The Prebiotic Activity of Simulated Gastric and Intestinal Digesta of Polysaccharides from the Hericium erinaceus

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Analysis of HEP
2.2. Prebiotic Activity
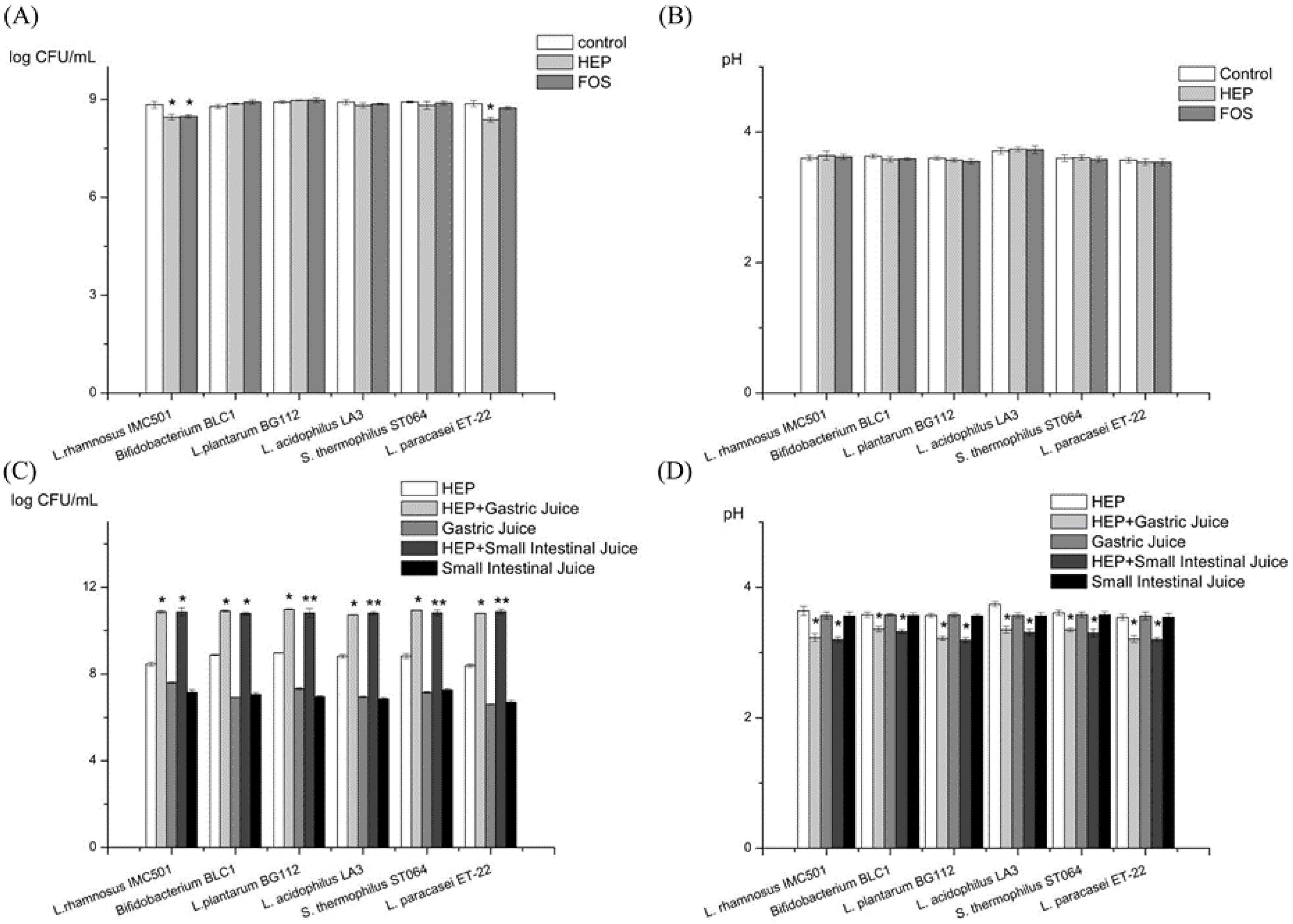
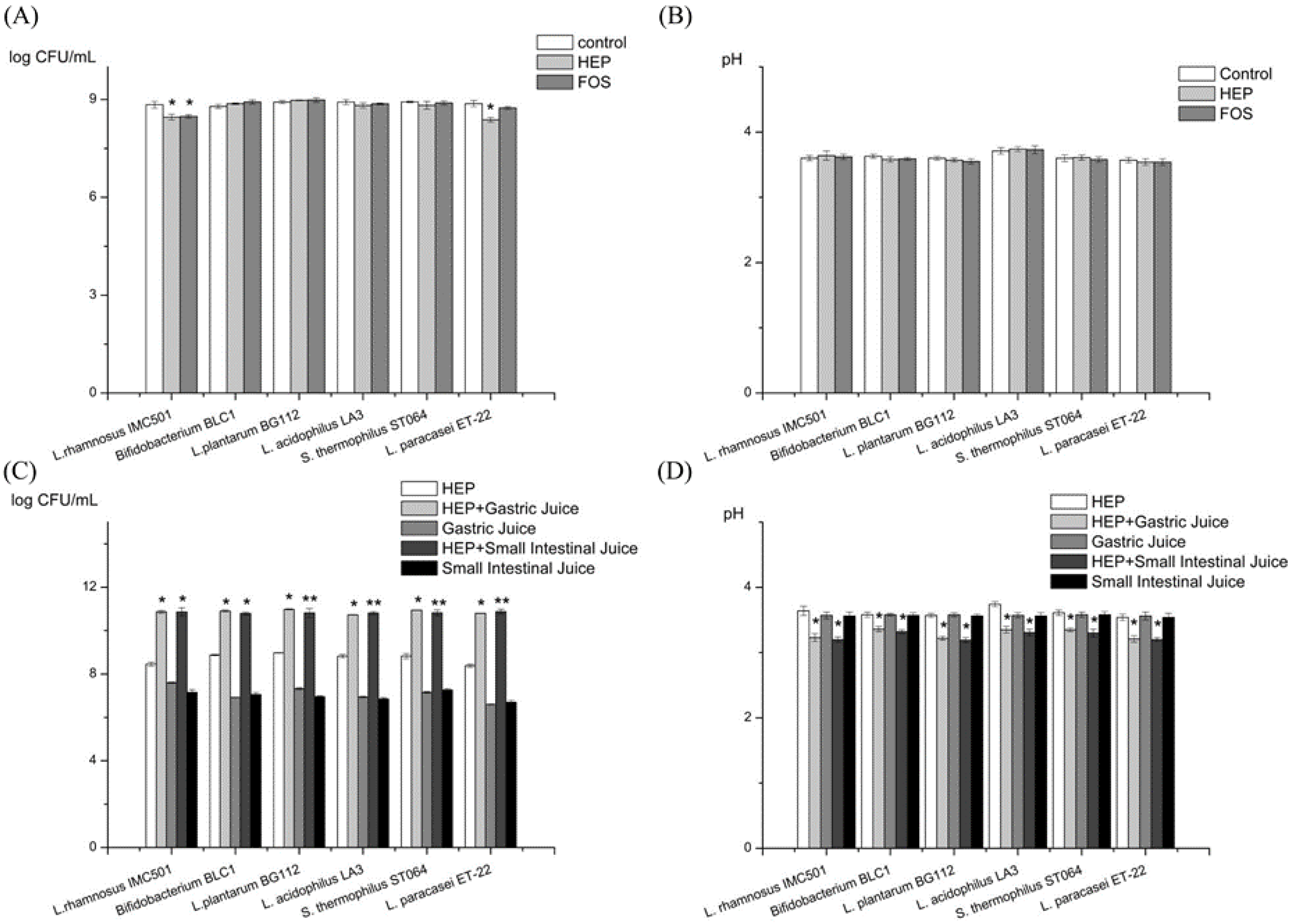
2.2.1. Prebiotic Activity of HEP
2.2.2. Prebiotic Activity of the Gastric and the Intestinal Digesta of HEP
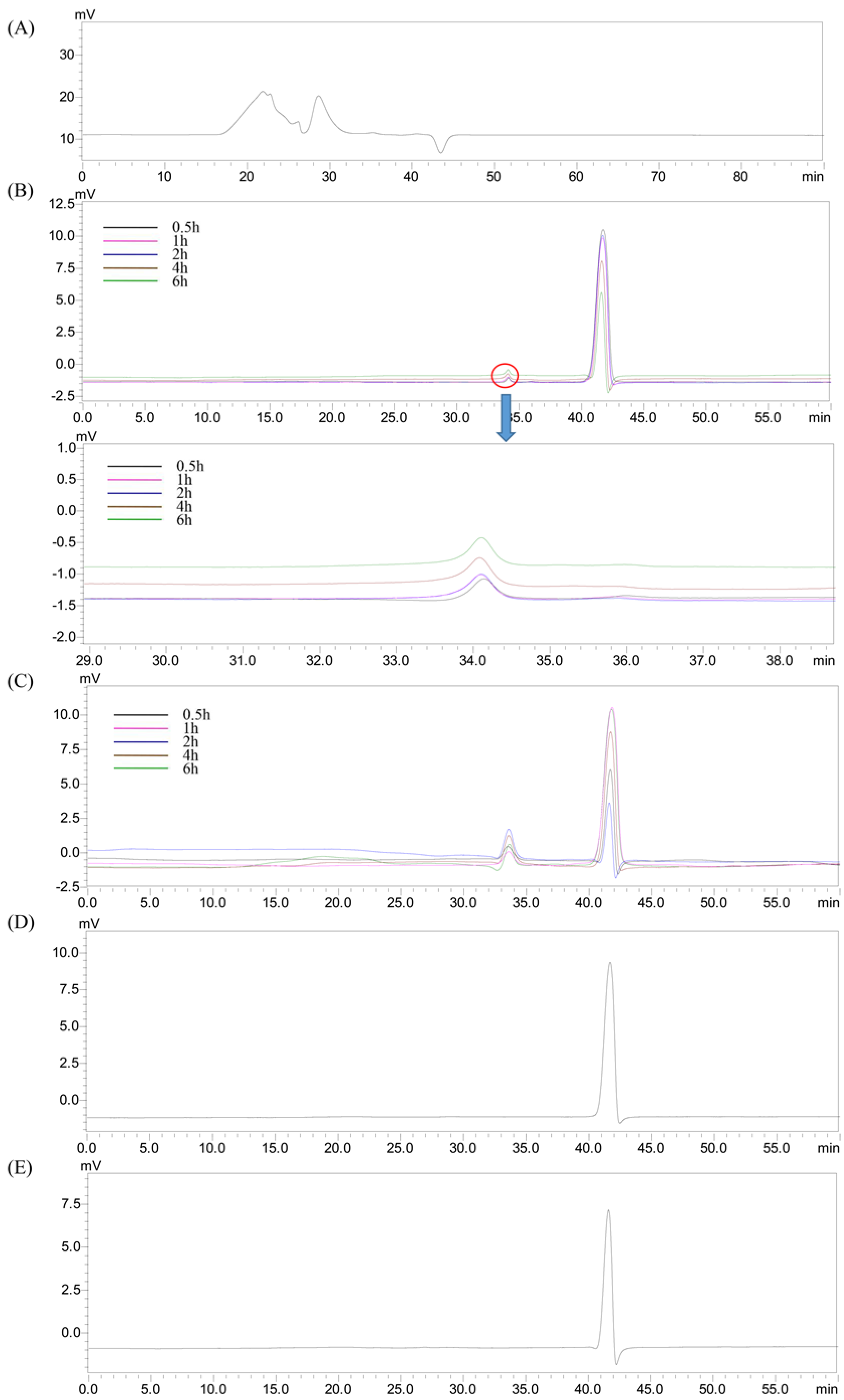
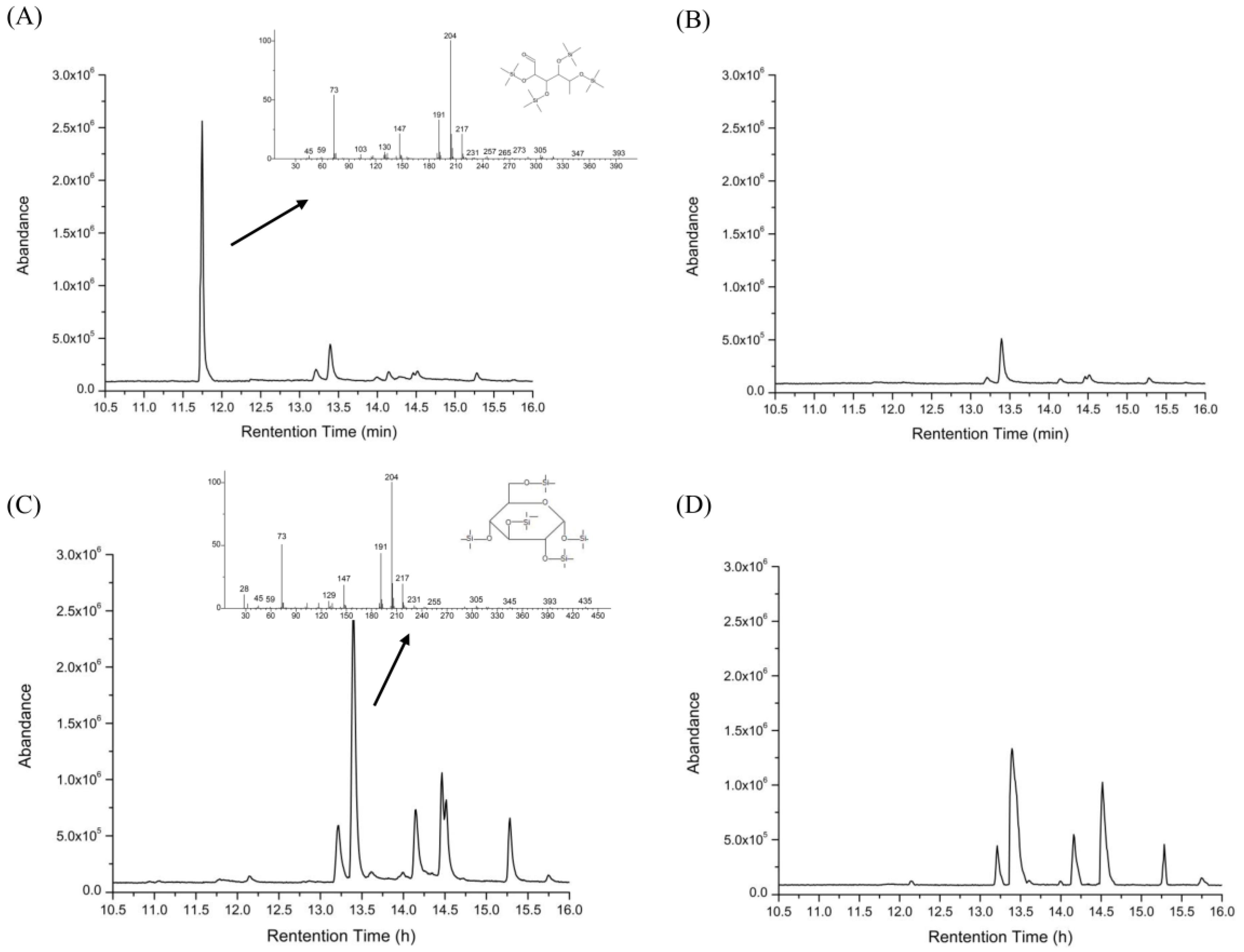
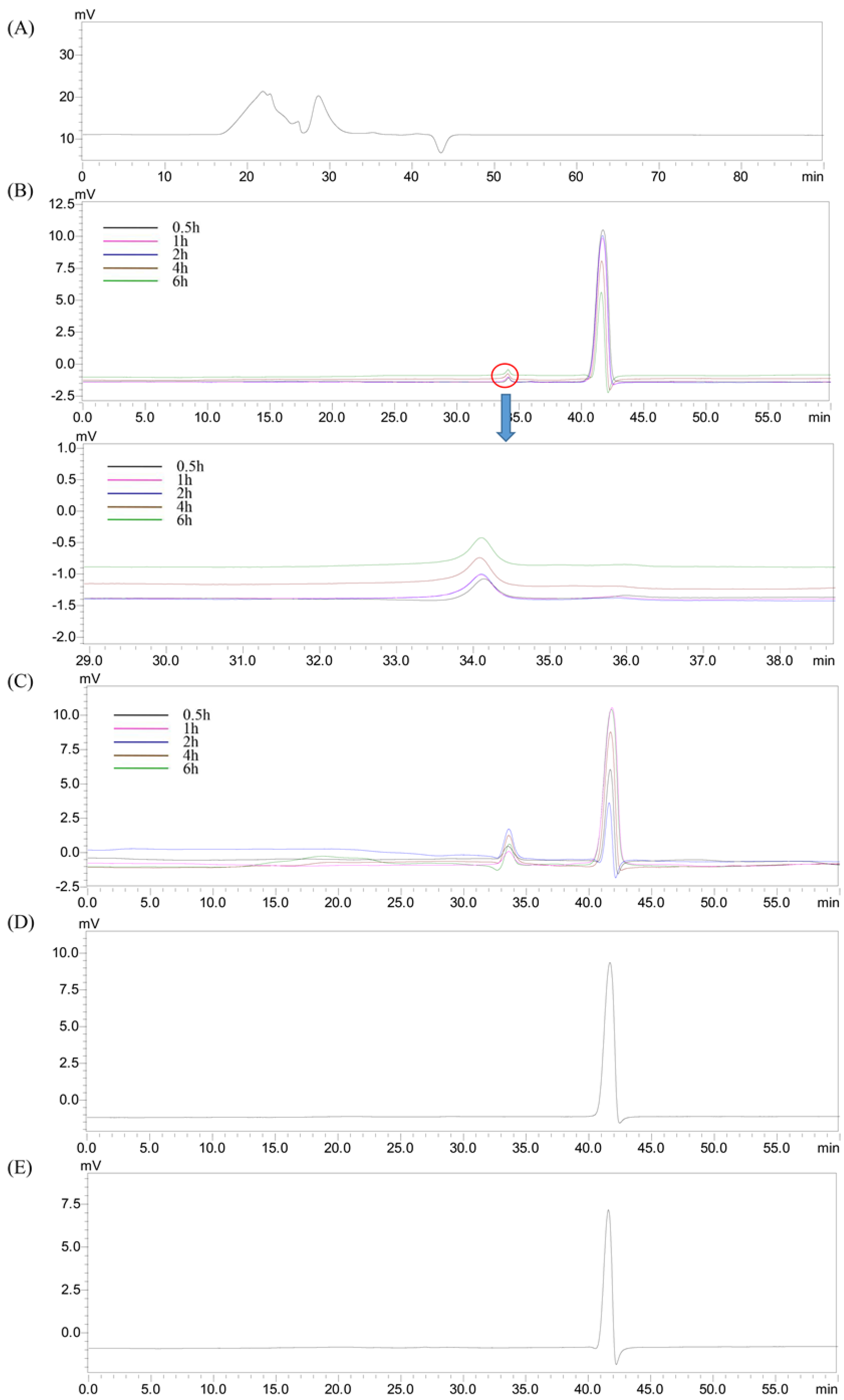
2.3. Changes in the Molecular Weight of HEP after Digestion
2.4. Changes of Reducing Sugars After Gastric and Intestinal Digestion
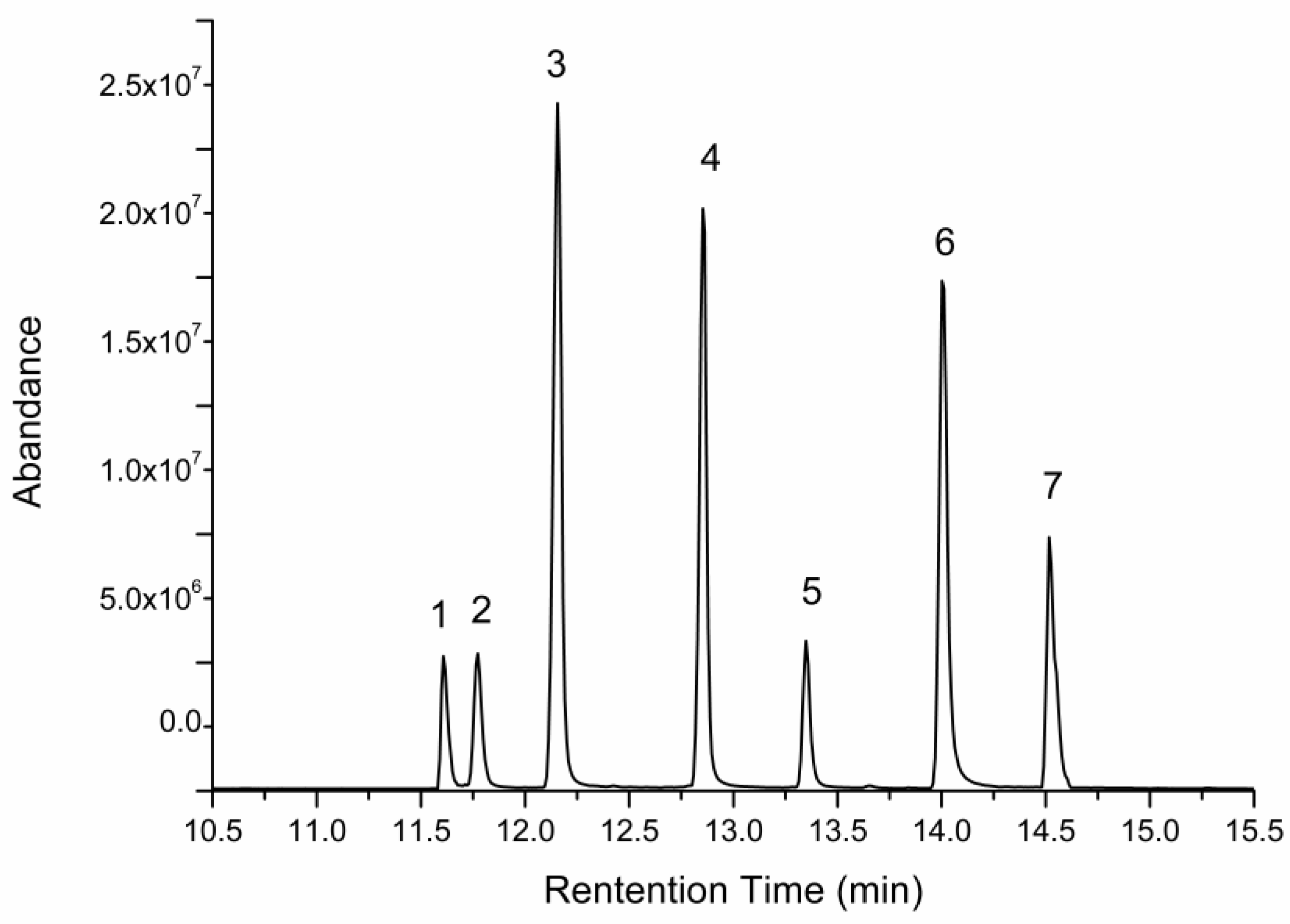
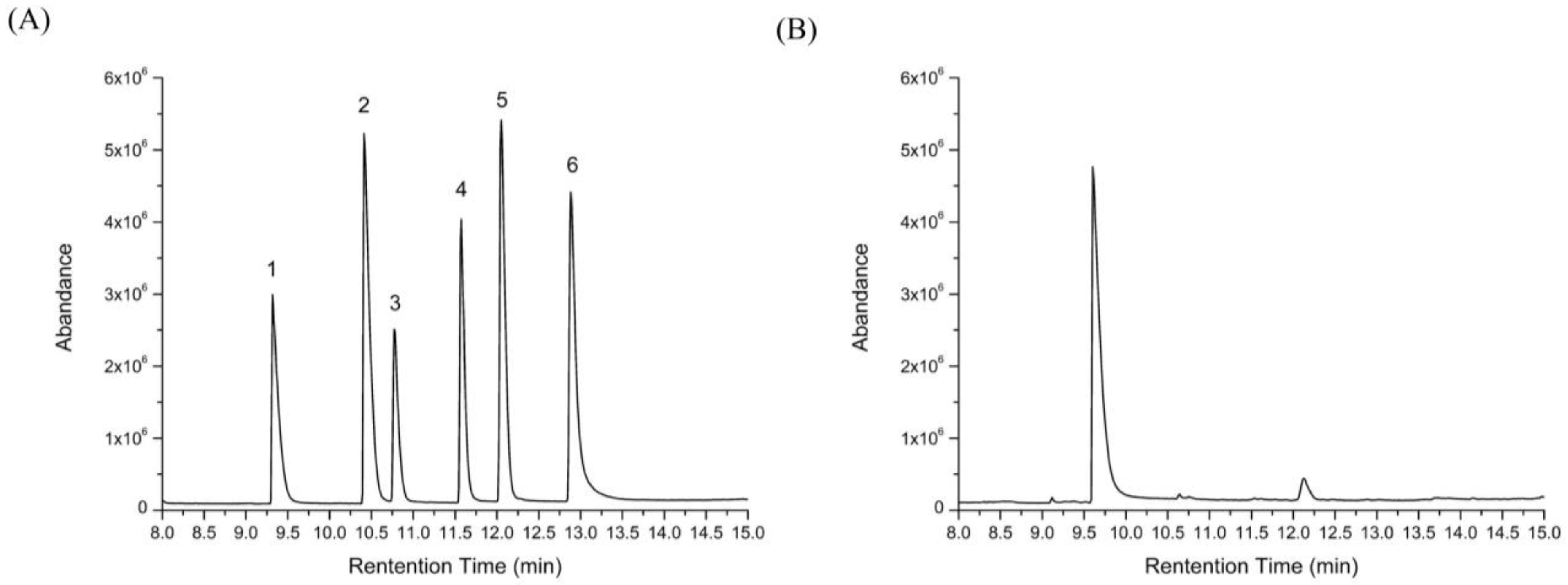
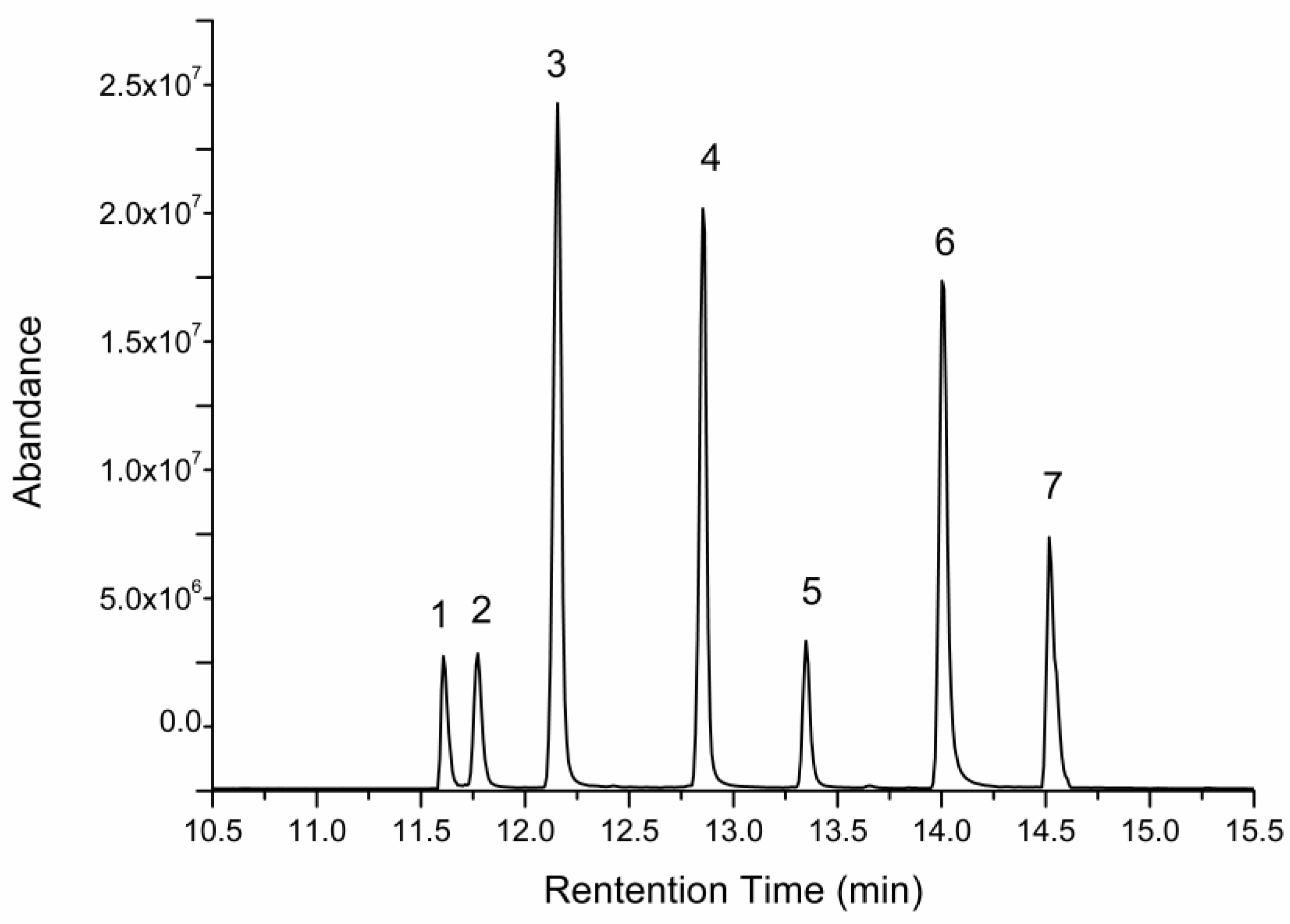
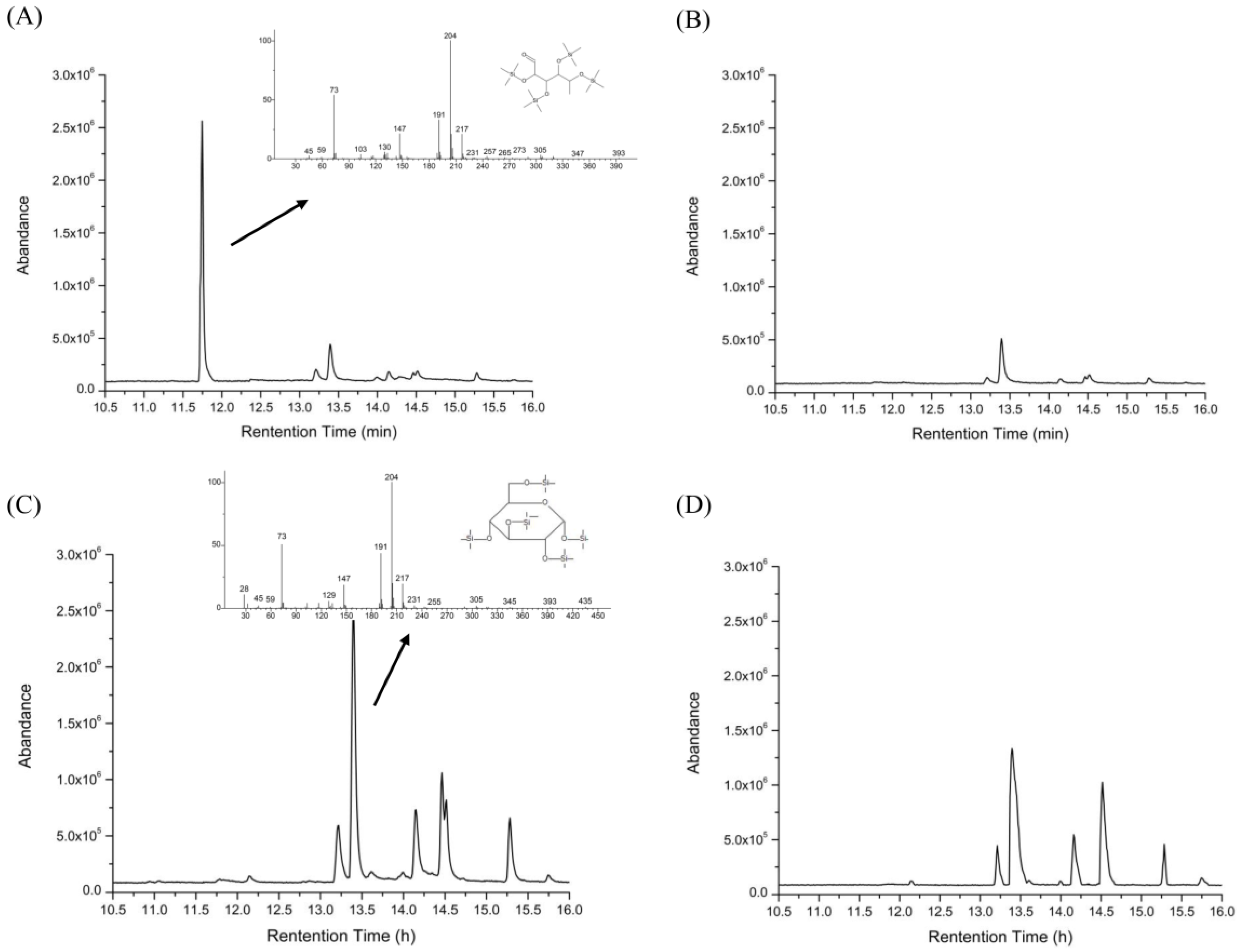
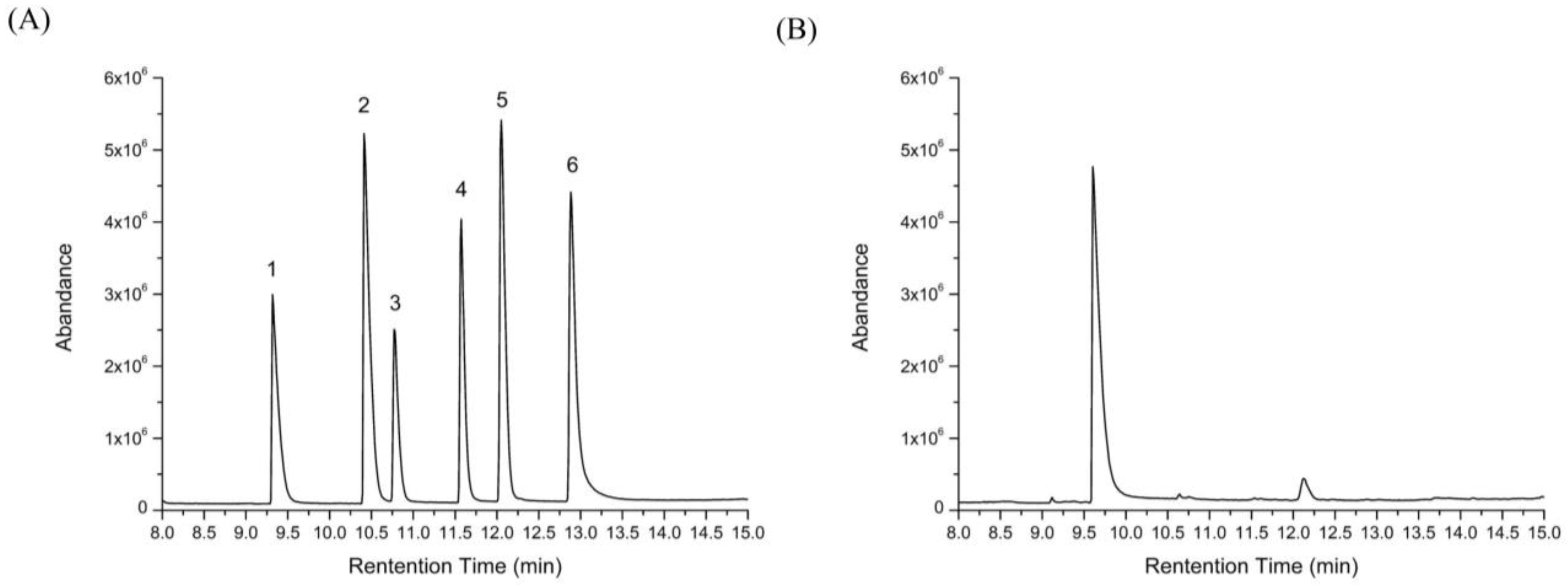
2.5. Changes of Free Monosaccharides after Gastric and Intestinal Digestion
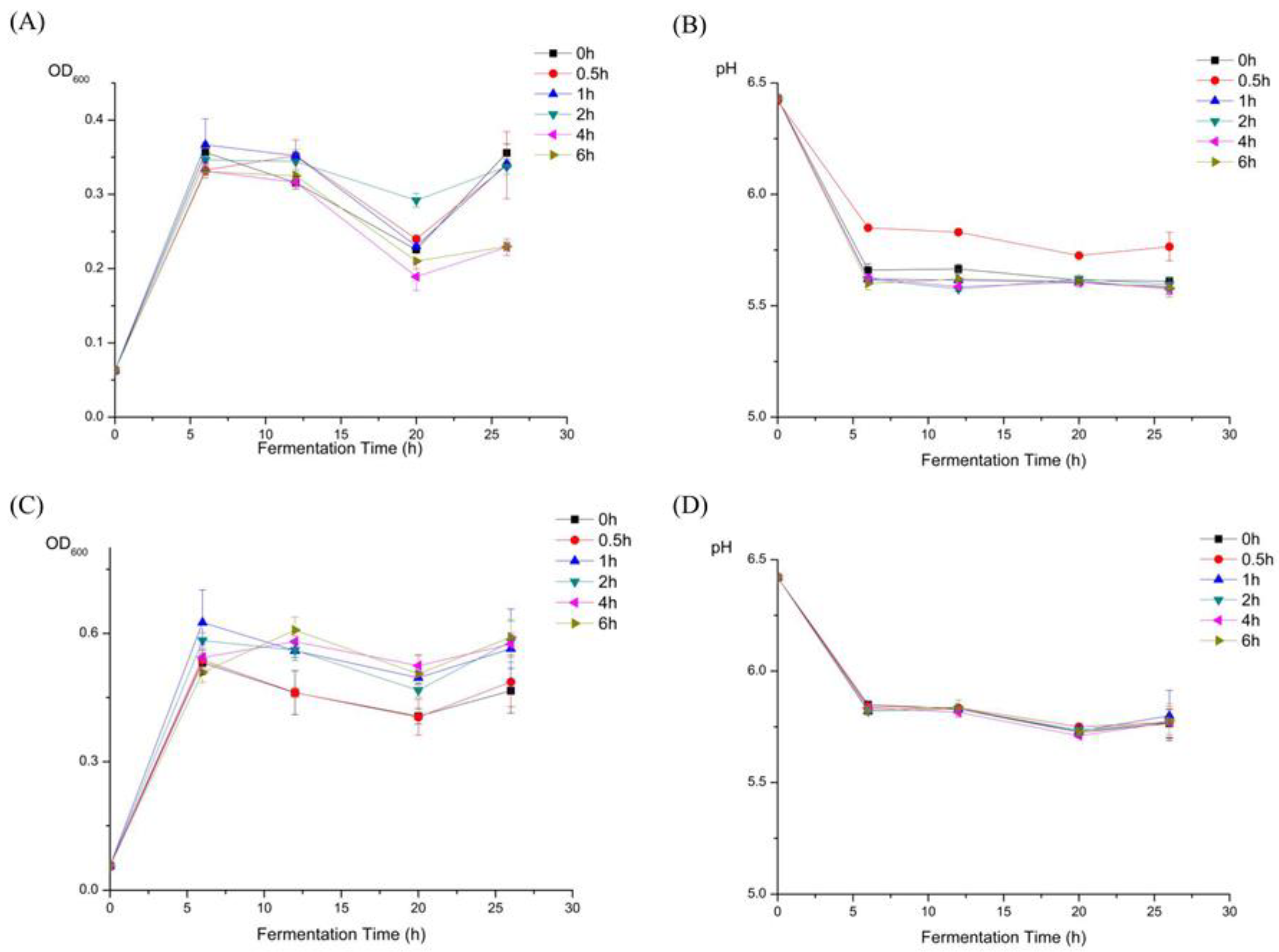
2.6. Changes in the Molecular Weight of HEP after Fermentation
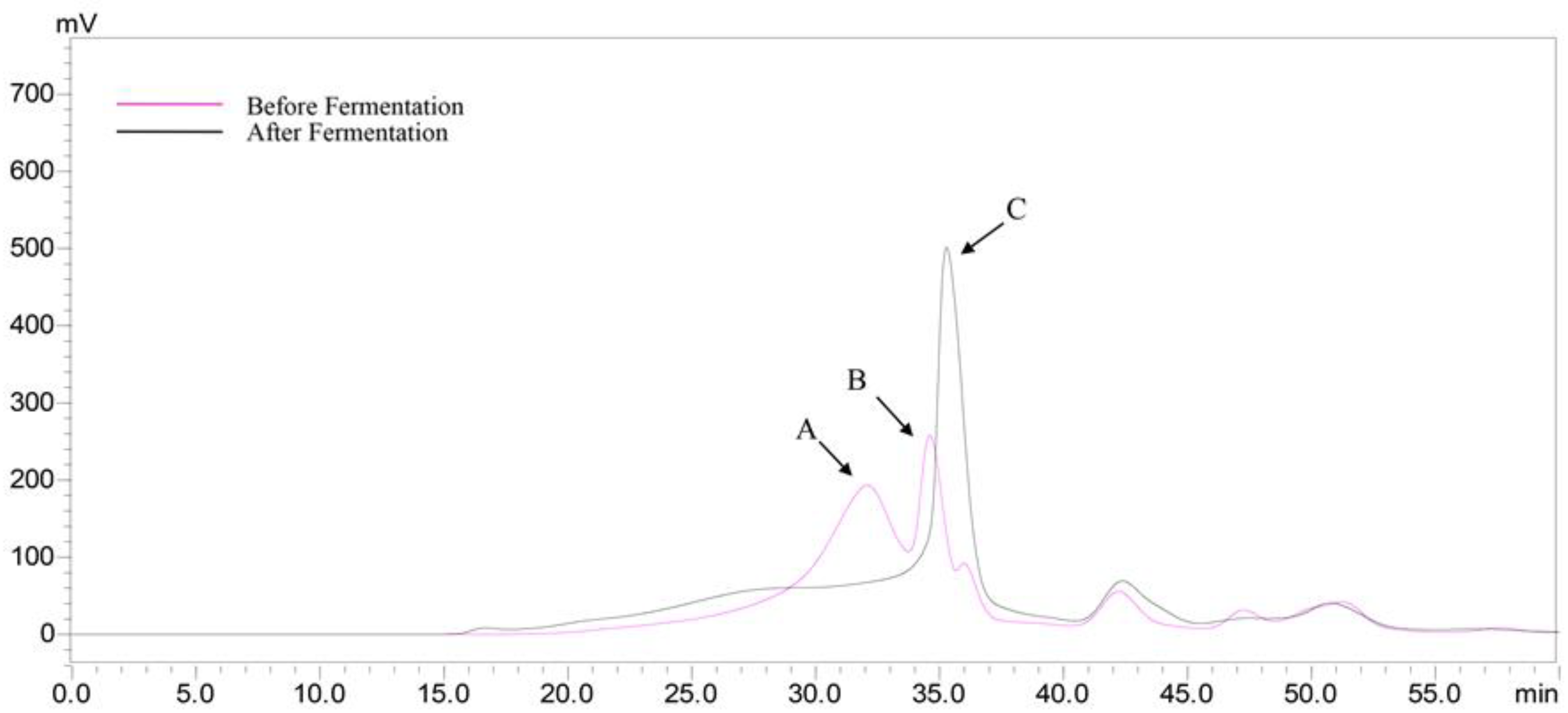
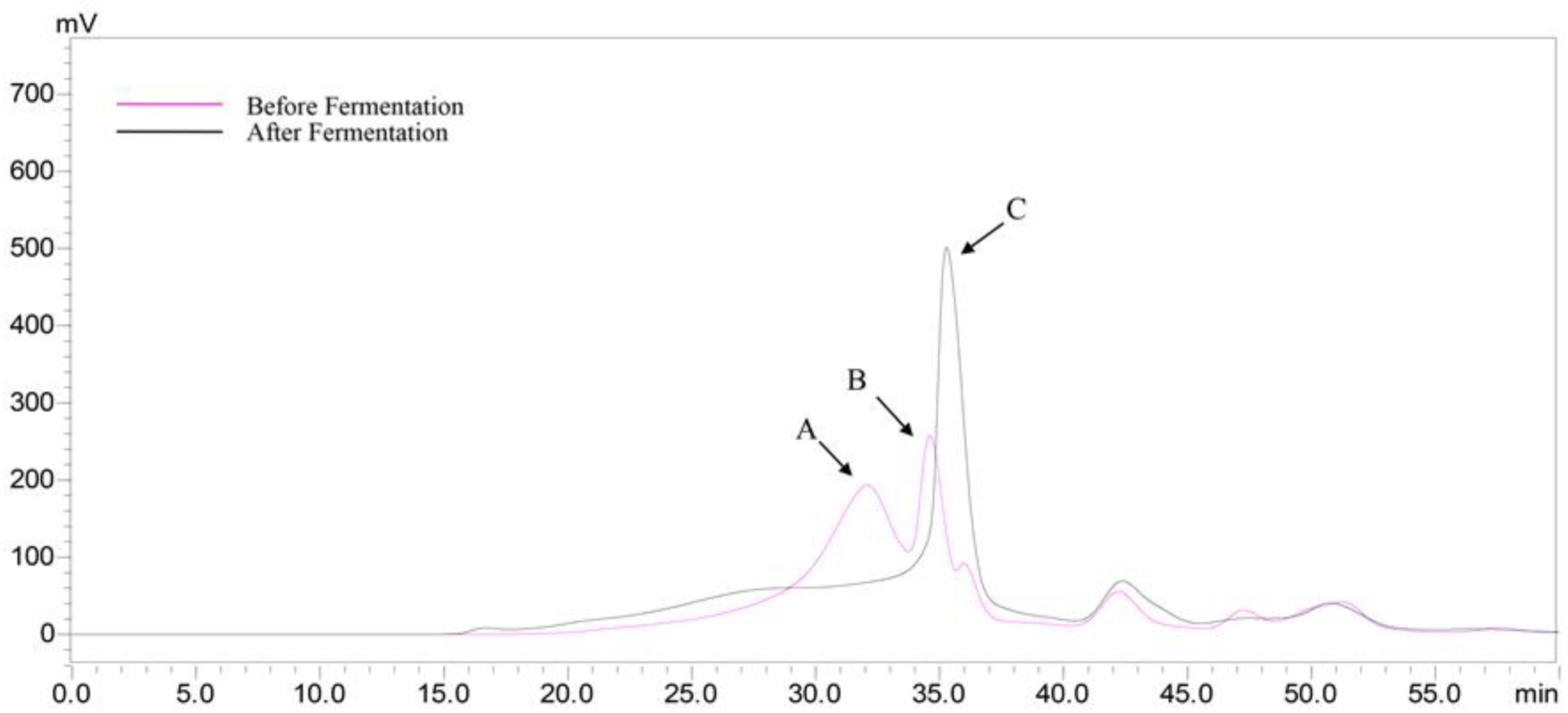
2.7. Production of Short Chain Fatty Acids (SCFAs) after Fermentation
3. Materials and Methods
3.1. Materials
3.2. Isolation and Purification of HEP
3.3. Composition Analysis
3.4. Prebiotic Activity
3.4.1. Rejuvenation of Microbial Strains
3.4.2. Culture of Microbial Strains
3.5. Simulated Gastric Digestion In Vitro
3.6. Simulated Intestinal Digestion In Vitro
3.7. Determination of Molecular Weight
3.8. Determination of Reducing Sugar Content
3.9. Determination of Free Monosaccharides
3.10. Determination of Short-Chain Fatty Acids Production
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ko, H.G.; Park, H.G.; Park, S.H.; Choi, C.W.; Kim, S.H.; Park, W.M. Comparative study of mycelial growth and basidiomata formation in seven different species of the edible mushroom genus hericium. Bioresour. Technol. 2005, 96, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.Q.; Fu, L.; Xu, M.; Sun, P.L.; Zhang, J.S. Structure of a water-soluble heteropolysaccharide from fruiting bodies of hericium erinaceus. Carbohydr. Polym. 2012, 88, 558–561. [Google Scholar] [CrossRef]
- Yang, J.H.; Lin, H.C.; Mau, J.L. Antioxidant properties of several commercial mushrooms. Food Chem. 2002, 35, 519–526. [Google Scholar] [CrossRef]
- Mizuno, T.; Saito, H.; Nishitoba, T.; Kawagishi, H. Antitumor-active substances from mushrooms. Food Rev. Intern. 1995, 11, 23–61. [Google Scholar] [CrossRef]
- Yeh, S.P.; Hsia, L.F.; Chiu, C.S.; Chiu, S.T.; Liu, C.H. A smaller particle size improved the oral bioavailability of monkey head mushroom, hericium erinaceum, powder resulting in enhancement of the immune response and disease resistance of white shrimp, litopenaeus vannamei. Fish Shellfish Immunol. 2011, 30, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.M. Isolation of antimicrobial substances from. Mycobiology 2000, 28, 33–38. [Google Scholar] [CrossRef]
- Wang, J.C.; Hu, S.H.; Wang, J.T.; Chen, K.S.; Chia, Y.C. Hypoglycemic effect of extract of hericium erinaceus. J. Sci. Food Agr. 2010, 85, 641–646. [Google Scholar] [CrossRef]
- Hinsberger, A.; Sandhu, B.K. Digestion and absorption. Cur. Paediatrics 2004, 24, 250–254. [Google Scholar] [CrossRef]
- Pedersen, A.M.; Bardow, A.; Jensen, S.B.; Nauntofte, B. Saliva and gastrointestinal functions of taste, mastication, swallowing and digestion. Oral Dis. 2010, 8, 117–129. [Google Scholar] [CrossRef]
- Pinto, J.; Spã-Nola, V.; Llorent-Martã-Nez, E.J.; Ml, N.D.C.R.; Molina-Garcã-A., L.; Castilho, P.C. Polyphenolic profile and antioxidant activities of madeiran elderberry (sambucus lanceolata) as affected by simulated in vitro digestion. Food Res. Int. 2017, 100, 404–410. [Google Scholar] [CrossRef] [PubMed]
- 1Antonio, C.; Antonio, G.; Franciscoa, T.; Juancarlos, E.; Reyes, B. Availability of polyphenols in fruit beverages subjected to in vitro gastrointestinal digestion and their effects on proliferation, cell-cycle and apoptosis in human colon cancer caco-2 cells. Food Chem. 2009, 114, 813–820. [Google Scholar]
- Hu, J.L.; Nie, S.P.; Min, F.F.; Xie, M.Y. Artificial simulated saliva, gastric and intestinal digestion of polysaccharide from the seeds of plantago asiatica L. Carbohydr. Polym. 2013, 92, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, W.; Chen, Z.; Gao, X.; Yuan, G.; Pan, Y.; Chen, H. Effects of simulated gastrointestinal digestion in vitro on the chemical properties, antioxidant activity, α-amylase and α-glucosidase inhibitory activity of polysaccharides from inonotus obliquus. Food Res. Int. 2018, 103, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chen, G.; Wan, P.; Hu, B.; Chen, L.; Ou, S.; Zeng, X.; Ye, H. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro of polysaccharides from the flowers of camellia sinensis induced by human gut microbiota. Food Func. 2017, 8, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeir, J.; De, V.M. Probiotics, prebiotics, and synbiotics--approaching a definition. Am. J. Clin. Nutr. 2001, 73, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Jaskari, J.; Kontula, P.; Siitonen, A.; Jousimies-Somer, H.; Mattila-Sandholm, T.; Poutanen, K. Oat beta-glucan and xylan hydrolysates as selective substrates for bifidobacterium and lactobacillus strains. Appl. Microbiol. Biotechnol. 1998, 49, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Menne, E.; Guggenbuhl, N.; Roberfroid, M. Fn-type chicory inulin hydrolysate has a prebiotic effect in humans. J. Nutr. 2000, 130, 1197–1199. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.T.; Li, W.Z.; Chen, J.; Zhong, Q.X.; Ju, Y.J.; Zhao, J.; Bzhelyansky, A.; Li, S.P. An evaluation system for characterization of polysaccharides from the fruiting body of hericium erinaceus and identification of its commercial product. Carbohydr. Polym. 2015, 124, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gao, Y.; Xu, D.; Gao, Q. A polysaccharide from cultured mycelium of hericium erinaceus and its anti-chronic atrophic gastritis activity. Int. J. Biol. Macromol. 2015, 81, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Akbari-Alavijeh, S.; Soleimanian-Zad, S.; Sheikh-Zeinoddin, M.; Hashmi, S. Pistachio hull water-soluble polysaccharides as a novel prebiotic agent. Int. J. Biol. Macromol. 2017, 107, 808–816. [Google Scholar] [CrossRef] [PubMed]
- 2Azmi, A.F.; Mustafa, S.; Hashim, D.M.; Manap, Y.A. Prebiotic activity of polysaccharides extracted from Gigantochloa levis (buluh beting) shoots. Molecules 2012, 17, 1635–1651. [Google Scholar]
- Thambiraj, S.R.; Phillips, M.; Koyyalamudi, S.R.; Reddy, N. Yellow lupin (lupinus luteus L.) polysaccharides: Antioxidant, immunomodulatory and prebiotic activities and their structural characterisation. Food Chem. 2018, 267, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, B.; Fu, X.; You, L.J.; Abbasi, A.M.; Liu, R.H. The digestibility of mulberry fruit polysaccharides and its impact on lipolysis under simulated saliva, gastric and intestinal conditions. Food Hydrocoll. 2016, 58, 171–178. [Google Scholar] [CrossRef]
- Chen, J.; Liang, R.H.; Liu, W.; Liu, C.M.; Li, T.; Tu, Z.C.; Wan, J. Degradation of high-methoxyl pectin by dynamic high pressure microfluidization and its mechanism. Food Hydrocoll. 2012, 28, 121–129. [Google Scholar] [CrossRef]
- Harrington, R.E.; Zimm, B.H. Degradation of polymers by controlled hydrodynamic shear1. J. Phys. Chem. 1966, 69, 161–175. [Google Scholar] [CrossRef]
- He, Z.; Wang, X.; Li, G.; Zhao, Y.; Zhang, J.; Niu, C.; Zhang, L.; Zhang, X.; Ying, D.; Li, S. Antioxidant activity of prebiotic ginseng polysaccharides combined with potential probiotic lactobacillus plantarum c88. Int. J. Food Sci Technol. 2015, 50, 1673–1682. [Google Scholar] [CrossRef]
- Zhang, T.; Yang, Y.; Liang, Y.; Jiao, X.; Zhao, C. Beneficial effect of intestinal fermentation of natural polysaccharides. Nutrients 2018, 10, 1055. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Luo, D.H.; Liang, Z.Y. Structure of polysaccharides from the fruiting body of hericium erinaceus pers. Carbohydr. Polym. 2004, 57, 241–247. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Chemical analysis of microbial cells, In Methods in Microbiology; Academic Press: London, UK, 1956; pp. 272–274. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Biochem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- 3Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhao, L.; Cha, Q.; Sun, Y.; Ye, H.; Zeng, X. Structural characterization and immunostimulatory activity of a homogeneous polysaccharide from Sinonovacula constricta. J. Agric. Food Chem. 2015, 63, 7986–7994. [Google Scholar] [CrossRef] [PubMed]
- Braünlich, P.M.; Inngjerdingen, K.T.; Inngjerdingen, M.; Johnson, Q.; Paulsen, B.S.; Mabusela, W. Polysaccharides from the south african medicinal plant artemisia afra: Structure and activity studies. Fitoterapia 2017, 124, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Tadayoni, M.; Sheikhzeinoddin, M.; Soleimanianzad, S. Isolation of bioactive polysaccharide from acorn and evaluation of its functional properties. Int. J. Biol Macromol. 2015, 72, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, C.; Clement, V.; Rouvet, M.; Vallespamies, B. Dissolution tests as a tool for predicting bioaccessibility of nutrients during digestion. Food Hydrocoll. 2009, 23, 1228–1235. [Google Scholar] [CrossRef]
- Di, T.; Chen, G.; Sun, Y.; Ou, S.; Zeng, X.; Ye, H. Antioxidant and immunostimulating activities in vitro of sulfated polysaccharides isolated from Gracilaria rubra. J. Func. Foods 2017, 28, 64–75. [Google Scholar] [CrossRef]
- Yang, B.; Jiang, Y.; Zhao, M.; Chen, F.; Wang, R.; Chen, Y.; Zhang, D. Structural characterisation of polysaccharides purified from longan (dimocarpus longan lour.) fruit pericarp. Food Chem. 2009, 115, 609–614. [Google Scholar] [CrossRef]
- Schneider, S.M.; Girardpipau, F.; Anty, R.; Eg, V.D.L.; Philipsengeerling, B.J.; Knol, J.; Filippi, J.; Arab, K.; Hébuterne, X. Effects of total enteral nutrition supplemented with a multi-fibre mix on faecal short-chain fatty acids and microbiota. Clin. Nutr. 2006, 25, 82–90. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the simulated gastric and intestinal digesta of HEP in this work are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Molecular Weight (Da) | Reducing Sugar Content (mg/mL) |
|---|---|---|
| Stomach | ||
| 0.5 h | 936.8 ± 12.7a | 0.610 ± 0.007a |
| 1 h | 918.1 ± 0.9a | 0.644 ± 0.014a |
| 2 h | 905.3 ± 25.5a | 0.714 ± 0.015b |
| 4 h | 867.4 ± 37.6a | 0.791 ± 0.008c |
| 6 h | 867.0 ± 22.3a | 0.864 ± 0.008d |
| Intestine | ||
| 0.5 h | 575.9 ± 8.0b | 15.018 ± 0.188a |
| 1 h | 564.6 ± 10.9b | 16.463 ± 0.290ab |
| 2 h | 564.0 ± 8.7b | 17.337 ± 0.768c |
| 4 h | 530.1 ± 7.3b | 19.445 ± 0.602b |
| 6 h | 529.3 ± 7.2b | 22.698 ± 0.752d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhao, C.; Diao, M.; Zhong, S.; Sun, M.; Sun, B.; Ye, H.; Zhang, T. The Prebiotic Activity of Simulated Gastric and Intestinal Digesta of Polysaccharides from the Hericium erinaceus. Molecules 2018, 23, 3158. https://doi.org/10.3390/molecules23123158
Yang Y, Zhao C, Diao M, Zhong S, Sun M, Sun B, Ye H, Zhang T. The Prebiotic Activity of Simulated Gastric and Intestinal Digesta of Polysaccharides from the Hericium erinaceus. Molecules. 2018; 23(12):3158. https://doi.org/10.3390/molecules23123158
Chicago/Turabian StyleYang, Yang, Changhui Zhao, Mengxue Diao, Shuning Zhong, Maocheng Sun, Bo Sun, Haiqing Ye, and Tiehua Zhang. 2018. "The Prebiotic Activity of Simulated Gastric and Intestinal Digesta of Polysaccharides from the Hericium erinaceus" Molecules 23, no. 12: 3158. https://doi.org/10.3390/molecules23123158
APA StyleYang, Y., Zhao, C., Diao, M., Zhong, S., Sun, M., Sun, B., Ye, H., & Zhang, T. (2018). The Prebiotic Activity of Simulated Gastric and Intestinal Digesta of Polysaccharides from the Hericium erinaceus. Molecules, 23(12), 3158. https://doi.org/10.3390/molecules23123158