [CrossRef]
- Sayeed, M.A.; Bufano, M.K.; Xu, P.; Eckhoff, G.; Charles, R.C.; Alam, M.M.; Sultana, T.; Rashu, M.R.; Berger, A.; Gonzalez-Escobedo, G.; et al. A Cholera Conjugate Vaccine Containing O-specific Polysaccharide (OSP) of V. cholerae O1 Inaba and Recombinant Fragment of Tetanus Toxin Heavy Chain (OSP:rTTHc) Induces Serum, Memory and Lamina Proprial Responses against OSP and Is Protective in Mice. PLoS Negl. Trop. Dis. 2015, 9, e0003881. [Google Scholar] [CrossRef] [PubMed]
- Mawas, F.; Niggemann, J.; Jones, C.; Corbel, M.J.; Kamerling, J.P.; Vliegenthart, J.F.G. Immunogenicity in a Mouse Model of a Conjugate Vaccine Made with a Synthetic Single Repeating Unit of Type 14 Pneumococcal Polysaccharide Coupled to CRM197. Infect. Immun. 2002, 70, 5107–5114. [Google Scholar] [CrossRef] [PubMed]
- Torosantucci, A.; Bromuro, C.; Chiani, P.; de Bernardis, F.; Berti, F.; Galli, C.; Norelli, F.; Bellucci, C.; Polonelli, L.; Costantino, P.; et al. A novel glyco-conjugate vaccine against fungal pathogens. J. Exp. Med. 2005, 202, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Bromuro, C.; Romano, M.; Chiani, P.; Berti, F.; Tontini, M.; Proietti, D.; Mori, E.; Torosantucci, A.; Costantino, P.; Rappuoli, R.; et al. Beta-glucan-CRM197 conjugates as candidates antifungal vaccines. Vaccine 2010, 28, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Brune, K.D.; Buldun, C.M.; Li, Y.; Taylor, I.J.; Brod, F.; Biswas, S.; Howarth, M. Dual Plug-and-Display Synthetic Assembly Using Orthogonal Reactive Proteins for Twin Antigen Immunization. Bioconjug. Chem. 2017, 28, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- Negahdaripour, M.; Golkar, N.; Hajighahramani, N.; Kianpour, S.; Nezafat, N.; Ghasemi, Y. Harnessing self-assembled peptide nanoparticles in epitope vaccine design. Biotechnol. Adv. 2017, 35, 575–596. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |














© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bundle, D.R.; Paszkiewicz, E.; Elsaidi, H.R.H.; Mandal, S.S.; Sarkar, S. A Three Component Synthetic Vaccine Containing a β-Mannan T-Cell Peptide Epitope and a β-Glucan Dendritic Cell Ligand. Molecules 2018, 23, 1961. https://doi.org/10.3390/molecules23081961
Bundle DR, Paszkiewicz E, Elsaidi HRH, Mandal SS, Sarkar S. A Three Component Synthetic Vaccine Containing a β-Mannan T-Cell Peptide Epitope and a β-Glucan Dendritic Cell Ligand. Molecules. 2018; 23(8):1961. https://doi.org/10.3390/molecules23081961
Chicago/Turabian StyleBundle, David R., Eugenia Paszkiewicz, Hassan R. H. Elsaidi, Satadru Sekhar Mandal, and Susmita Sarkar. 2018. "A Three Component Synthetic Vaccine Containing a β-Mannan T-Cell Peptide Epitope and a β-Glucan Dendritic Cell Ligand" Molecules 23, no. 8: 1961. https://doi.org/10.3390/molecules23081961
APA StyleBundle, D. R., Paszkiewicz, E., Elsaidi, H. R. H., Mandal, S. S., & Sarkar, S. (2018). A Three Component Synthetic Vaccine Containing a β-Mannan T-Cell Peptide Epitope and a β-Glucan Dendritic Cell Ligand. Molecules, 23(8), 1961. https://doi.org/10.3390/molecules23081961