
Fucoidan Exerts Anticancer Effects Against Head and Neck Squamous Cell Carcinoma In Vitro

 , , and
, , and
Abstract
1. Introduction
2. Results
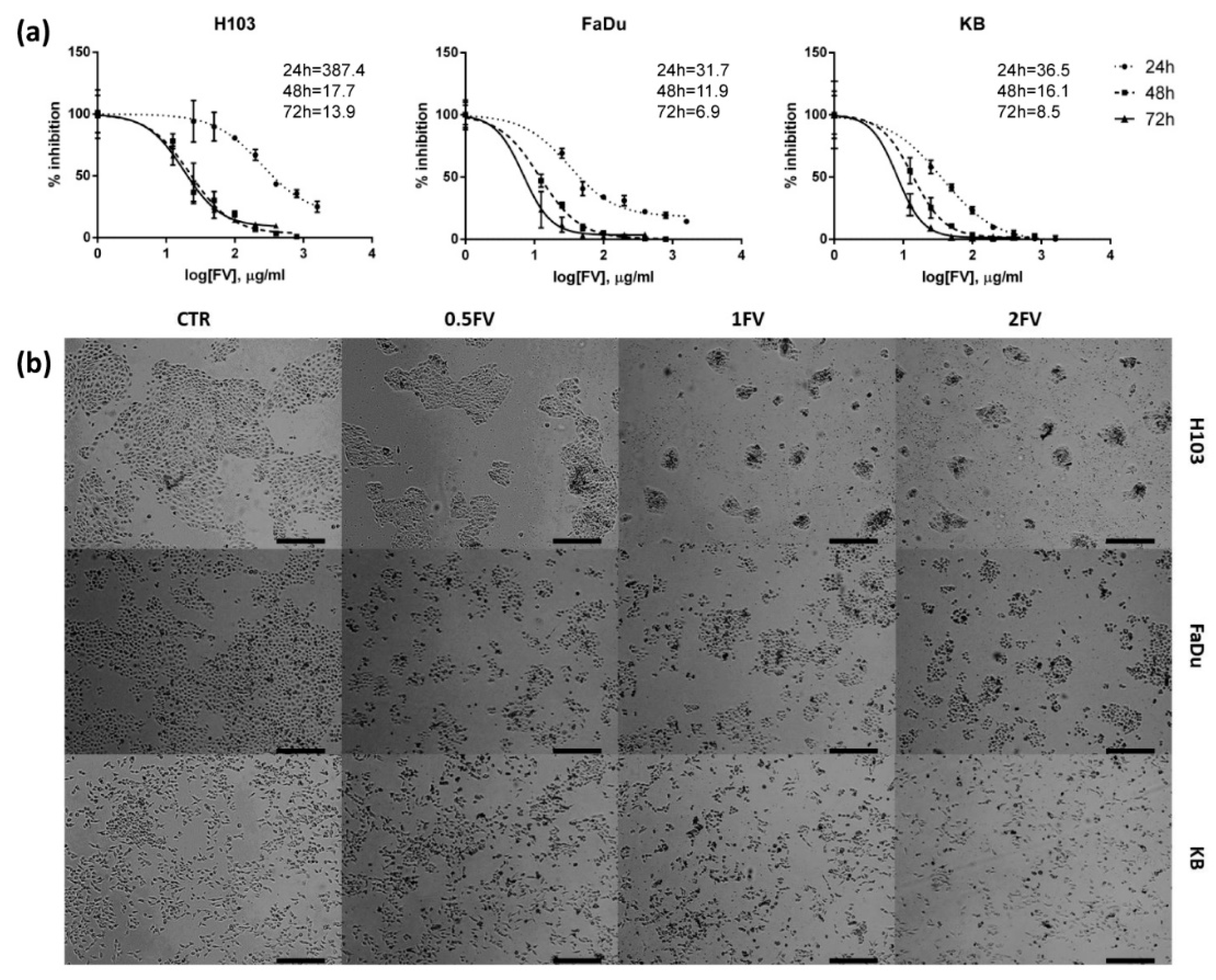
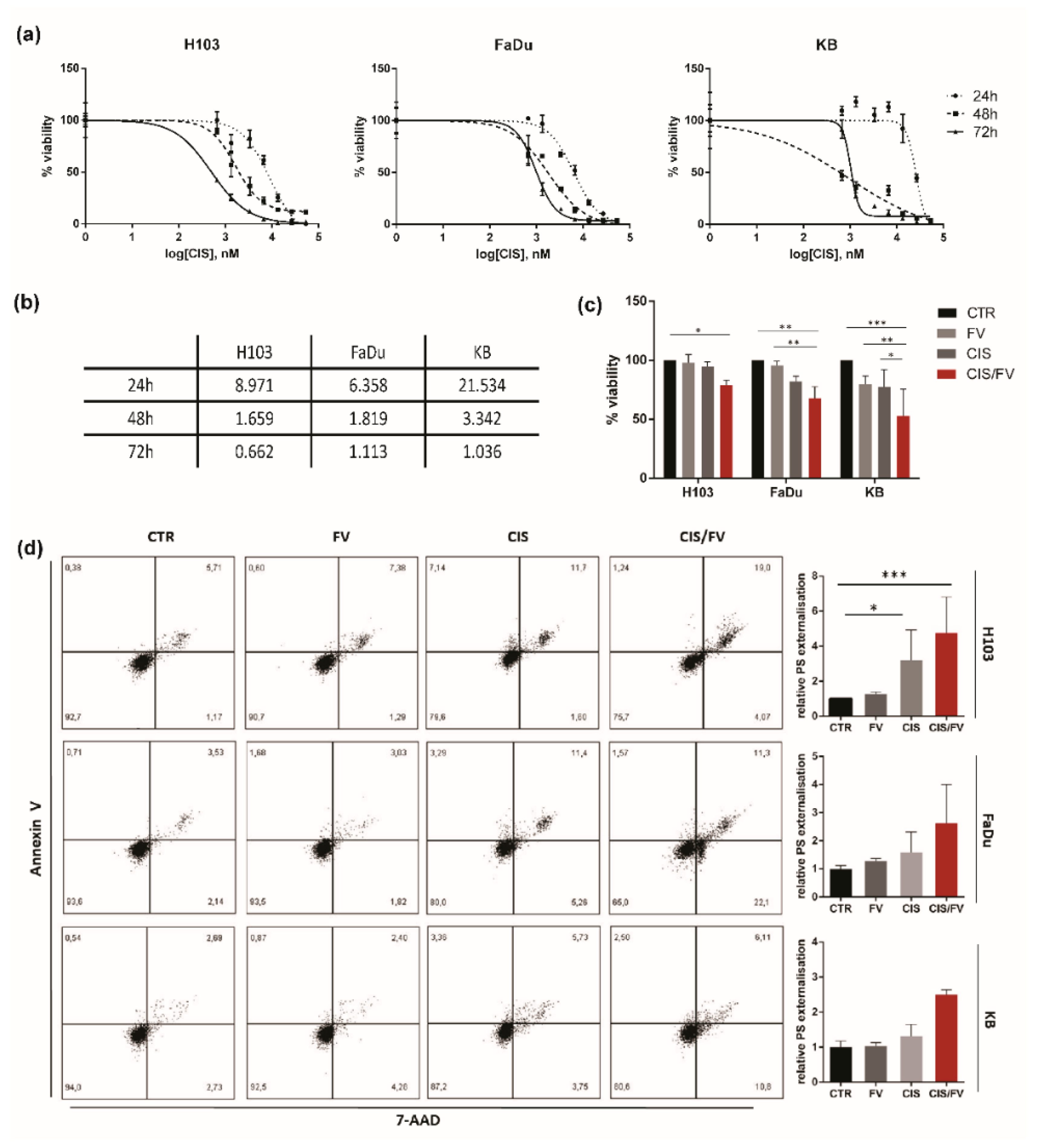
2.1. Fucoidan Inhibits Proliferation of HNSCC In Vitro
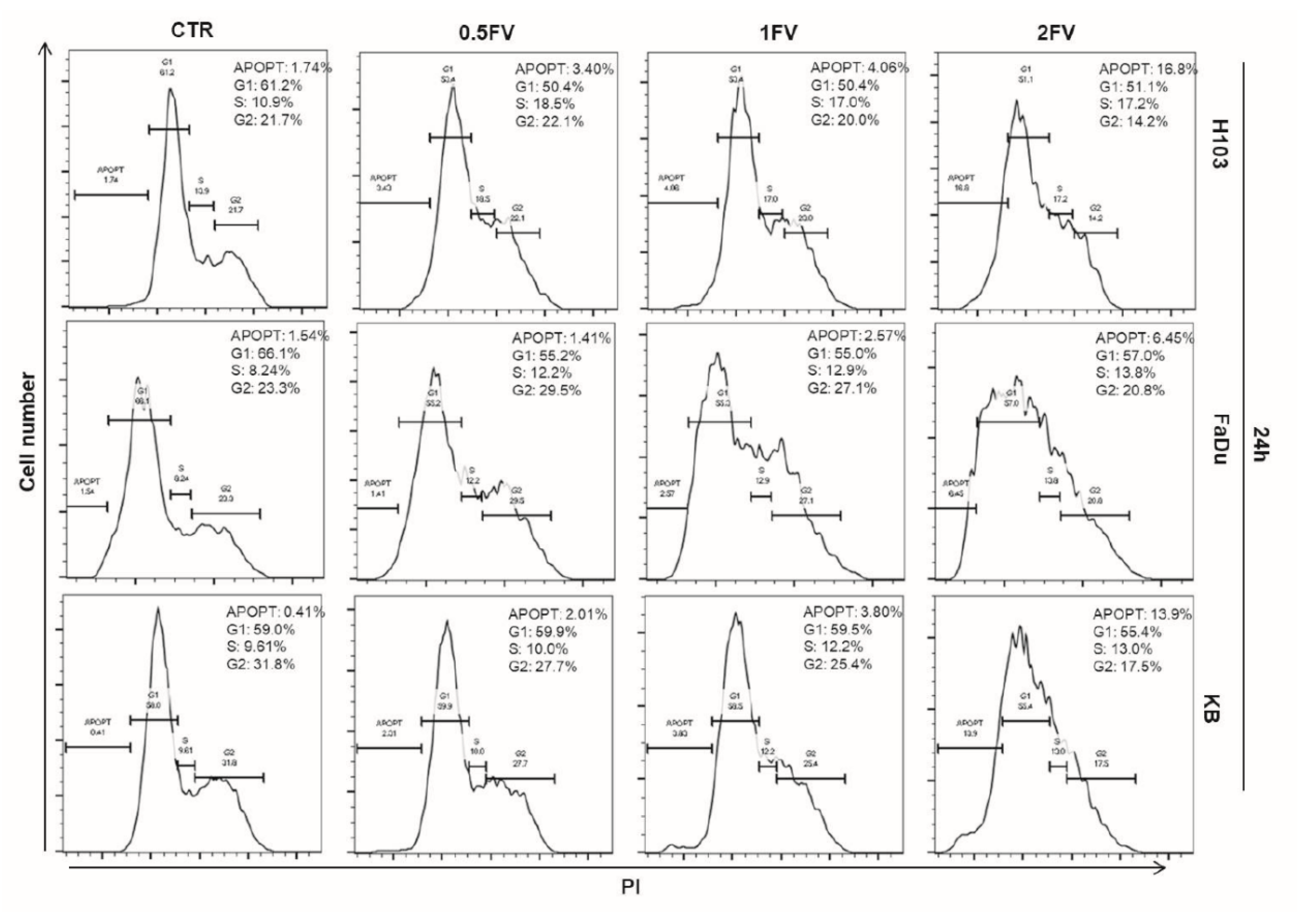
2.2. Treatment With Fucoidan Leads to Cell Cycle Perturbations Causing Arrest in G2/S or G1 Phase in HNSCC Cells
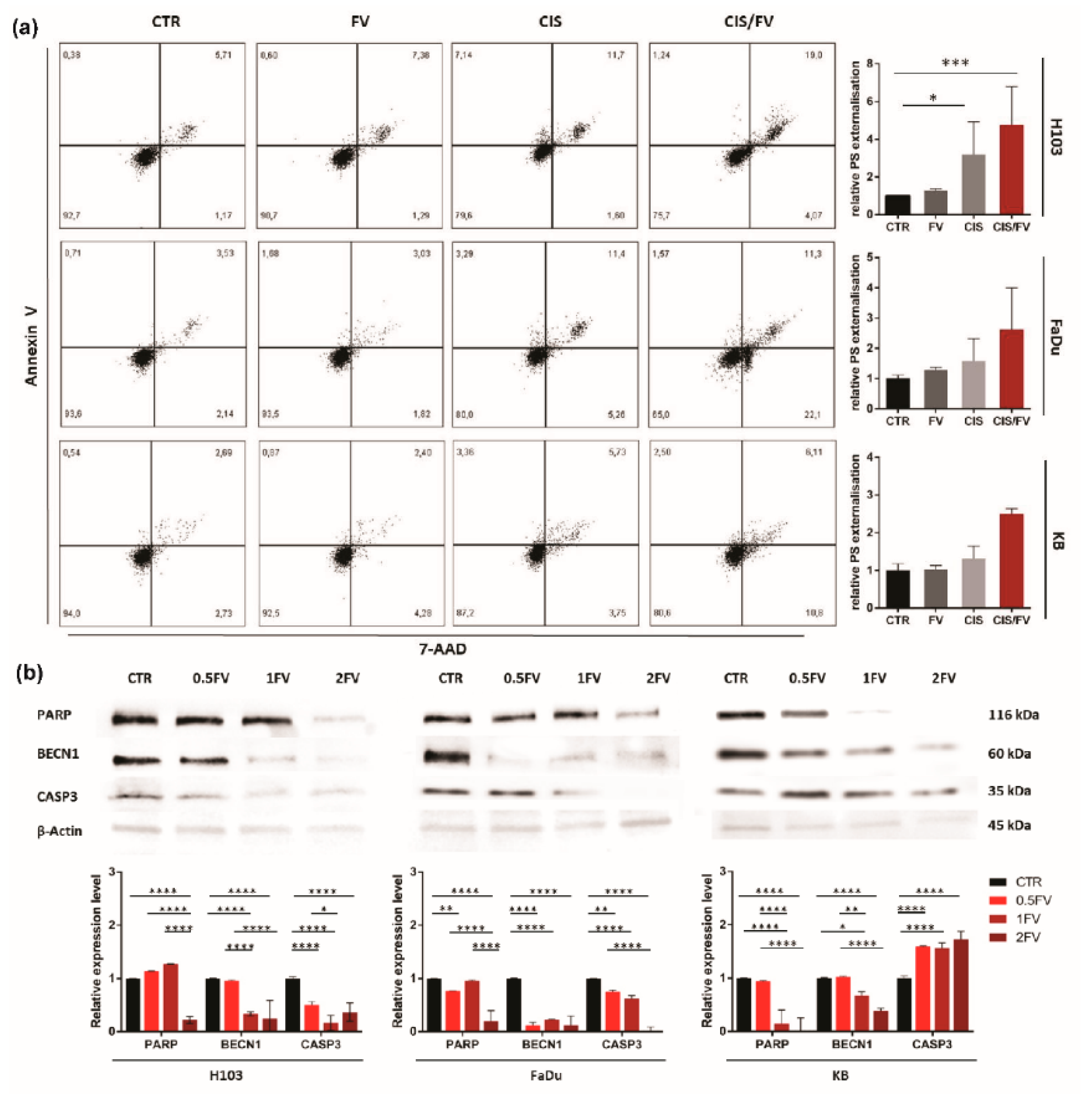
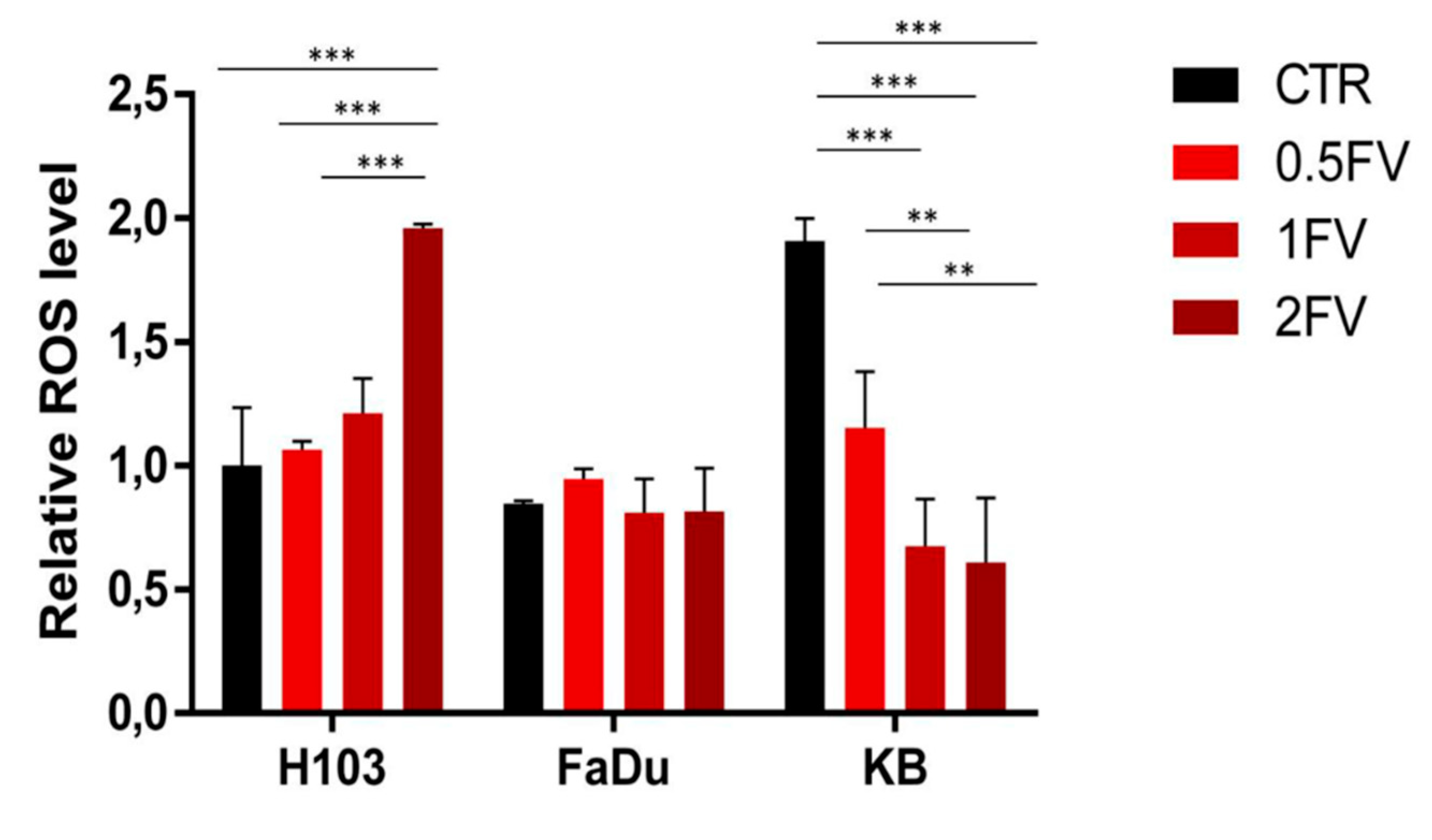
2.3. Fucoidan Induces Apoptosis of HNSCC Cells Due to the Regulation of ROS Production
2.4. FV Enhances Response to Cisplatin
3. Discussion
4. Materials and Methods
4.1. Fucoidan and Cisplatin
4.2. Cell Culture
4.3. MTT Assay
4.4. Flow Cytometry
4.4.1. Cell Cycle Analysis
4.4.2. Apoptosis Analysis
4.4.3. ROS Production Analysis
4.5. Western Blot
4.6. Analysis of Drug Combinatory Effect
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Berteau, O.; Mulloy, B. Sulfated fucans, fresh perspectives: Structures, functions, and biological properties of sulfated fucans and an overview of enzymes active toward this class of polysaccharide. Glycobiology 2003, 13, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Blaszczak, W.; Sobecka, A.; Barczak, W.; Suchorska, W.M. Fucoidan and its potential as a complementary agent in cancer therapy. Lett. Oncol. Sci. 2017, 14, 86–95. [Google Scholar]
- Sanjeewa, K.K.A.; Lee, J.S.; Kim, W.S.; Jeon, Y.J. The potential of brown-algae polysaccharides for the development of anticancer agents: An update on anticancer effects reported for fucoidan and laminaran. Carbohydr. Polym. 2017, 177, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Han, M.H.; Lee, D.S.; Jeong, J.W.; Hong, S.H.; Choi, I.W.; Cha, H.J.; Kim, S.; Kim, H.S.; Park, C.; Kim, G.Y.; et al. Fucoidan Induces ROS-Dependent Apoptosis in 5637 Human Bladder Cancer Cells by Downregulating Telomerase Activity via Inactivation of the PI3K/Akt Signaling Pathway. Drug Dev. Res. 2016, 78, 37–48. [Google Scholar] [CrossRef]
- Liu, F.; Luo, G.; Xiao, Q.; Chen, L.; Luo, X.; Lv, J.; Chen, L. Fucoidan inhibits angiogenesis induced by multiple myeloma cells. Oncol. Rep. 2016, 36, 1963–1972. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, T.; Hayakawa, K.; Kusakabe, T.; Takada, H.; Nakazato, K.; Hisanaga, E.; Iha, M. Inhibitory effect of fucoidan on Huh7 hepatoma cells through downregulation of CXCL12. Nutr. Cancer 2009, 61, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Kuo, C.H.; Chen, P.W. Compressional-puffing pretreatment enhances neuroprotective effects of fucoidans from the brown seaweed sargassum hemiphyllum on 6-hydroxydopamine-induced apoptosis in SH-SY5Y cells. Molecules 2018, 23, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-L.; Tai, C.-J.; Huang, C.-W.; Chang, F.-R.; Wang, J.-Y. Efficacy of Low-Molecular-Weight Fucoidan as a Supplemental Therapy in Metastatic Colorectal Cancer Patients: A Double-Blind Randomized Controlled Trial. Mar. Drugs 2017, 15, 122. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.; Lin, T.; Wu, Y.; Tsao, S. Fucoidan inhibition of lung cancer in vivo and in vitro? Role of the Smurf2-dependent ubiquitin proteasome pathway in TGFß receptor degradation. Oncotarget 2014, 5, 7870–7885. [Google Scholar] [CrossRef]
- Xue, M.; Ge, Y.; Zhang, J.; Wang, Q.; Hou, L.; Liu, Y.; Sun, L.; Li, Q. Anticancer properties and mechanisms of fucoidan on mouse breast cancer in vitro and in Vivo. PLoS ONE 2012, 7, 3–11. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Miller, K.D.; Ahmedin, J. Cáncer Statistics. Ca Cáncer J. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Heck, J.E.; Berthiller, J.; Vaccarella, S.; Winn, D.M.; Smith, E.M.; Shan’gina, O.; Schwartz, S.M.; Purdue, M.P.; Pilarska, A.; Eluf-Neto, J.E.; et al. Sexual behaviours and the risk of head and neck cancers: A pooled analysis in the International Head and Neck Cancer Epidemiology (INHANCE) consortium. Int. J. Epidemiol. 2010, 39, 166–181. [Google Scholar] [CrossRef] [PubMed]
- Hashibe, M.; Brennan, P.; Chuang, S.C.; Boccia, S.; Castellsague, X.; Chen, C.; Curado, M.P.; Dal Maso, L.; Daudt, A.W.; Fabianova, E.; et al. Interaction between tobacco and alcohol use and the risk of head and neck cancer: Pooled analysis in the International Head and Neck Cancer Epidemiology Consortium. Cancer Epidemiol. Biomark. Prev. 2009, 18, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Elting, L.S.; Cooksley, C.; Chambers, M.; Cantor, S.B.; Manzullo, E.; Rubenstein, E.B. The burdens of cancer therapy: Clinical and economic outcomes of chemotherapy-induced mucositis. Cancer 2003, 98, 1531–1539. [Google Scholar] [CrossRef] [PubMed]
- Blaszczak, W.; Barczak, W.; Wegner, A.; Golusinski, W.; Suchorska, W.M. Clinical value of monoclonal antibodies and tyrosine kinase inhibitors in the treatment of head and neck squamous cell carcinoma. Med. Oncol. 2017, 34, 60. [Google Scholar] [CrossRef]
- Tesniere, A.; Panaretakis, T.; Kepp, O.; Apetoh, L.; Ghiringhelli, F.; Zitvogel, L.; Kroemer, G. Molecular characteristics of immunogenic cancer cell death. Cell Death Differ. 2008, 15, 3–12. [Google Scholar] [CrossRef]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef]
- Hyun, J.-H.; Kim, S.-C.; Kang, J.-I.; Kim, M.-K.; Boo, H.-J.; Kwon, J.-M.; Koh, Y.-S.; Hyun, J.-W.; Park, D.-B.; Yoo, E.-S.; et al. Apoptosis inducing activity of fucoidan in HCT-15 colon carcinoma cells. Biol. Pharm. Bull. 2009, 32, 1760–1764. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Shapiro, G.I.; Harper, J.W. Anticancer drug targets: Cell cycle and checkpoint control. J. Clin. Invest. 1999, 104, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Yan, M. De; Yao, C.J.; Chow, J.M.; Chang, C.L.; Hwang, P.A.; Chuang, S.E.; Whang-Peng, J.; Lai, G.M. Fucoidan elevates MicroRNA-29b to regulate DNMT3B-MTSS1 axis and inhibit EMT in human hepatocellular carcinoma cells. Mar. Drugs 2015, 13, 6099–6116. [Google Scholar] [CrossRef] [PubMed]
- Banafa, A.M.; Roshan, S.; Liu, Y.Y.; Chen, H.J.; Chen, M.J.; Yang, G.X.; He, G.Y. Fucoidan induces G1 phase arrest and apoptosis through caspases-dependent pathway and ROS induction in human breast cancer MCF-7 cells. J. Huazhong Univ. Sci. Technol.-Med. Sci. 2013, 33, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-S.; Lee, J.H.; Lee, S.H. Antitumor Effects of Fucoidan on Human Colon Cancer Cells via Activation of Akt Signaling. Biomol. Ther. (Seoul) 2015, 23, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Hu, A.; Huang, J.-J.; Zhang, J.-F.; Dai, W.-J.; Li, R.-L.; Lu, Z.-Y.; Duan, J.-L.; Li, J.-P.; Chen, X.-P.; Fan, J.-P.; et al. Curcumin induces G2/M cell cycle arrest and apoptosis of head and neck squamous cell carcinoma in vitro and in vivo through ATM/Chk2/p53-dependent pathway. Oncotarget 2017, 8, 50747–50760. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Xiao, Q.; Wang, L.; Liu, X.; Wang, X.; Yang, Z.; Zhang, H.; Dong, P. Fucoidan prevents multiple myeloma cell escape from chemotherapy-induced drug cytotoxicity. Fitoterapia 2013, 84, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Aisa, Y.; Miyakawa, Y.; Nakazato, T.; Shibata, H.; Saito, K.; Ikeda, Y.; Kizaki, M. Fucoidan induces apoptosis of human HS-Sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am. J. Hematol. 2005, 78, 7–14. [Google Scholar] [CrossRef]
- Kim, E.J.; Park, S.Y.; Lee, J.-Y.; Park, J.H.Y. Fucoidan present in brown algae induces apoptosis of human colon cancer cells. BMC Gastroenterol. 2010, 10, 96. [Google Scholar] [CrossRef]
- Park, H.; Kim, G.-Y.; Moon, S.-K.; Kim, W.; Yoo, Y.; Choi, Y. Fucoidan Inhibits the Proliferation of Human Urinary Bladder Cancer T24 Cells by Blocking Cell Cycle Progression and Inducing Apoptosis. Molecules 2014, 19, 5981–5998. [Google Scholar] [CrossRef]
- Mathew, L.; Burney, M.; Gaikwad, A.; Nyshadham, P.; Nugent, E.K.; Gonzalez, A.; Smith, J.A. Preclinical Evaluation of Safety of Fucoidan Extracts from Undaria pinnatifida and Fucus vesiculosus for Use in Cancer Treatment. Integr. Cancer Ther. 2017, 16, 572–584. [Google Scholar] [CrossRef]
- Dithmer, M.; Kirsch, A.M.; Richert, E.; Fuchs, S.; Wang, F.; Schmidt, H.; Coupland, S.E.; Roider, J.; Klettner, A. Fucoidan does not exert anti-tumorigenic effects on uveal melanoma cell lines. Mar. Drugs 2017, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.Y.; Lin, T.Y.; Hu, C.H.; Shu, D.T.F.; Lu, M.K. Fucoidan upregulates TLR4/CHOP-mediated caspase-3 and PARP activation to enhance cisplatin-induced cytotoxicity in human lung cancer cells. Cancer Lett. 2018, 432, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, S.; Synytsya, A.; Capek, P.; Lee, C.W.; Choi, J.W.; Cho, S.; Kim, W.J.; Park, Y. Il Low Molecular Weight Mannogalactofucans Derived from Undaria pinnatifida Induce Apoptotic Death of Human Prostate Cancer Cells In Vitro and In Vivo. Mar. Biotechnol. 2018, 20, 813. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.Y. Fucoidan as a marine anticancer agent in preclinical development. Mar. Drugs 2014, 12, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Roshan, S.; Liu, Y.Y.; Banafa, A.; Chen, H.J.; Li, K.X.; Yang, G.X.; He, G.Y.; Chen, M.J. Fucoidan induces apoptosis of HepG2 cells by down-regulating p-Stat3. J. Huazhong Univ. Sci. Technol.-Med. Sci. 2014, 34, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.; Lin, T.; Lu, M.; Leng, P.; Tsao, S. Fucoidan induces Toll-like receptor 4-regulated reactive oxygen species and promotes endoplasmic reticulum stress-mediated apoptosis in lung cancer. Nat. Publ. Gr. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Chen, M.-C.; Hsu, W.-L.; Hwang, P.-A.; Chou, T.-C. Low Molecular Weight Fucoidan Inhibits Tumor Angiogenesis through Downregulation of HIF-1/VEGF Signaling under Hypoxia. Mar. Drugs 2015, 13, 4436–4451. [Google Scholar] [CrossRef]
- Lai, D.; Tan, C.L.; Gunaratne, J.; Quek, L.S.; Nei, W.; Thierry, F.; Bellanger, S. Localization of HPV-18 E2 at Mitochondrial Membranes Induces ROS Release and Modulates Host Cell Metabolism. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef]
- Marullo, R.; Werner, E.; Zhang, H.; Chen, G.Z.; Shin, D.M.; Doetsch, P.W. HPV16 E6 and E7 proteins induce a chronic oxidative stress response via NOX2 that causes genomic instability and increased susceptibility to DNA damage in head and neck cancer cells. Carcinogenesis 2015, 36, 1397–1406. [Google Scholar] [CrossRef]
- Marur, S.; Forastiere, A. Head and neck cancer: Changing epidemiology, diagnosis, and treatment. Mayo Clin. Proc. 2008, 83, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Stokes, W.A.; Sumner, W.A.; Breggren, K.L.; Rathbun, J.T.; Raben, D.; McDermott, J.D.; Gan, G.; Karam, S.D. A comparison of concurrent cisplatin versus cetuximab with radiotherapy in locally-advanced head and neck cancer: A bi-institutional analysis. Rep. Pract. Oncol. Radiother. 2017, 22, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Dirix, P.; Nuyts, S. Evidence-based organ-sparing radiotherapy in head and neck cancer. Lancet Oncol. 2010, 11, 85–91. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Wu, T.T.; Zhou, S.H.; Bao, Y.Y.; Wang, Q.Y.; Fan, J.; Huang, Y.P. Apigenin suppresses GLUT-1 and p-AKT expression to enhance the chemosensitivity to cisplatin of laryngeal carcinoma Hep-2 cells: An In vitro study. Int. J. Clin. Exp. Pathol. 2014, 7, 3938–3947. [Google Scholar]
- Varoni, E.M.; Lo Faro, A.F.; Sharifi-Rad, J.; Iriti, M. Anticancer Molecular Mechanisms of Resveratrol. Front. Nutr. 2016, 3. [Google Scholar] [CrossRef]
- Kma, L. Roles of Plant Extracts and Constituents in Cervical Cancer Therapy. Asian Pac. J. Cancer Prev. 2013, 14, 3429–3436. [Google Scholar] [CrossRef]
- Zhang, Z.; Teruya, K.; Yoshida, T.; Eto, H.; Shirahata, S. Fucoidan extract enhances the anti-cancer activity of chemotherapeutic agents in MDA-MB-231 and MCF-7 breast cancer cells. Mar. Drugs 2013, 11, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Abudabbus, A.; Badmus, J.A.; Shalaweh, S.; Bauer, R.; Hiss, D. Effects of Fucoidan and Chemotherapeutic Agent Combinations on Malignant and Non-malignant Breast Cell Lines. Curr. Pharm. Biotechnol. 2017, 18, 748–757. [Google Scholar] [CrossRef]
- Thakur, V.; Lu, J.; Roscilli, G.; Aurisicchio, L.; Cappelletti, M. The natural compound fucoidan from New Zealand Undaria pinnatifida synergizes with the ERBB inhibitor lapatinib enhancing melanoma growth inhibition Fucoidan extracted from New Zealand. Oncotarget 2017, 8, 7887–7896. [Google Scholar] [CrossRef]
- Schrader, C.; Boehm, A.; Reiche, A.; Diet, A.; Mozet, C.; Wichmann, G. Combined effects of lapatinib and cisplatin on colony formation of head and neck squamous cell carcinoma. Anticancer Res. 2012, 32, 3191–3199. [Google Scholar]
- Stoehr, M.; Mozet, C.; Boehm, A.; Aigner, A.; Dietz, A.; Wichmann, G. Simvastatin suppresses head and neck squamous cell carcinoma ex vivo and enhances the cytostatic effects of chemotherapeutics. Cancer Chemother. Pharmacol. 2014, 73, 827–837. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the fucoidan are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Cell Line | q-Value | Combinatory Effect |
|---|---|---|---|
| MTT | H103 | 2.90 | Synergism |
| FaDu | 1.51 | Synergism | |
| KB | 1.24 | Synergism | |
| Apoptosis analysis | H103 | 1.19 | Synergism |
| FaDu | 0.94 | Additivity | |
| KB | 1.22 | Synergism |
| Cell Number | |||
|---|---|---|---|
| Incubation Time | H103 | FaDu | KB |
| 24 h | 2000 | 2500 | 3000 |
| 48 h | 1000 | 2000 | 1500 |
| 72 h | 750 | 1000 | 750 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blaszczak, W.; Lach, M.S.; Barczak, W.; Suchorska, W.M. Fucoidan Exerts Anticancer Effects Against Head and Neck Squamous Cell Carcinoma In Vitro. Molecules 2018, 23, 3302. https://doi.org/10.3390/molecules23123302
Blaszczak W, Lach MS, Barczak W, Suchorska WM. Fucoidan Exerts Anticancer Effects Against Head and Neck Squamous Cell Carcinoma In Vitro. Molecules. 2018; 23(12):3302. https://doi.org/10.3390/molecules23123302
Chicago/Turabian StyleBlaszczak, Wiktoria, Michal Stefan Lach, Wojciech Barczak, and Wiktoria Maria Suchorska. 2018. "Fucoidan Exerts Anticancer Effects Against Head and Neck Squamous Cell Carcinoma In Vitro" Molecules 23, no. 12: 3302. https://doi.org/10.3390/molecules23123302
APA StyleBlaszczak, W., Lach, M. S., Barczak, W., & Suchorska, W. M. (2018). Fucoidan Exerts Anticancer Effects Against Head and Neck Squamous Cell Carcinoma In Vitro. Molecules, 23(12), 3302. https://doi.org/10.3390/molecules23123302