Xylosylated Detoxification of the Rice Flavonoid Phytoalexin Sakuranetin by the Rice Sheath Blight Fungus Rhizoctonia solani


Abstract
:
1. Introduction
2. Results and Discussion
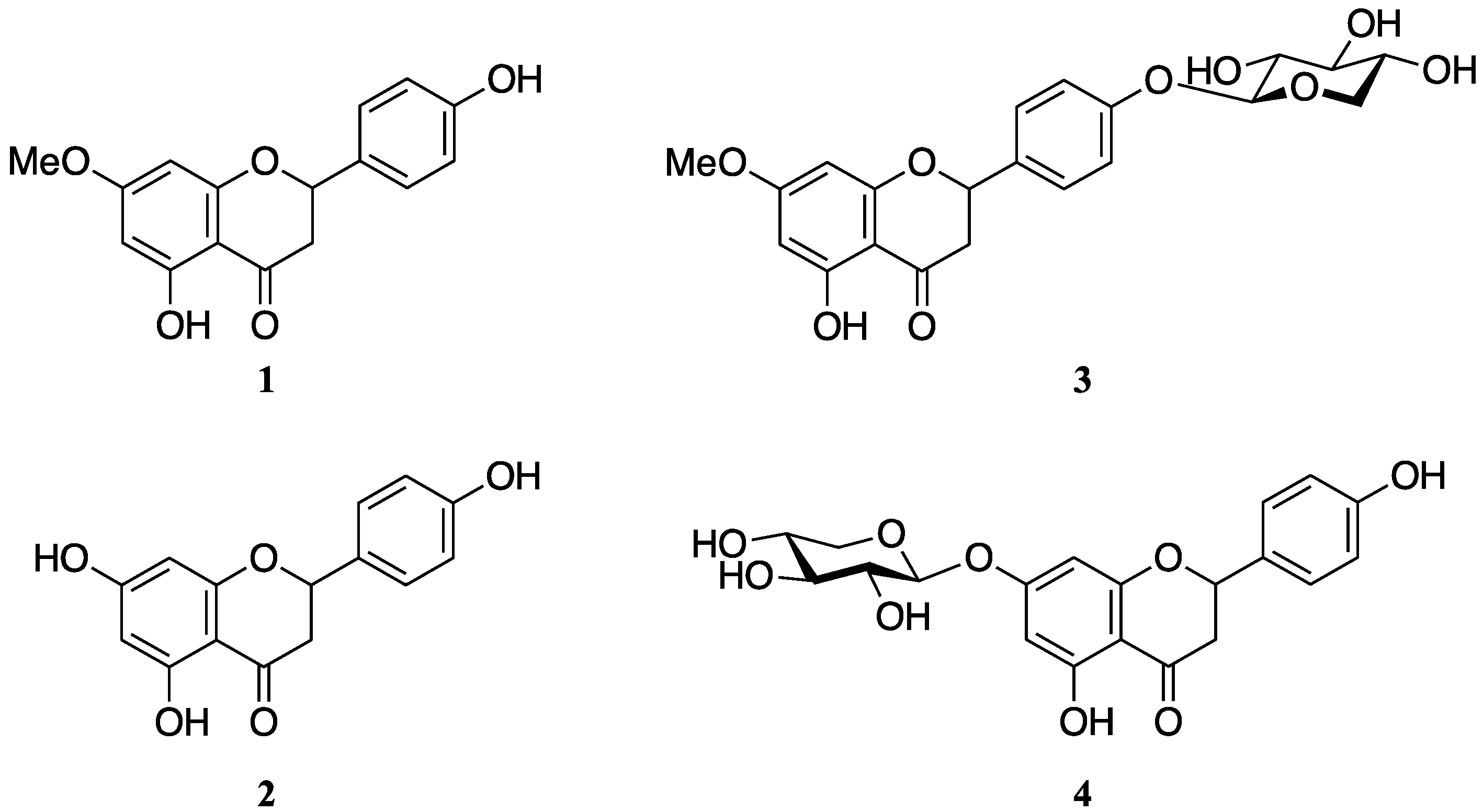
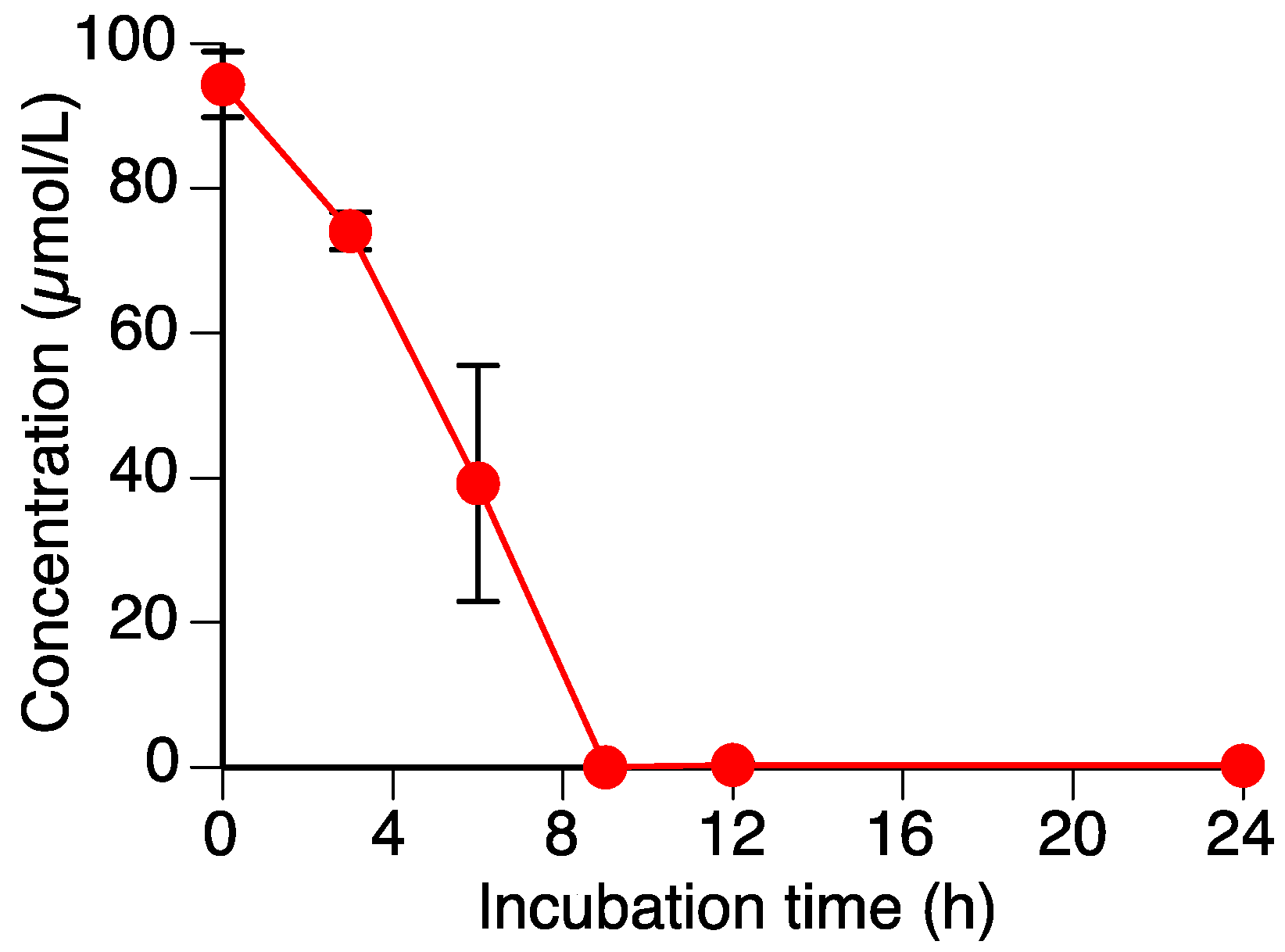
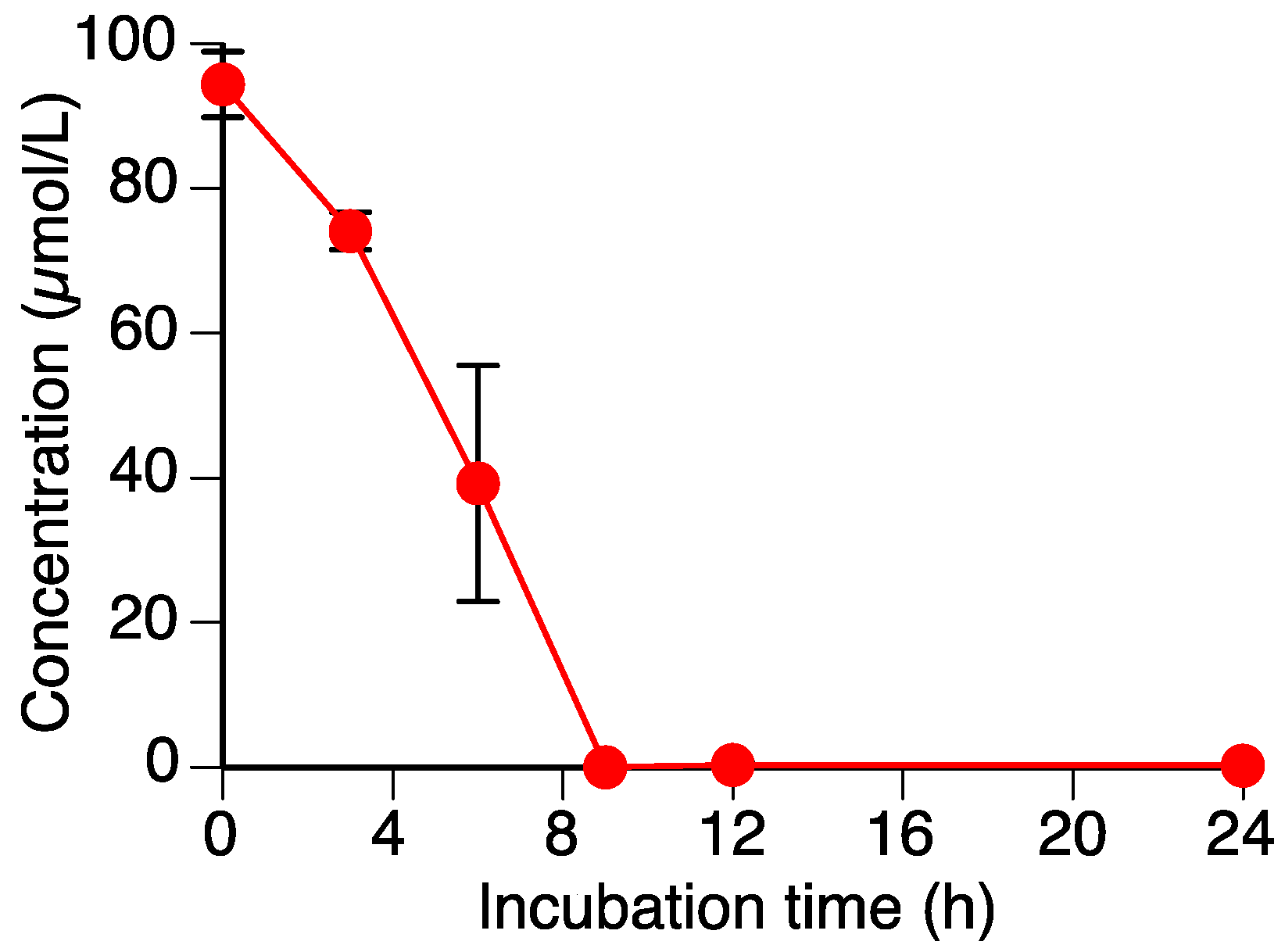
2.1. Sakuranetin (1) is Metabolized by Rhizoctonia solani
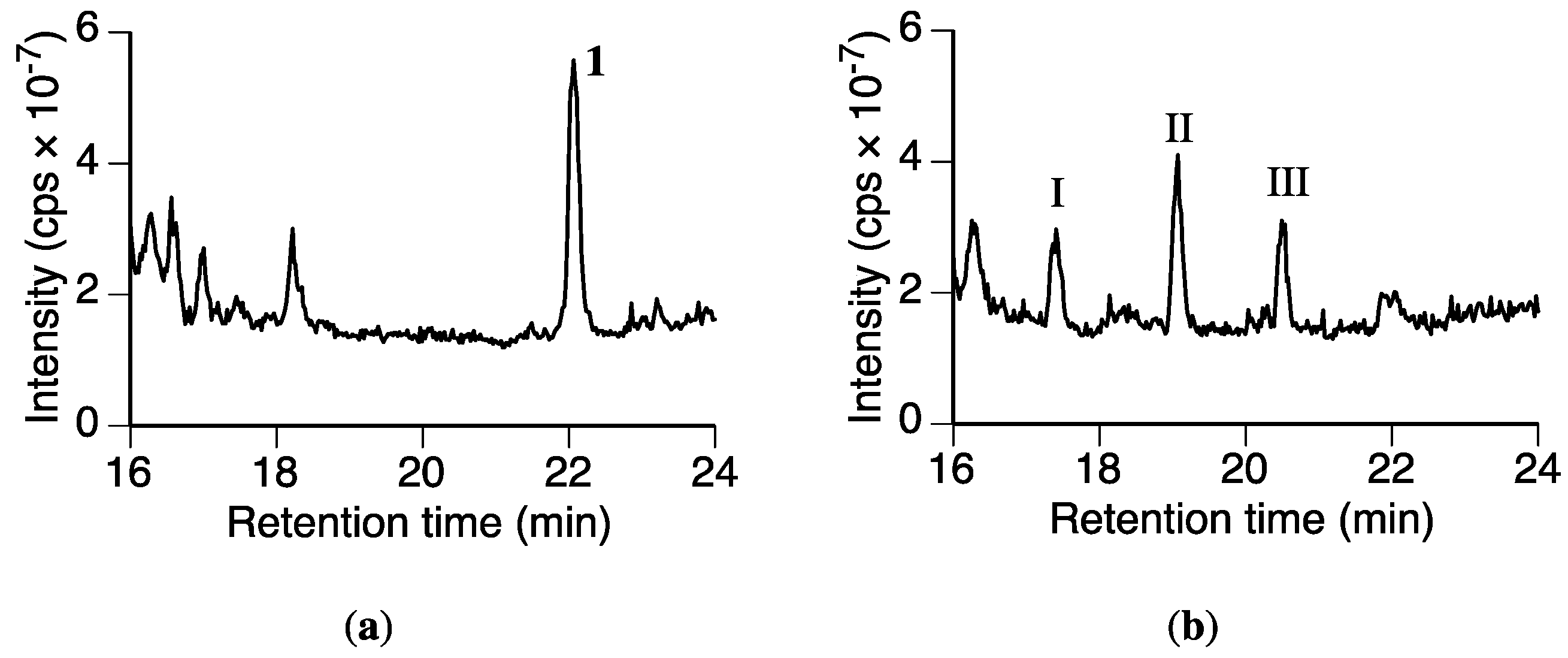
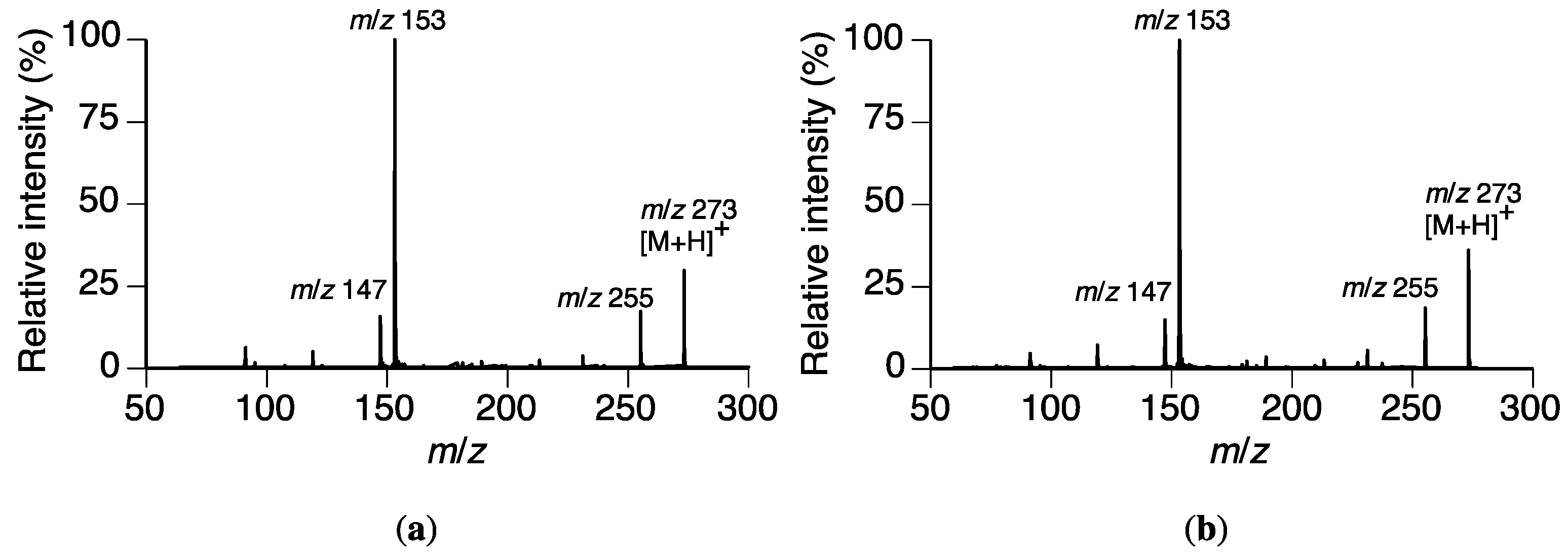
2.2. Identification of Naringenin (2) in the Rhizoctonia solani Suspension Culture Containing Sakuranetin (1)
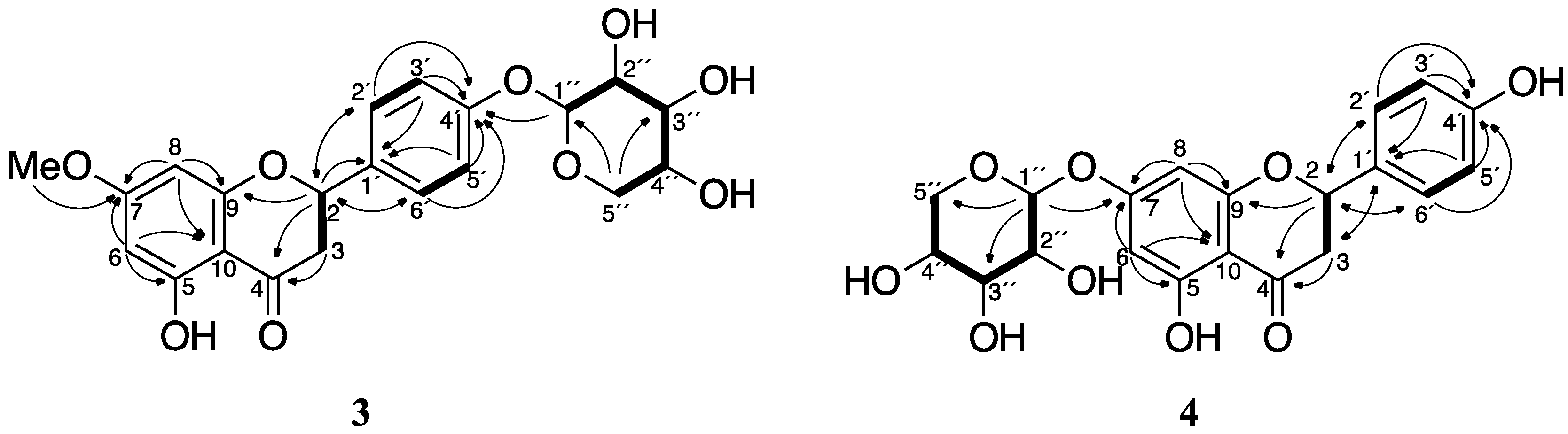
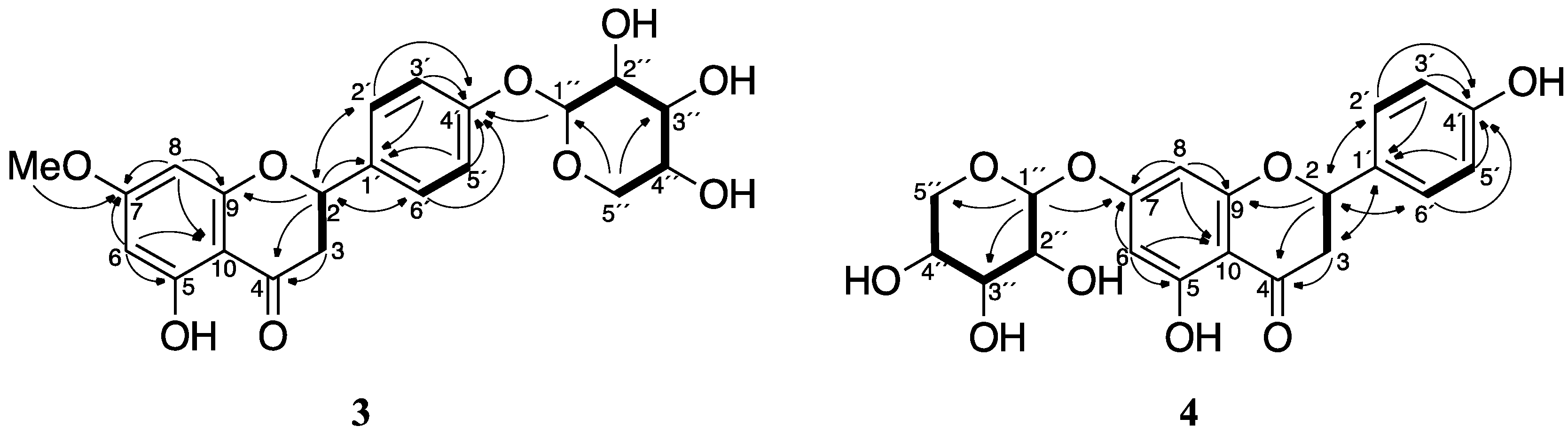
2.3. Purification and Identification of Sakuranetin-4′-O-β-d-xylopyranoside (3) and Naringenin-7-O-β-d-xylopyranoside (4) in Rhizoctonia solani Suspension Culture Containing Sakuranetin (1)
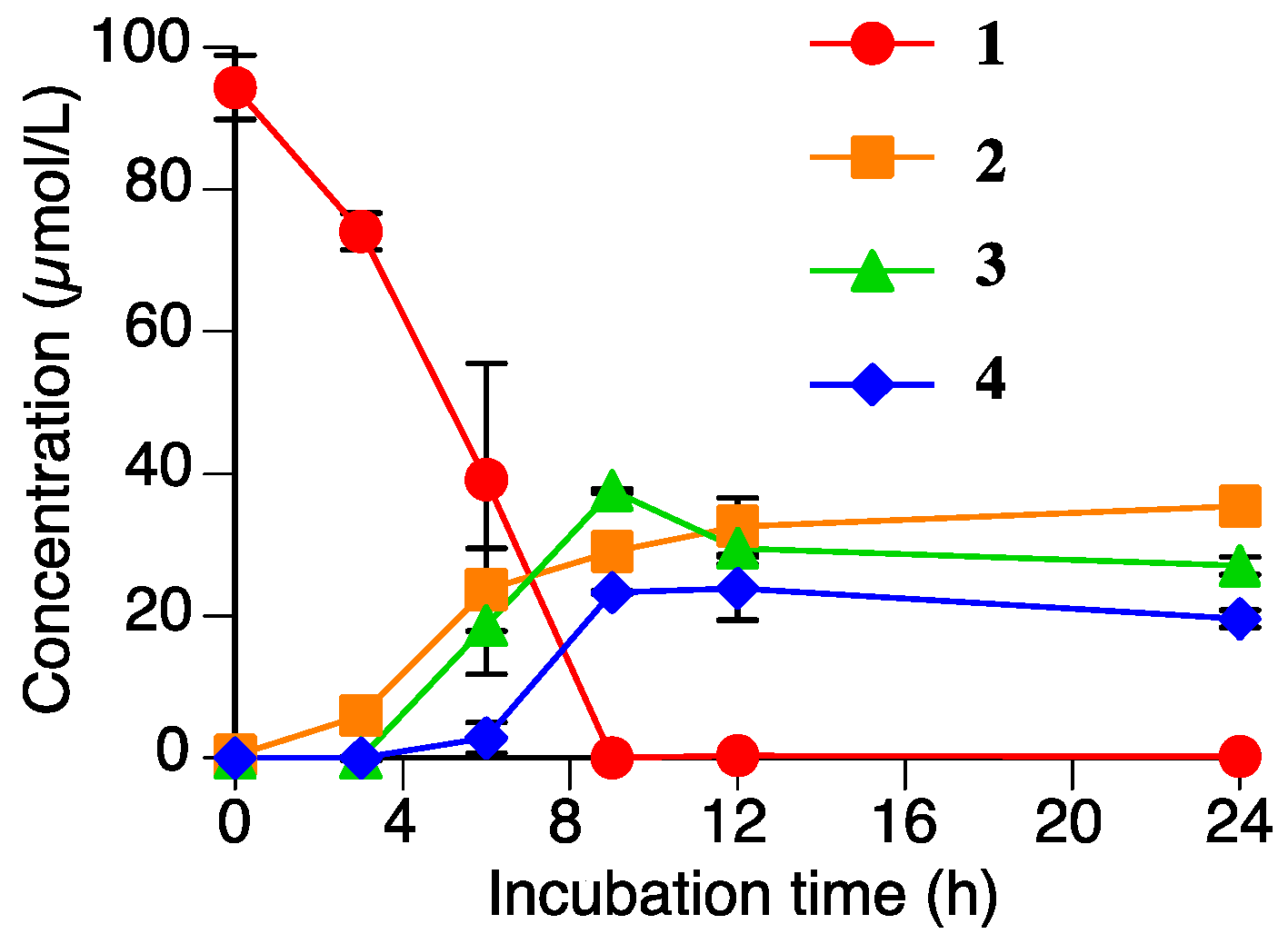
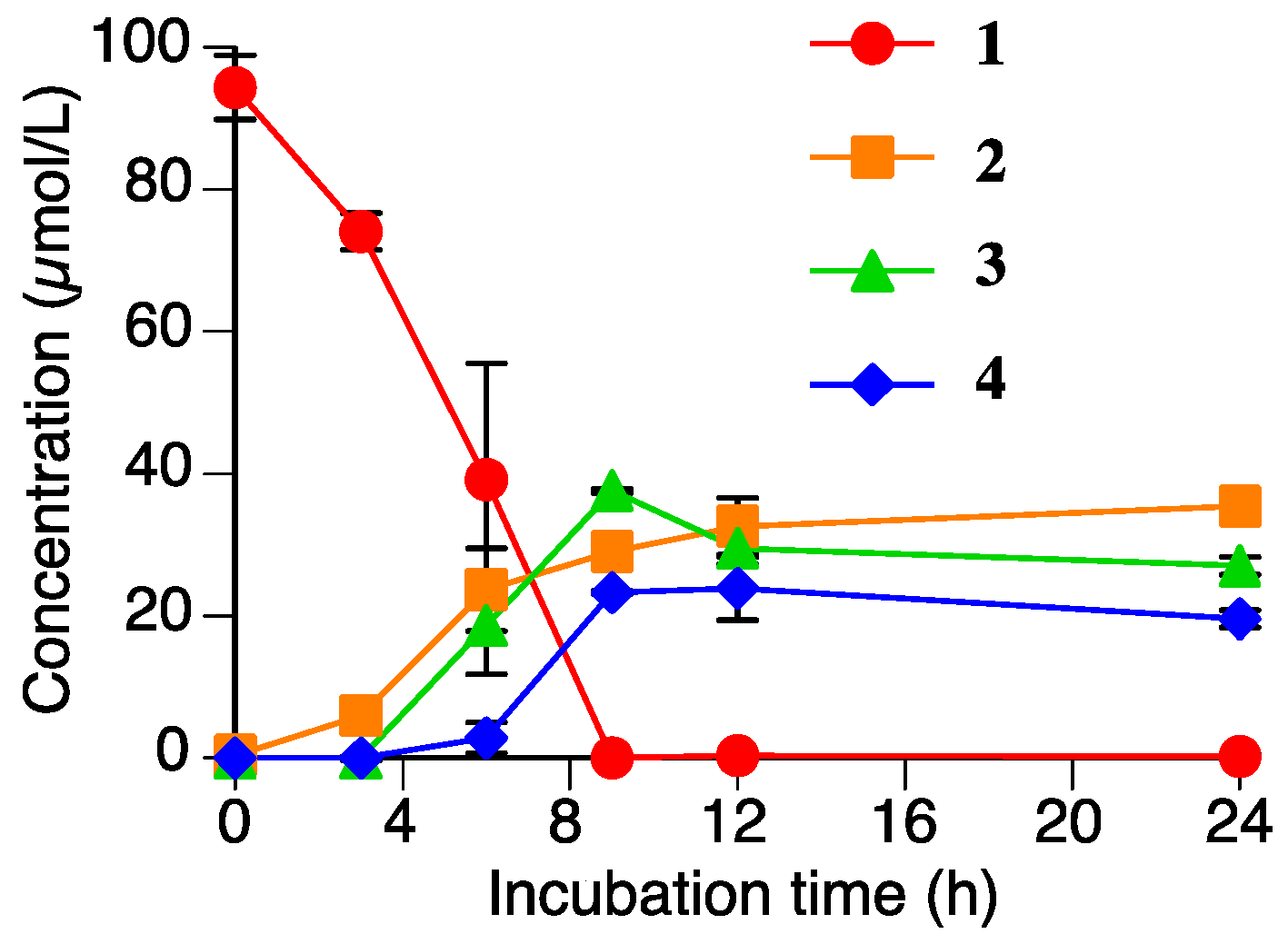
2.4. Accumulation of Naringenin (2), Sakuranetin-4′-O-β-d-xylopyranoside (3), and Naringenin-7-O-β-d-xylopyranoside (4) in the Rhizoctonia solani Suspension Culture Containing Sakuranetin (1)
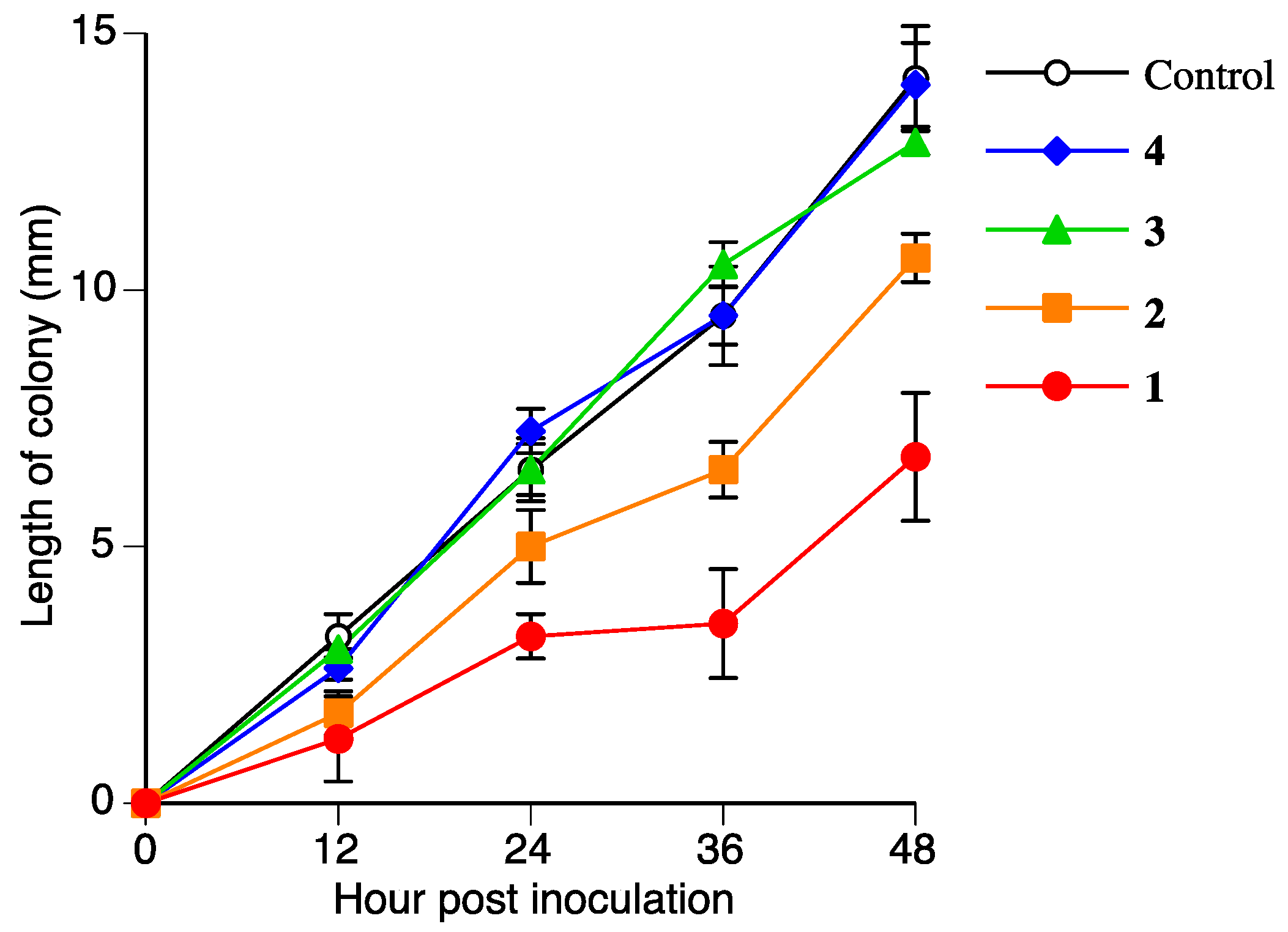
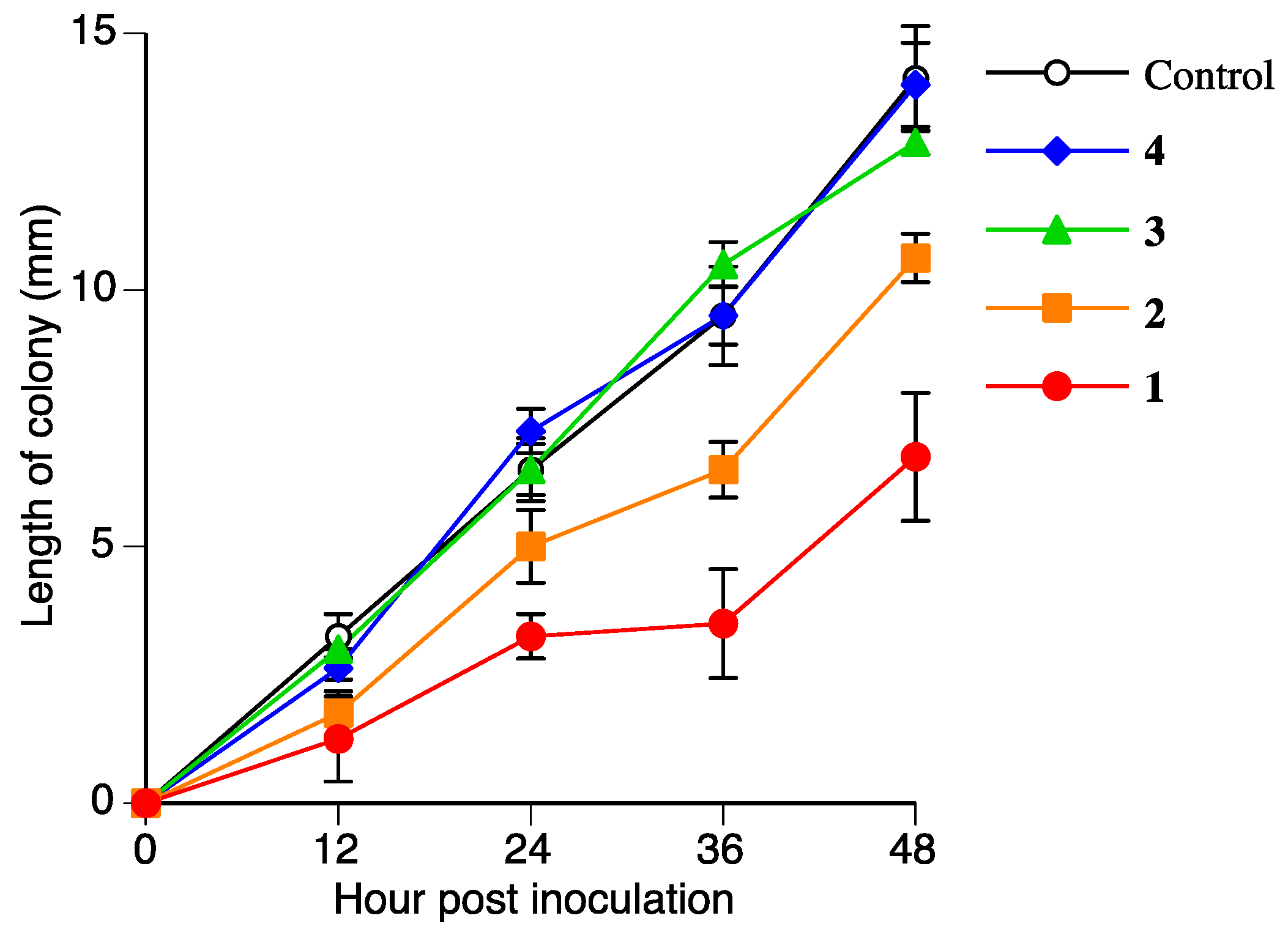
2.5. Antifungal Activities of Naringenin (2), Sakuranetin-4′-O-β-d-xylopyranoside (3), and Naringenin-7-O-β-d-xylopyranoside (4)
3. Materials and Methods
3.1. General Analytical Methods
3.2. Chemicals
3.3. Fungal Material
3.4. Incubation of the Rhizoctonia solani Suspension Cultures with Sakuranetin (1)
3.5. Screening of the Sakuranetin (1) Metabolites from the Rhizoctonia solani Suspension Culture
3.6. LC-MS/MS Analysis to Identify Naringenin (2) as a Metabolite
3.7. Purification of Sakuranetin-4′-O-β-d-xylopyranoside (3) and Naringenin-7-O-β-d-xylopyranoside (4) from the Rhizoctonia solani Suspension Culture
3.8. GC-MS Analysis of the Hydrolysates of Sakuranetin-4′-O-β-d-xylopyranoside (3) and Naringenin-7-O-β-d-xylopyranoside (4)
3.9. Quantitation of Sakuranetin (1) and Its Metabolites in the Rhizoctonia solani Suspension Culture
3.10. Assay of the Antifungal Activity Against Rhizoctonia solani
4. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Horie, K.; Sakai, K.; Okugi, M.; Toshima, H.; Hasegawa, M. Ultraviolet-induced amides and casbene diterpenoids from rice leaves. Phytochem. Lett. 2016, 15, 57–62. [Google Scholar] [CrossRef]
- Hasegawa, M.; Mitsuhara, I.; Seo, S.; Okada, K.; Yamane, H.; Iwai, T.; Ohashi, Y. Analysis on blast fungus-responsive characters of a flavonoid phytoalexin sakuranetin; accumulation in infected rice leaves, antifungal activity and detoxification by fungus. Molecules 2014, 19, 11404–11418. [Google Scholar] [CrossRef] [PubMed]
- Toyomasu, T.; Usui, M.; Sugawara, C.; Otomo, K.; Hirose, Y.; Miyao, A.; Hirochika, H.; Okada, K.; Shimizu, T.; Koga, J.; et al. Reverse genetic approach to verify physiological roles of rice phytoalexins: Characterization of a knockdown mutant of OsCPS4 phytoalexin biosynthetic gene in rice. Physiol. Plant. 2014, 150, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Mitsuhara, I.; Seo, S.; Imai, T.; Koga, J.; Okada, K.; Yamane, H.; Ohashi, Y. Phytoalexin accumulation in the interaction between rice and the blast fungus. Mol. Plant Microbe Interact. 2010, 23, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Abdoli, A. Pathogen inactivation of cruciferous phytoalexins: Detoxification reactions, enzymes and inhibitors. RSC Adv. 2017, 7, 23633–23646. [Google Scholar] [CrossRef]
- VanEtten, H.D.; Matthews, D.E.; Matthews, P.S. Phytoalexin detoxification: Importance for pathogenicity and practical implications. Annu. Rev. Phytopathol. 1989, 27, 143–164. [Google Scholar] [CrossRef] [PubMed]
- VanEtten, H.; Temporini, E.; Wasmann, C. Phytoalexin (and phytoanticipin) tolerance as a virulence trait: Why is it not required by all pathogens? Physiol. Mol. Plant Pathol. 2001, 59, 83–93. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Ahiahonu, P.W.K. Metabolism and detoxification of phytoalexins and analogs by phytopathogenic fungi. Phytochemistry 2005, 66, 391–411. [Google Scholar] [CrossRef] [PubMed]
- Katsumata, S.; Hamana, K.; Horie, K.; Toshima, H.; Hasegawa, M. Identification of sternbin and naringenin as detoxified metabolites from the rice flavanone phytoalexin sakuranetin by Pyricularia oryzae. Chem. Biodiver. 2017, 14, e1600240. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Ohashi, Y.; Mitsuhara, I.; Seo, S.; Toshima, H.; Hasegawa, M. Identification of a degradation intermediate of the momilactone A rice phytoalexin by the rice blast fungus. Biosci. Biotechnol. Biochem. 2012, 76, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Ogoshi, A. Ecology and pathogenicity of anastomosis and intraspecific groups of Rhizoctonia solani Kühn. Annu. Rev. Phytopathol. 1987, 25, 125–143. [Google Scholar] [CrossRef]
- Park, H.L.; Yoo, Y.; Hahn, T.-R.; Bhoo, S.H.; Lee, S.-W.; Cho, M.-H. Antimicrobial activity of UV-induced phenylamides from rice leaves. Molecules 2014, 19, 18139–18151. [Google Scholar] [CrossRef] [PubMed]
- Paez, M.; Martínez-Castro, I.; Sanz, J.; Olano, A.; Garcia-Raso, A.; Saura-Calixto, F. Identification of the components of aldoses in a tautomeric equilibrium mixture as their trimethylsilyl ethers by capillary gas chromatography. Chromatographia 1987, 23, 43–46. [Google Scholar] [CrossRef]
- Simkhada, D.; Kim, E.; Lee, H.C.; Sohng, J.K. Metabolic engineering of Escherichia coli for the biological synthesis of 7-O-xylosyl naringenin. Mol. Cells 2009, 28, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Aida, Y.; Tamogami, S.; Kodama, O.; Tsukiboshi, T. Synthesis of 7-methoxyapigeninidin and its fungicidal activity against Gloeocercospora sorghi. Biosci. Biotechnol. Biochem. 1996, 60, 1495–1496. [Google Scholar] [CrossRef] [PubMed]
- Füzfai, Z.; Molnár-Perl, I. Gas chromatographic–mass spectrometric fragmentation study of flavonoids as their trimethylsilyl derivatives: Analysis of flavonoids, sugars, carboxylic and amino acids in model systems and in citrus fruits. J. Chromatogr. A 2007, 1149, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Horie, K.; Inoue, Y.; Sakai, M.; Yao, Q.; Tanimoto, Y.; Koga, J.; Toshima, H.; Hasegawa, M. Identification of UV-induced diterpenes including a new diterpene phytoalexin, phytocassane F, from rice leaves by complementary GC/MS and LC/MS approaches. J. Agric. Food Chem. 2015, 63, 4050–4059. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds 3 and 4 are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Compound 3 | Compound 4 | ||
|---|---|---|---|---|
| δC | δH (Multiplicity, J in Hz) | δC | δH (Multiplicity, J in Hz) | |
| 2 | 80.2 | 5.45 (dd, 12.7, 3.1) | 80.15/80.17 1 | 5.50 (dd, 13.0, 3.2) |
| 3 | 44.1 | 2.79 (dd, 17.2, 3.1) | 43.58/43.60 1 | 2.78 (m) 2 |
| 3.14 (dd, 17.2, 12.7) | 3.24/3.25 1 (dd, 17.0, 13.0) | |||
| 4 | 197.9 | 197.97/197.99 1 | ||
| 5 | 165.3 | 164.72/164.76 1 | ||
| 6 | 95.8 | 6.05 (d, 2.3) | 97.6 | 6.11/6.12 1 (d, 2.2) |
| 7 | 169.6 | 166.46/166.53 1 | ||
| 8 | 95.0 | 6.08 (d, 2.3) | 96.4 | 6.15 (d, 2.2) |
| 9 | 164.6 | 164.2 | ||
| 10 | 104.1 | 104.5 | ||
| 7-O-Me | 56.3 | 3.81 (s) | – | – |
| 5-OH | – | – | 12.07 | |
| 1′ | 134.2 | 130.63/130.67 1 | ||
| 2′, 6′ | 128.8 | 7.44 (d, 8.7) | 129.10/129.14 1 | 7.41 (d, 8.7) |
| 3′, 5′ | 117.9 | 7.11 (d, 8.7) | 116.3 | 6.91 (d, 8.7) |
| 4′ | 159.2 | 158.84/158.85 1 | ||
| 1″ | 102.8 | 4.90 (d, 7.2) | 101.39/101.46 1 | 5.06/5.07 1 (d, 7.0 or 5.8) 3 |
| 2″ | 74.7 | 3.44 (m)2 | 74.1 | 3.48 (m) 2 |
| 3″ | 77.7 | 3.44 (m)2 | 77.4 | 3.48 (m) 2 |
| 4″ | 71.0 | 3.57 (m) | 70.6 | 3.60 (m) |
| 5″ | 66.9 | 3.37 (dd, 11.4, 10.2) | 66.6 | 3.50 (m) 2 |
| 3.92 (dd, 11.4, 5.3) | 3.91 (dd, 11.2, 4.8) | |||
| Compound | Concentration (µmol/L) | ||
|---|---|---|---|
| 75 | 150 | 300 | |
| Length of Colony [mm] (Inhibition [%]) | |||
| 1 | 9.8 ± 0.6 (31) | 8.1 ± 0.7 (42) | 6.8 ± 1.2 (52) |
| 2 | 14.5 ± 0.6 (−3) | 11.3 ± 0.2 (20) | 10.6 ± 0.5 (25) |
| 3 | 12.8 ± 0.2 (10) | 13.1 ± 0.9 (7) | 12.9 ± 0.2 (9) |
| 4 | 14.4 ± 0.2 (−2) | 14.0 ± 0.2 (1) | 14.0 ± 0.8 (1) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsumata, S.; Toshima, H.; Hasegawa, M. Xylosylated Detoxification of the Rice Flavonoid Phytoalexin Sakuranetin by the Rice Sheath Blight Fungus Rhizoctonia solani. Molecules 2018, 23, 276. https://doi.org/10.3390/molecules23020276
Katsumata S, Toshima H, Hasegawa M. Xylosylated Detoxification of the Rice Flavonoid Phytoalexin Sakuranetin by the Rice Sheath Blight Fungus Rhizoctonia solani. Molecules. 2018; 23(2):276. https://doi.org/10.3390/molecules23020276
Chicago/Turabian StyleKatsumata, Shun, Hiroaki Toshima, and Morifumi Hasegawa. 2018. "Xylosylated Detoxification of the Rice Flavonoid Phytoalexin Sakuranetin by the Rice Sheath Blight Fungus Rhizoctonia solani" Molecules 23, no. 2: 276. https://doi.org/10.3390/molecules23020276
APA StyleKatsumata, S., Toshima, H., & Hasegawa, M. (2018). Xylosylated Detoxification of the Rice Flavonoid Phytoalexin Sakuranetin by the Rice Sheath Blight Fungus Rhizoctonia solani. Molecules, 23(2), 276. https://doi.org/10.3390/molecules23020276