Soybean-Hop Alleviates Estrogen Deficiency-Related Bone Loss and Metabolic Dysfunction in Ovariectomized Rats Fed a High-Fat Diet


Abstract
1. Introduction
2. Results
2.1. Effects of Soy-Hop on Food Intake, Body Weight, and Metabolic Parameters
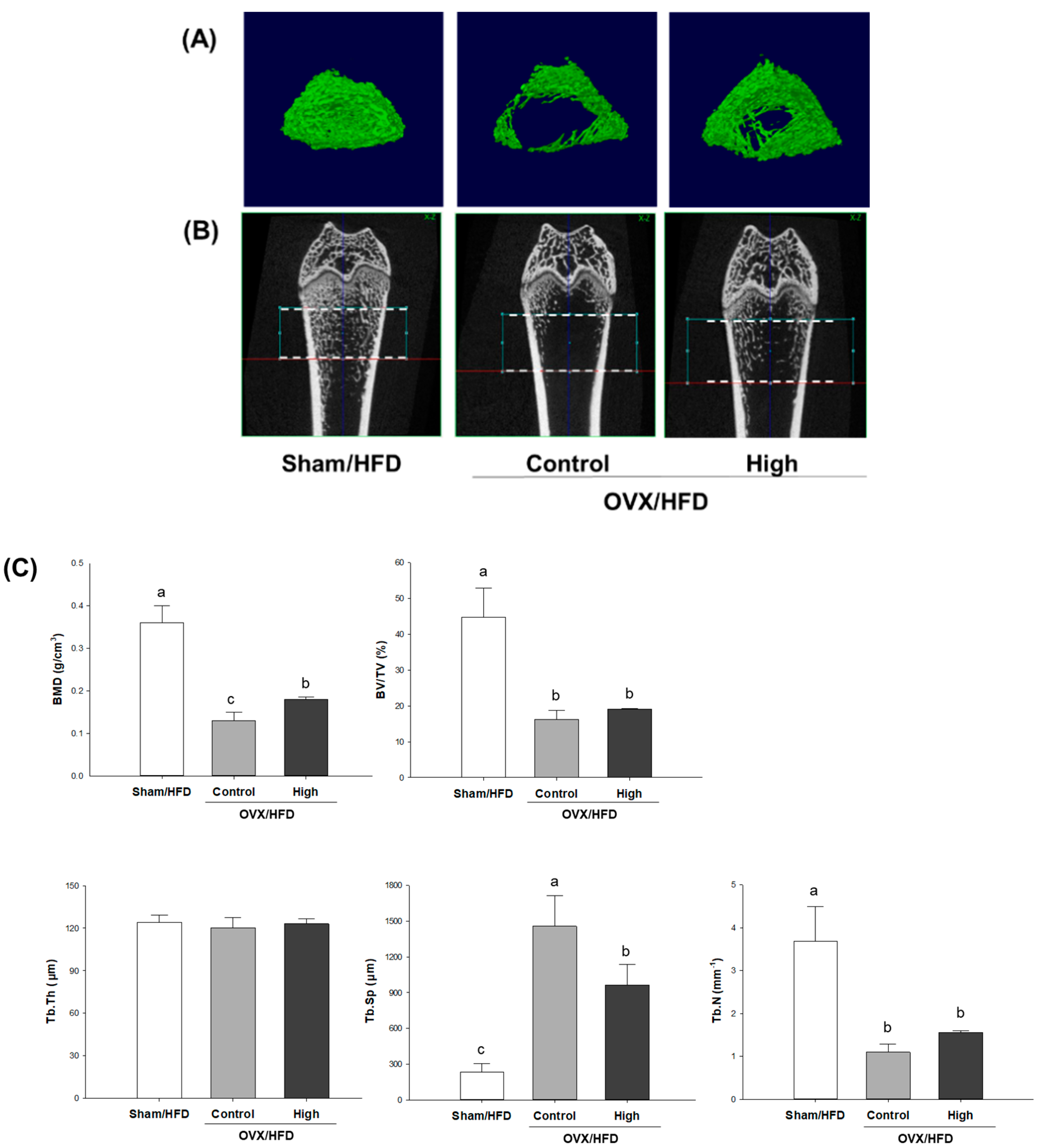
2.2. Effects of Soy-Hop on Dynamics of Bone Formation and Bone Resorption
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Treatments
4.3. Biochemical Assays
4.4. Quantitative Real-Time PCR Analysis
4.5. Immunohistochemical Staining
4.6. Microcomputed Tomography (Micro-CT) Scanning
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yonezawa, R.; Wada, T.; Matsumoto, N.; Morita, M.; Sawakawa, K.; Ishii, Y.; Sasahara, M.; Tsuneki, H.; Saito, S.; Sasaoka, T. Central versus peripheral impact of estradiol on the impaired glucose metabolism in ovariectomized mice on a high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E445–E456. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M.; Gent, L.; Davis, K.; Clegg, D.J. Metabolic impact of sex hormones on obesity. Brain Res. 2010, 1350, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Mirhashemi, F.; Liehl, B.; Taugner, F.; Kluth, O.; Kluge, R.; Joost, H.G.; Schurmann, A. Estrogen deficiency aggravates insulin resistance and induces β-cell loss and diabetes in female New Zealand obese mice. Horm Metab. Res. 2013, 45, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Clegg, D.J.; Brown, L.M.; Woods, S.C.; Benoit, S.C. Gonadal hormones determine sensitivity to central leptin and insulin. Diabetes 2006, 55, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Beral, V. Breast cancer and hormone-replacement therapy in the Million Women Study. Lancet 2003, 362, 419–427. [Google Scholar] [CrossRef]
- James, V.; Pamela, J.; Jay, H. Menopausal Hormone Replacement Therapy and Risk of Ovarian Cancer. JAMA 2002, 288, 334–341. [Google Scholar]
- Jungbauer, A.; Medjakovic, S. Phytoestrogens and the metabolic syndrome. J. Steroid Biochem. Mol. Biol. 2014, 139, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Lethaby, A.; Marjoribanks, J.; Kronenberg, F.; Roberts, H.; Eden, J.; Brown, J. Phytoestrogens for menopausal vasomotor symptoms. Cochrane Database Syst. Rev. 2013, 13, 1–98. [Google Scholar] [CrossRef] [PubMed]
- Overk, C.R.; Guo, J.; Chadwick, L.R.; Lantvit, D.D.; Minassi, A.; Appendino, G.; Chen, S.-N.; Lankin, D.C.; Farnsworth, N.R.; Pauli, G.F.; et al. In Vivo estrogenic comparisons of Trifolium pratense (red clover) Humulus lupulus (hops), and the pure compounds isoxanthohumol and 8-prenylnaringenin. Chem.-Biol. Interact. 2008, 176, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Overk, C.R.; Yao, P.; Chadwick, L.R.; Nikolic, D.; Sun, Y.; Cuendet, M.A.; Deng, Y.; Hedayat, A.S.; Pauli, G.F.; Farnsworth, N.R.; et al. Comparison of the in vitro estrogenic activities of compounds from hops (Humulus lupulus) and red clover (Trifolium pratense). J. Agric. Food Chem. 2005, 53, 6246–6253. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Keiler, A.M.; Zierau, O.; Kretzschmar, G. Hop extracts and hop substances in treatment of menopausal complaints. Planta Med. 2013, 79, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Alekel, D.L.; Germain, A.S.; Peterson, C.T.; Hanson, K.B.; Stewart, J.W.; Toda, T. Isoflavone-rich soy protein isolate attenuates bone loss in the lumbar spine of perimenopausal women. Am. J. Clin. Nutr. 2000, 72, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Choi, Y.H.; Kwon, H.; Lee, S.B.; Kim, D.H.; Sung, C.K.; Park, Y.I.; Dong, M.S. Estrogenic/antiestrogenic activities of a Epimedium koreanum extract and its major components: In Vitro and In Vivo studies. Food Chem. Toxicol. 2012, 50, 2751–2759. [Google Scholar] [CrossRef] [PubMed]
- Li, C.M.; Dong, X.L.; Fan, X.D.; Wu, J.H.; Wang, Q.H.; Tian, X.L.; Guo, D.J.; Wong, M.S.; Qiu, T.Q.; Chan, S.W. Aqueous extract of danshen (Salvia miltiorrhiza Bunge) protects ovariectomized rats fed with high-fat diet from endothelial dysfunction. Menopause 2013, 20, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Mittal, G.; Chandraiah, G.; Ramarao, P.; Ravi Kumar, M.N. Pharmacodynamic evaluation of oral estradiol nanoparticles in estrogen deficient (ovariectomized) high-fat diet induced hyperlipidemic rat model. Pharm. Res. 2009, 26, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Ludgero-Correia, A., Jr.; Aguila, M.B.; Mandarim-de-Lacerda, C.A.; Faria, T.S. Effects of high-fat diet on plasma lipids, adiposity, and inflammatory markers in ovariectomized C57BL/6 mice. Nutrition 2012, 28, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Ok, H.M.; Gebreamanuel, M.R.; Oh, S.A.; Jeon, H.; Lee, W.J.; Kwon, O. A Root-Based Combination Supplement Containing Pueraria lobata and Rehmannia glutinosa and Exercise Preserve Bone Mass in Ovariectomized Rats Fed a High-Fat Diet. Calcif. Tissue Int. 2015, 97, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.C.; Yoo, S.W.; Cho, G.J.; Kim, T.; Hur, J.Y.; Park, Y.K.; Lee, K.W.; Kim, S.H. Correlation between estrogens and serum adipocytokines in premenopausal and postmenopausal women. Menopause 2007, 14, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Jurimae, J.; Jurimae, T. Plasma adiponectin concentration in healthy pre- and postmenopausal women: Relationship with body composition, bone mineral, and metabolic variables. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E42–E47. [Google Scholar] [CrossRef] [PubMed]
- Kanaley, J.A.; Sames, C.; Swisher, L.; Swick, A.G.; Ploutz-Snyder, L.L.; Steppan, C.M.; Sagendorf, K.S.; Feiglin, D.; Jaynes, E.B.; Meyer, R.A.; et al. Abdominal fat distribution in pre- and postmenopausal women: The impact of physical activity, age, and menopausal status. Metabolism 2001, 50, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Isobe, T.; Saitoh, S.; Takagi, S.; Takeuchi, H.; Chiba, Y.; Katoh, N.; Shimamoto, K. Influence of gender, age and renal function on plasma adiponectin level: The Tanno and Sobetsu study. Eur. J. Endocrinol. 2005, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Ryder, J.W.; Chibalin, A.V.; Zierath, J.R. Intracellular mechanisms underlying increases in glucose uptake in response to insulin or exercise in skeletal muscle. Acta Physiol. Scand. 2001, 171, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.D.; Singrakhia, M.D. Painful osteoporotic vertebral fracture. Pathogenesis, evaluation, and roles of vertebroplasty and kyphoplasty in its management. J. Bone Jt. Surg. Am. 2003, 85, 2010–2022. [Google Scholar] [CrossRef]
- Seibel, M.J. Biochemical markers of bone turnover: Part I: Biochemistry and variability. Clin. Biochem. Rev. 2005, 26, 97–122. [Google Scholar] [PubMed]
- Vasikaran, S.D. Utility of biochemical markers of bone turnover and bone mineral density in management of osteoporosis. Crit. Rev. Clin. Lab. Sci. 2008, 45, 221–258. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, Y.; Nakamura, T.; Ohta, H.; Kushida, K.; Gorai, I.; Shiraki, M.; Fukunaga, M.; Hosoi, T.; Miki, T.; Chaki, O.; et al. Guidelines for the use of biochemical markers of bone turnover in osteoporosis (2004). J. Bone Miner. Metab. 2005, 23, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, M.C.; Carvalho, J.F.; Prokopowitsch, A.S.; Pereira, R.M. RANK, RANKL and osteoprotegerin in arthritic bone loss. Braz. J. Med. Biol. Res. 2005, 38, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, S.I. Bone metabolism and the cardiometabolic syndrome: Pathophysiologic insights. J. Cardiometab. Syndr. 2006, 1, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Haam, J.H.; Kim, Y.S.; Kim, M.J.; Koo, H.S.; Kim, H.Y.; Kim, H.J.; Park, K.H.; Joo, N.S.; Park, K.C. A cross-sectional study of the association between adipokine levels and bone mineral density according to obesity and menopausal status in Korean women. J. Bone Miner. Metab. 2017, 35, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Ferron, M. The contribution of bone to whole-organism physiology. Nature 2012, 481, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Chedraui, P.; Perez-Lopez, F.R.; Escobar, G.S.; Palla, G.; Montt-Guevara, M.; Cecchi, E.; Genazzani, A.R.; Simoncini, T.; Research Group for the Omega Women’s Health Project. Circulating leptin, resistin, adiponectin, visfatin, adipsin and ghrelin levels and insulin resistance in postmenopausal women with and without the metabolic syndrome. Maturitas 2014, 79, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |




{kind=link}
{kind=link}
{kind=link}
| Variables | Sham/HFD | OVX/HFD | p-Values | |||
|---|---|---|---|---|---|---|
| Control | Low | Medium | High | |||
| Total food intake (g) | 520.2 ± 15.9 bc | 587.3 ± 18.0 a | 538.5 ± 12.7 bc | 554.9 ± 16.3 ab | 502.0 ± 14.9 c | 0.0045 |
| Total body weight gain (g) | 118.1 ± 11.1 cd | 181.0 ± 12.5 a | 146.6 ± 7.5 bc | 151.2 ± 8.6 b | 110.0 ± 9.7 d | <0.0001 |
| Total fat pad (g) | 30.4 ± 3.3 b | 41.3 ± 3.2 a | 34.6 ± 2.1 ab | 31.7 ± 2.3 b | 27.6 ± 2.1 b | 0.0081 |
| Soleus muscle weight (mg) | 236.0 ± 12.5 | 266.0 ± 12.7 | 260.0 ± 8.4 | 262.0 ± 7.9 | 238.0 ± 8.8 | 0.1226 |
| Femur weight (mg) | 905.5 ± 16.3 | 882.0 ± 22.7 | 878.5 ± 14.9 | 865.0 ± 19.7 | 880.0 ± 24.6 | 0.4770 |
| Uterus weight (mg) | 514.0 ± 35.2 a | 122.0 ± 8.4 b | 138.0 ± 24.4 b | 101.0 ± 9.8 b | 151.0 ± 21.2 b | <0.0001 |
| Leptin (ng/mL) | 9.3 ± 1.7 b | 17.9 ± 2.2 a | 14.2 ± 2.5 ab | 10.9 ± 1.5 b | 9.3 ± 1.5 b | 0.0106 |
| Adiponectin (μg/mL) | 7.3 ± 0.3 c | 11.7 ± 0.8 a | 10.7 ± 1.3 ab | 12.3 ± 0.8 a | 8.8 ± 0.5 bc | 0.0001 |
| LDL-cholesterol (mg/dL) | 29.3 ± 3.9 c | 62.4 ± 9.3 a | 47.9 ± 6.9 abc | 50.9 ± 5.3 ab | 31.4 ± 6.1 bc | 0.0034 |
| HDL-cholesterol (mg/dL) | 61.5 ± 6.2 | 58.4 ± 6.4 | 62.0 ± 8.2 | 47.3 ± 2.1 | 44.8 ± 5.0 | 0.1178 |
| TC (mg/dL) | 113.0 ± 6.3 ab | 139.5 ± 13.0 a | 129.6 ± 9.0 a | 113.2 ± 6.7 ab | 88.9 ± 8.5 b | 0.0035 |
| TG (mg/dL) | 96.6 ± 9.8 a | 93.7 ± 13.0 a | 86.3 ± 7.6 a | 74.9 ± 6.0 ab | 59.1 ± 3.0 b | 0.0283 |
| FBG (mmol/L) | 15.8 ± 0.9 c | 22.2 ± 1.4 a | 19.6 ± 1.5 ab | 19.2 ± 1.1 abc | 17.2 ± 1.0 bc | 0.0050 |
| Insulin (μg/L) | 34.4 ± 4.7 ab | 34.9 ± 3.1 ab | 46.7 ± 6.8 a | 31.3 ± 4.6 b | 25.7 ± 3.7 b | 0.0486 |
| C-peptide (nmol/L) | 1.5 ± 0.2 b | 1.5 ± 0.1 b | 2.0 ± 0.3 a | 1.3 ± 0.1 b | 1.1 ± 0.1 b | 0.0126 |
| HOMA-IR | 24.5 ± 3.9 b | 34.8 ± 5.0 ab | 42.8 ± 7.0 a | 26.3 ± 5.0 b | 20.3 ± 3.6 b | 0.0179 |
| Estradiol (ng/L) | 26.4 ± 4.2 | 23.7 ± 3.9 | 23.3 ± 3.3 | 24.0 ± 3.4 | 20.9 ± 3.3 | 0.8780 |
| Variables | Sham/HFD | OVX/HFD | p-Values | |||
|---|---|---|---|---|---|---|
| Control | Low | Medium | High | |||
| Osteocalcin (pg/mL) | 2824.8 ± 114.1 b | 3749.0 ± 372.8 a | 2388.4 ± 124.3 bc | 2938.7 ± 266.3 b | 1919.9 ± 87.1 c | <0.0001 |
| ALP (ng/mL) | 446.2 ± 87.8 ab | 717.2 ± 208.2 a | 377.9 ± 103.1 ab | 266.6 ± 64.2 b | 198.8 ± 107.9 b | 0.0474 |
| CTX (ng/mL) | 11.4 ± 0.4 c | 15.9 ± 0.8 a | 14.7 ± 0.6 ab | 15.4 ± 1.1 a | 12.3 ± 1.6 bc | 0.0122 |
| NTX (nM BCE) | 59.8 ± 3.2 | 73.2 ± 4.3 | 71.7 ± 6.1 | 62.6 ± 3.7 | 68.1 ± 5.9 | 0.2493 |
| RANKL | 1.0 ± 0.2 b | 1.5 ± 0.2 a | 1.3 ± 0.1 ab | 1.2 ± 0.1 ab | 1.2 ± 0.1 ab | 0.0671 |
| OPG | 1.0 ± 0.5 | 0.9 ± 0.3 | 1.0 ± 0.4 | 1.4 ± 0.4 | 1.6 ± 0.5 | 0.6675 |
| RANKL/OPG | 1.0 ± 0.2 ab | 1.4 ± 0.4 a | 0.6 ± 0.3 b | 0.5 ± 0.2 b | 0.4 ± 0.2 b | 0.0610 |
| Formulation | g % | kcal % |
| Protein | 24 | 20 |
| Carbohydrate | 41 | 35 |
| Fat | 24 | 45 |
| kcal/kg | 4776 | |
| Ingredients | g | kcal |
| Casein (from milk) | 238.8 | 800 |
| Corn starch | 185.1 | 620 |
| Sucrose | 59.7 | 200 |
| Dextrose | 157.6 | 528 |
| Cellulose | 59.7 | 0 |
| Soybean oil | 29.9 | 225 |
| Lard | 208.9 | 1575 |
| Mineral mixture | 41.8 | 0 |
| Vitamin mixture | 11.9 | 40 |
| tert-Butylhydroquinone | 0.02 | 0 |
| l-Cysteine | 3.6 | 12 |
| Choline bitartrate | 3.0 | 0 |
| Total | 1000.02 | 4000 |
| Name | GenBank No. | Sequence (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|
| RANKL | NM057149 | F: 5′-GCA GCA TCG CTC TGT TCC TGT A-3′ | 164 |
| R: 5′-GCA TGA GRC AGG TAG TGC TTC TGT G-3′ | |||
| OPG | NM012870 | F: 5′-GGC AGG GCA TAC TTC CTG TT-3′ | 109 |
| R: 5′-GCC ACT TGT TCA TTG TGG TCC-3′ | |||
| β-actin | NM031144 | F: 5′-CTC TGT GTG GAT TGG TGG CT-3′ | 150 |
| R: 5′-GGG TGT AAA ACG CAG CTC AG-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noh, D.; Lim, Y.; Lee, H.; Kim, H.; Kwon, O. Soybean-Hop Alleviates Estrogen Deficiency-Related Bone Loss and Metabolic Dysfunction in Ovariectomized Rats Fed a High-Fat Diet. Molecules 2018, 23, 1205. https://doi.org/10.3390/molecules23051205
Noh D, Lim Y, Lee H, Kim H, Kwon O. Soybean-Hop Alleviates Estrogen Deficiency-Related Bone Loss and Metabolic Dysfunction in Ovariectomized Rats Fed a High-Fat Diet. Molecules. 2018; 23(5):1205. https://doi.org/10.3390/molecules23051205
Chicago/Turabian StyleNoh, Dasom, Yeni Lim, Hansol Lee, Hyejin Kim, and Oran Kwon. 2018. "Soybean-Hop Alleviates Estrogen Deficiency-Related Bone Loss and Metabolic Dysfunction in Ovariectomized Rats Fed a High-Fat Diet" Molecules 23, no. 5: 1205. https://doi.org/10.3390/molecules23051205
APA StyleNoh, D., Lim, Y., Lee, H., Kim, H., & Kwon, O. (2018). Soybean-Hop Alleviates Estrogen Deficiency-Related Bone Loss and Metabolic Dysfunction in Ovariectomized Rats Fed a High-Fat Diet. Molecules, 23(5), 1205. https://doi.org/10.3390/molecules23051205